Copyright0 1975 American Society for Microbiology Printed in U.S.A.
Study of the Fine Structure
of
Adeno-Associated
Virus DNA
with Bacterial Restriction
Endonucleases
KENNETH I. BERNS,* JAMES KORT, KENNETHH. FIFE, E.WAYNEGROGAN, ANDILENE SPEAR
Department of Microbiology, The Johns Hopkins University SchoolofMedicine, Baltimore, Maryland21205
Received forpublication3April 1975
A physical map of the adeno-associated virus type 2 genome has been
constructed on the basis of the five fragments produced by the restriction
endonucleases HindII + IIIfrom Hemophilus influenzae. There are three endo
R HindII cleavage sites and one endo R HindIll site. Evidence has been
obtained to support the existence oftwonucleotide sequence permutations in
adeno-associated virusDNA, thestartpoints ofwhich have been estimatedtobe separated by 1% of thegenome.Thethree cleavage fragments produced by endo
R-EcoR, have been ordered and oriented with respect tothe endo R-HindII +
IIIcleavagemap.
Adeno-associated virus (AAV) is a small, defective, DNA virus which contains a linear
single-strandedgenome of 1.4 x 101molwt.For a productive infection adenovirus isrequiredas a helper (1, 12). AAV is unusual in that there are two types of virions which contain DNA molecules with complementary nucleotide se-quences (2, 4, 6, 12, 16). After isolation the
complementary polynucleotidechains
originat-ing from different virions base pair to form double-stranded DNA ifexposed to conditions
favorable for annealing. Because the genome contains a limited number of nucleotide se-quence permutations, two basictypes of linear duplexes may be formed, the first with even
ends, and a second with single-stranded cohe-sive termini. The latter may go on to form hydrogen-bonded circlesor concatenates (9).
In an effort to further elucidate the fine structure of the DNA isolated from AAV vi-rions, we have characterized with respect to
molecular weight and physical map order the fragments produced by digestion of double-stranded linear AAV DNA of unitlength using
three bacterial restriction endonucleases, endo R-HindII + III and endo R-Eco RI, isolated from Hemophilus influenzae and Escherichia
coli,respectively. Theseenzymescleave double-stranded DNAatspecific nucleotide sequences leading to the production ofunique fragments (13, 17). In this paper we report that AAV-2 containsthreeHindIIsitesandoneHinduII site. The resulting five fragments have been sized and ordered withrespecttotheir locationsalong
the genome. There are two R, cleavage sites in AAV-2 DNA (5), and the resulting
frag-ments have been oriented with regard to the fiveHind restrictionfragments.
The endo R-HindII + Ill digestionwas
com-patible with the presence of two major nucleo-tide sequence permutations in AAV DNA, the start points of which would be approxi-mately1%apart on the AAV genome.
MATERIALS AND METHODS
Production of AAV containing radioactively la-beledDNA. AAV-2 (AAV-2 H) (10) containing radi-oactively labeled DNA was grown in human KB cells co-infected withadenovirustype 2 ashelper in Eagle medium (8) which had been supplemented with 5% horse serum (Microbiological Associates). The multi-plicityof infection forbothAAVand adenoviruswas 10infectious units/cell. Toproduce 'H-labeled AAV DNA, ['H]thymidine (10 gCi/ml; New England Nuclear Corp.) was added 6 to 7 h after infection. AAV containing "P-labeled DNA were produced in KBcells which hadbeen suspended in lowPO4(10-' M) Eagle medium 10to 12h prior to infection. The "'P (9 pCi/ml; New England Nuclear) was added to the culture 7 h after infection. At 48 h after infection cells were harvested, and virus were puri-fied as previously described (4) by treatment with deoxycholate and trypsin and repeated equilibrium centrifugation in CsCl density gradients. The puri-ficationprocedure was modified by omission of steps involving sonication. After treatment with deoxy-cholate and trypsin the crude virus suspension was homogenized with 20 strokes using a Dounce homogenizer before additionofCsCl.
DNApurification. AAV DNA was isolated by ex-posure of virionsin 1 x SSC(0.15M'NaCl plus0.015 sodium citrate), 0.15% Sarkosyl to 0.1 N NaOH for 10 min atroomtemperatureand tosubsequent sedimen-tation through a 5 to 20% alkaline sucrose gradient (0.1 N NaOH, 0.9 M NaCl, 0.05 M EDTA, 0.15% 712
on November 10, 2019 by guest
http://jvi.asm.org/
Sarkosyl) usingthe SW50rotor in a Beckman model L ultracentrifuge (11). Conditions of sedimentation were 45,000 rpm for 4.5 h at 20 C. Intact AAV single strands were isolated from the gradient and dialyzed against 0.01 M Tris buffer, pH 7.8. To produce double-stranded AAV DNA the complementary single strands thus obtained were exposed to 0.1 N NaOH. After 10 min atroom temperature an amount of Tris buffer, pH 7.8, equivalent to the NaOH was added, and the solution was made 49% formamide (Mathe-son, Coleman, and Bell) and maintained at room temperature for 24 h. DNA concentration in the annealingprocedure was 2 to 10
Ag/ml.
Unit length duplex linear monomers were isolated from neutral sucrosegradients(9).Lambdabacteriophage DNA was agift from T. J. Kelly, Jr., andSV40 DNAwas agiftfrom D.Nathans. Digestion of DNA with restriction nucleases. Endo R-HindII + III and endo R-HindIII which had beenprepared as described (7) were kindly provided by W. Scott, N. Muzyczka, and D. Nathans. Endo R -EcoRIwasprepared byP.Geshelin, followingthe procedures of R. N. Yoshimori (Ph.D. thesis, Univer-sity of California Medical Center, San Francisco, 1971).
In all enzymedigestions a 0.10 volume of enzyme was used and the DNA concentration was 5 to 7.5
Mg/ml. The endo R HindII + III reaction mixture contained 7 mM 0-mercaptoethanol, 8 mM MgCl2, 6.6 mMTris, pH 7.6, and 50Mgofgelatin per ml. The digestionwasallowed toproceed for 1 to 3 h at 37C. A preparation of endoR-HindIl + IIIenriched for endo R-HindII was also used. The reaction mixture
con-tained 6.6 mMTris, pH 7.4, and 8.7 mMMgCl2 in addition to enzyme and DNA. Digestion conditions were as above. In the case of endo R * HindIII digestion the reaction mixture contained 7 mMMgCl,,6.6 mM Tris, pH 7.6, and 50 gg of gelatin per ml. The digestion wasfor 16 to 18 hat 37C. The endo R-Eco
R,digestionreaction contained 10 mM
MgCl2,
75mM NaCl,and 15mM Tris, pH 7.8. Digestion was for 2 h at 37C.Agarosegels.Cylindrical 10-cm 1.4%agarose gels were run using abuffer containing 40 mM Tris, pH 7.8, 5 mM Na acetate, and 0.1 mM EDTA in a Buchler Polyanalyst. Seakem agarose was obtained from MCI Biomedical. Electrophoresis was at 10 V/cm for 60 to 70 min atroom temperature. A 0.10 volume ofbromophenolblue in 50% sucrose and 1 mM EDTA was added to the sample as a marker. DNA
bands in thegelwerestainedby placing the gel after electrophoresis into gel-running buffer containing ethidium bromide (0.5Mg/ml)for 15 to 30 min. The bands were visualized under UV light and photo-graphed using Polaroid type 57 film.
Polyacrylamide gels. Vertical polyacrylamide slab gelelectrophoresis and subsequent autoradiogra-phy were carried out as described by Danna et al. (7).
Radioactivity assays. Assays of 3H and S2p radio-activity were done using Triton X-100/toluene (1:2) containing Omnifluor (4 g/liter; New England Nu-clear) and 10% water in a Beckman LS 230 scintilla-tion counter. In some cases samples containing 32P were assayed in the same counterbymeasuring Ceren-kov radiation.
81P labeling of 5' termini. Alkaline phosphatase and polynucleotide kinase (19) were provided by B. Weiss. [-y32PATP was obtained from New England Nuclear.Labelingofthe5'terminiofAAVDNA was doneaccording tothemethod of Weiss et al. (19).
RESULTS
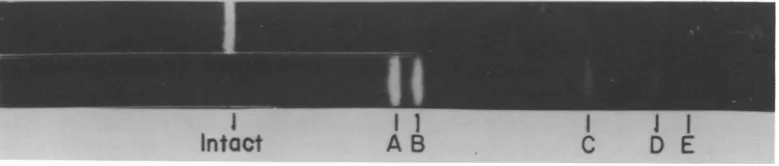
Digestion with endo R HindIl + III. A complete digestion oflinearduplex AAVDNA of unitlength withendoR-Hindll + III yielded fivediscrete bands (Fig. 1). Increasingthe time ofdigestion orthe amount of enzyme useddid not lead to any further fragmentation of the DNA. Decreased time ofdigestion resulted in the appearance of bands representing larger fragments (Fig. 2) which were consideredtobe the product ofpartial digestion. The five frag-ments resulting from complete digestion have been labeled A toE in order ofdecreasingsize (Table 1). The four fragments resulting from partialdigestionwerealllargerthanfragmentA and have been labeled 1 to4 (Table 1).
Determination of the molecularweightsof fragmentsA toE. The molecularweightsofthe fragments produced by endo R -Hindll + III digestion were estimated by comparing their mobilities to the mobilities in parallel gels of fragments ofSV40 DNA (form I) produced by the same enzymes (7) and of fragments of adenovirus type 2 DNA produced by digestion
with endo R -EcoRI (15). The molecularweights
I I I I I I
Intact
AB
C
D E
FIG. 1. A completeendoR*HindII + III digest of unit length linear duplex AAV DNA. Thefivefragments produced were separatedin a 1.4% agarose gel. Fragments have been labeled A to E in order of increasing electrophoretic mobility.
on November 10, 2019 by guest
http://jvi.asm.org/
[image:2.499.54.444.544.627.2]714
I
II I IIntact
1 23 4 A B
I
I
[image:3.499.64.461.69.149.2]C
D E
FIG. 2. Apartial digestion ofunit-length linear duplex AA V DNA was achieved by decreasing the time of digest. Fragments were separated in a 1.4% agarose gel. The four partial digestion products noted in the bottom gelhave been numbered 1 to 4 in order of increasing electrophoretic mobility. The two bands in the top gel corresponding to I and 4 represent the endo R HindIIIcleavage products.
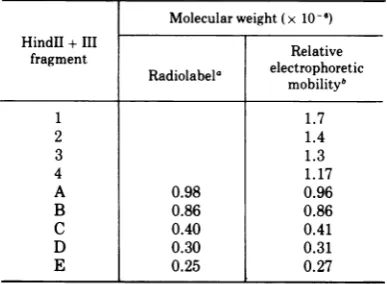
TABLE 1. Molecular weight estimates of AA V DNA fragments produced by cleavage with endo R -Hind
Molecularweight (x 10-6f)
HindII +III Relative
fragment Radiolabela electrophoretic
mobilityb
1 1.7
2 1.4
3 1.3
4 1.17
A 0.98 0.96
B 0.86 0.86
C 0.40 0.41
D 0.30 0.31
E 0.25 0.27
aUniformlylabeled
sP
DNA wasdigested, and theresultingbands cut out from a gel and counted. The fraction of total counts recovered in any one band were converted to molecular weight, assuming a molecular weight of 2.8 x 106forunit-length duplex DNA. The results are the averages of the data from twoindependentexperiments.
'Molecular weights were calculated from relative electrophoretic mobility as described in the text and are the averages of the data from two independent experiments.
for AAVfragmentsA to Erangedfrom 9.6 x 10' for A to 2.4x 105for E(Table 1). Bythis method
the molecular weight determined for intact
duplexlinear AAV molecules of unit
length
was 2.8 x 106 to 2.9 x 106, which is in goodagreement with the figure of 2.8 x 106reported
by Gerry et al. (9). The total molecularweight
determined forfragmentsAtoEwas2.79 x 10'. Because the additive molecular weight of the bandswasthesame asthatofintactDNA,this strongly suggested that each band observed in the gels represented a unique fragment, a
con-clusionsupportedbydatapresented below. Determination of the relative amounts of endo R HindII + III
fragments
A to E. In order todirectlyascertain therelativeamounts of the five endo R-HindII + IIIfragments
observed, a preparation of AAV DNAuniformly labeled with 32P was digested and electro-phoresedthrougha 1.4%agarosegel. The bands were stained with ethidium bromide, cut out, and counted (Table 1). The relative radio-activities of the five fragments were in good agreement with the molecular weights de-termined for each of the fragments by relative electrophoreticmobility.
Digestion with endoR EcoR1.Digestionof linear duplex AAV DNA of unit length with endo R-EcoRI producesthreefragmentsAtoC,
twoof whicharelargeandoneof whichisabout the same length or slightly smaller than endo R-HindII + HI fragment K of SV40 DNA (1.3 x 106) (Fig. 3). The faint band observed to
migrate to aposition between SV40 A and B is
tentatively attributed to contamination ofthe endo R-Eco RI preparation with a second en-zyme endo R Eco
RI.
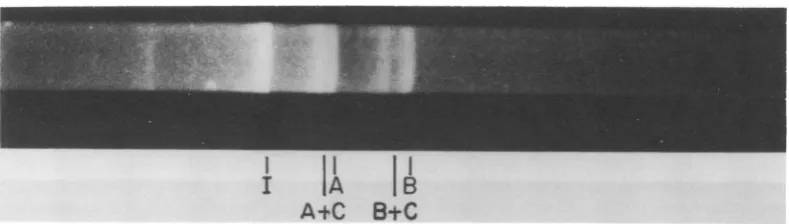
The relative electropho-retic mobilities indicated molecular weights of RI A and B of 1.57 x 106 and 1.1 x 106, respectively. Partialdigestionwith endo R-EcoRI
produced two fragmentsslightly larger than the two large fragments A and Bproduced by complete digestion, establishingamaporderofBCA(Fig.4).These dataare in agreementwith thoseofCarter and Khoury (5).
Alignment of the physical orders of the fragments produced by endo R*HindII + III and endo R-Eco
RI.
Inordertodeterminethetwo terminal endo R -HindII + III fragments,
unit-length duplex linear monomers were
la-beled at the 5' termini with 32P using
polynu-cleotidekinase. After phenol extraction the re-covered DNAwascleaved with endo R HindII + Im. Bands A to E were cut out, dissolved in concentrated perchloric acid, neutralized with NaOH, and counted. Only the bands corresponding tofragments B and C contained radioactivity (Table 2), thus identifying these astheterminal fragments.
With theknowledgethat endo R * HindII + III fragments B and C were terminal, the five J. VIROL.
on November 10, 2019 by guest
http://jvi.asm.org/
[image:3.499.62.256.226.368.2]fragments A to E wereassignedtotheproducts
of partial digestion with endo R -Hindll + III,
fragments 1 to 4, on the basis of molecular weight, i.e., combinationsofcompletedigestion
fragments A to E whose additive molecular weights most closely agreed with the molecular weights determined for partial digestion frag-ments 1 to 4 (Table 3). From this a tentative physical order of BEDAC was determined.
Endo R-HindllI cleaved unit length duplex
linear monomers at only one site, producing
endo R -HindII +IIIpartialdigestion fragments
1 and 4 (Fig. 2). These two fragments were separated on a neutral sucrose gradient and digested with endo R Hindll + III. The compo-sition offragment 1 was endo R-HindII + III fragments ACD, and the composition of
frag-ment 4wasendo R-HindII + IIIfragments EB
(Table 3).
Similarly, endo R-Eco
RI
fragments A and B were separated and digested with endo R Hindll + L. Endo R-Eco RI A containedendo R .HindII + III fragments A and C and a
fragmentslightly smaller than D. Endo R -Eco
RI
BcontainedendoR-HindII +IIIfragment
B and a fragment slightly smaller than E (Table 3).Cleavage of AAV DNA with an endoRH-Hind
preparation enriched for endo R HindIl pro-duced a new band between bands B and C (Fig. 5). The relative position of the new band corresponded to a molecular weight of 6 x 106, about what would be expected (5.8 x 105) for a fragment containing endo R-HindII + III
frag-ments D andE.
From these experiments we concluded that the physical order along the AAV DNA mole.. cule of the endo R Hindll + m fragments is BEDAC (Fig. 6). From a knowledge of the physical order and molecular weights of the products of digestion with endoRH EcoRI,it was possible to align these fragments on the endo R HHindII +Im cleavagemap(Fig. 6).
Evidence for two nucleotide sequence
per-Hin
d
31+M
SV40
I
I
I
..
T.-...
I
I
I
A
B
RTAAV
FIG. 3. A complete endo R-Eco R, digest of unit-length linear duplex AAVDNA. The three fragments produced were separated in a 4% polyacrylamide gel. Fragments have been labeled A to C in order of increasing electrophoretic mobility.As amolecularweight standard, aparallelgelcontaining anendo R HindII + III digestofSV40 DNAwaspresent.
I I IB
A+C B±C
FIG. 4. Apartial endo R-EcoR,digestion of unit-length linear duplex AAV DNA. Intact DNA, partial, and completedigestionproductswere separated on a 1.4% agarose gel. Fragment C is not visible.
on November 10, 2019 by guest
http://jvi.asm.org/
[image:4.499.46.445.312.474.2] [image:4.499.48.443.524.636.2]716 BERNS ET AL.
TABLE 2. Labeling of endo R HindII + III terminal fragments with32p by using polynucleotide kinasea
Fragment Counts/min
A 19
B 232
C 240
D 9
E 2
aFragments were visualized in a 1.4% agarose gel
[image:5.499.62.256.199.371.2]usingethidiumbromide, and were cut out, dissolved, and counted.
TABLE 3. Compositionof endo R HindII +IIIpartial digestion fragments 1 to 4and endo R EcoRI
fragments A and B
Composition Fragment Digestion
Mole-with endo cular R -Hindl weighta
+ III
endo R Hindll+III1 ACD ACD
endo R HindII + III2 AC
endo R HindHI+ III 3 AD
endo R HindII + III4 BE BEC
endo R EcoR, A ACD'b
endo R EcoRIB BE'b
aDetermination of composition by molecular
weight isdescribed in the text.
ID' and E' had slightly greatermobilities than D and E.
cThe molecular weight of partial digestion frag-ment 4 is actually closer to the additive molecular weights of fragments B + Dthan B + E. However, to be internally consistent with the other assignmentsin column 3, this fragmentwastentativelyidentifiedas B +E, which turned out to be also consistent with the data derived from furtherdigestionoffragments1and 4(column 2).
mutations in AAV DNA. Gerry et al. (9) concluded that AAV DNA contained a small
number of nucleotide sequence permutations,
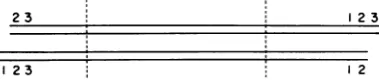
possibly only two, all of whose start points occurred within a limited region representing less than6% ofthegenome. This conclusion has been supported by thework of Koczot et al. (11) and Carteret al. (5). Digestion of a population ofDNAmoleculescontaining permuted nucleo-tide sequences with a bacterial restriction en-zyme should generate more than two terminal fragments (Fig. 7). Moreover, terminal frag-ments would not be equimolar with internal fragments. However, if the limited region of permutation in AAV DNA weresmallcompared to the average size of the terminal fragments
generated bythe restriction enzyme, resolution might prove to be difficult. With 1.4% agarose gels, asillustrated above, onlyfive bands were visible after digestion of unit-length duplex AAV DNA with endoR HindII + III. Further-more, all five bands were present inequimolar amounts including the two terminal fragments (B and C).
In order to improve resolution, the cleavage products of anendo R -HindII + III digestion of AAV DNA were separated by electrophoresis through a 4% polyacrylamide slab gel (Fig. 8). Under these conditions the bands representing fragments B and C were clearly resolved into twospecies. Additionally, onthe original auto-radiogram partialdigestion fragments 1, 2, and 4, which contain C orB, could also be resolved into doublets, whereas fragment 3 was still single. (In Fig. 8 only the 2and 4 doublets are stillvisible.) Assuming that the resolution of B andC into doubletsrepresentsspeciesdiffering in molecular weight rather than base composi-tion or conformation, the two species of frag-mentBdifferedinmolecular weight by approxi-mately 3 x 10' andthe twofragment C species differed by 2.8 x 10' to2.9 x 10' asestimated by mobility. If the resolution of terminal frag-mentsBandC into twospecies each did reflect the existence oftwonucleotidesequence permu-tations in AAV DNA, it would have been expected that the molecular weight difference between the two species of fragment B would have been equal tothe difference between the species offragment C as wasthe case.
Five experiments of the type illustrated in Fig.8 were performed. In every case thetwo C bands were sufficiently separated to be quan-titated by microdensitometry of the autoradio-gram.The AAV DNApreparation illustratedin Fig. 8 was used twice. The ratio of larger to smaller C bandspecies(asestimatedbyrelative
mobility)was0.67:1intheexperiment shown in Fig.8and0.88:1 in asecond experiment. Three more experiments with three different AAV DNA preparationsyielded ratios of1:1.Ineach case the presence of two B bands was evident but separation wasinsufficient toallow a simi-lar analysis. If we assume thatthe presence of twoB bands andtwo C bandson polyacrylam-ide electrophoresis does indeed represent the existence of twopermutations, the variation in the ratio may either reflect differences in the relative amounts ofeach permutationin differ-ent DNApreparationsorexperimentalartifact.
We conclude thatthe bulk of the evidence is
compatible with the existence of two major
nucleotide permutations in AAV DNA,
on November 10, 2019 by guest
http://jvi.asm.org/
I
I
I IA B
D+E
C
D E
FIG. 5. AA V linear duplex DNA was digested with an endo R HindII + III preparation enriched for endo R HindII. A new fragment corresponding to endo R HindII+ III D + E inmolecular weight was observed on electrophoresis through a 1.4% agarose gel.
Hind It+ m
Ba I E DI
n m n
RI
B
CI
FIG. 6. A cleavage map of theA. relative position of endo R EcoR,C from the molecular weight determ agarosegels for fragments A and B
23
23
FIG. 7. A modeltodemonstrateti linearduplex DNA moleculesconta tide sequencepermutations wouldyi terminal fragments with respect to from each end upon digestion w endonuclease.
thoughadditional structuralcor be absolutely ruledout.
DISCUSSION The
major
contribution of t regard to the fine structure of evidence compatible with the ecleotide sequence permutatioi strated by the high-resolution gels of endo R-HindII + III data might result from two
sj
duplex AAV DNA, one of wE deletion at each end. Thiswoi
difference inmolecular weight b species. We have been unable Ilengthduplex linear AAV DNA
onespeciesunder conditions with 0.6%agarose C gels where two lambda bacteriophage DNA endo R Eco
RI
fragments of approximately AAV size which differby aslittleas 2% inmo-lecular weight may be resolved (N. Muzyczka, personal communication). A deletion of 1% at only one orthe other endofthe DNA molecule would be the equivalent of the suggested
A permutation model. No evidence for subterm-inal or more internal deletions of 1% or more AVgenome. The has been obtained from high-resolution elec-was determined tron microscopy (Berns and
Kelly,
unpub-inations on 1.4% lished data). However, a 1% deletion (30to 40 (seeFig. 3). nucleotides) would be close to the borderline ofdetectability in our experience. Single-strand
2 3 terminion some molecules
might
contribute to heterogeneity in terminal fragments, but there is no evidence for such termini in unit length2 duplex linear AAV DNA molecules isolated hat a collectionof from neutralsucrosegradients(9).
ining two nucleo- It is not completelyclear why we areunable ield two species of to resolve the two endo R HindII + III AAV C molecularweight species in 1.4%agarose gels. However, we have ith a restriction beenable to separate the two species of C using 0.6% and 0.4% agarose gels although overall nplexitycannot resolution suffers and diffusion of the bands
increases (datanot shown).
Again assuming that the resolution of the terminal endo R HindII + III fragments into twospecies eachonpolyacrylamide gelsreflects ;his paper with primarily difference in molecular weight, these f AAV DNA is gels haveenabled usto measuretheseparation Lxistence of nu- onthe AAVgenomeofthe starting pointsforthe ns as demon- twopermutations. The figure of 1% of the AAV polyacrylamide genome is especially intriguing because this is
ligests.
Similar the same valuedeterminedforthe lengthofthepecies of linear natural terminal nucleotidesequencerepetition iich had a 1% by Gerry et al. (9) and is close to the value of ald imply a 2% 1.5% reported for the length of the inverted
letween
the two terminal repetition by BernsandKelly (3).This to resolve unit- is the figure which might be expected if AAV into more than DNA replicated via a concatenate and wereon November 10, 2019 by guest
http://jvi.asm.org/
[image:6.499.47.444.80.197.2] [image:6.499.45.237.241.338.2] [image:6.499.46.236.393.435.2]718
,SI
I 11I IY
1
234
AB
I
C
I
D
E
FIG. 8. An endo R HindII+IIIdigest of unitlengthlinearduplexAAVDNA molecules waselectrophoresed through a4%polyacrylamideslabgel.A complete digest displayeddoublet bandscorresponding to terminal endo R HindII + IIIfragmentsB and C. Apartial digestionrevealed thatpartial digestion products (2and4) containingterminalfragmentsBand Cwerealso doublets.
packaged via a "headful" mechanism as has been suggested for some bacteriophage DNAs (18). However, theexistence of only two permu-tationswould imply the longest concatenate to be onlya dimer or trimer.
We havebeen interested in the fine structure of AAV DNA because of the implications of such studies with regard to events at the molec-ular level during viral multiplication. The abil-ity to obtain unique fragments representing specific segments ofthe mature genome should be useful with regard to transcriptional map-ping andthe sequencing of interesting parts of the AAVgenome. Segments of particular inter-estwould includethe termini which have been reported tohave properties consistent with both a natural terminal nucleotide sequence repeti-tion (12----12)andaninverted terminal nucleo-tide sequence repetition(12----2'1',where 2' and 1' represent nucleotide sequences complemen-tary to 2 and 1, respectively) (3, 9, 11), and potential initiation and termination sites of transcription.
ACKNOWLEDGMENTS
We wish to thank J. LoprestiOlsen for technical assistance and H.0. Smithand D.Nathansforhelpfulsuggestions with regard to the manuscript.
This work wassupported bygrants from theU.S. Public HealthService(1RO1Al10843 fromthe National Institute of Allergy and InfectiousDiseases) and the National Science Foundation (GB 37950). K.I.B. is a Howard HughesMedical Investigator. J. K. and K. H. F. were supported by the HughesMedicalInstitute, andI.S. wassuppoftedbyPublic Health Service grant 5TO1 GM 00795 from the National InstituteofGeneral MedicalSciences.
LITERATURE CITED
1. Atchison, R. W., B. C. Casto, and W. McD. Hammon. 1965. Adenovirus-associated defective virusparticles. Science 194:754-756.
2. Berns, K. I., and S. Adler. 1972.Separationof twotypes ofadeno-associatedvirusparticlescontaining comple-mentarypolynucleotidechains. J. Virol. 9:394-396.
3. Berns, K. I., and T. J. Kelly, Jr. 1974. Visualization of the inverted terminal repetition in adeno-associated virus DNA. J.Mol. Biol. 82:267-271.
4. Berns, K. I., and J. A. Rose. 1970. Evidence for a single-stranded adenovirus-associated virus genome: isolation and separation of complementary single strands. J. Virol.5:693-699.
5. Carter, B. J., andG. Khoury. 1975.Specific cleavage of adenovirus-associated virus DNA by restriction endo-nuclease R EcoR,-characterization ofcleavage prod-ucts.Virology63:523-539.
6. Crawford,L.V., E. A.C. Follet,M.G.Burdon, and D. J. McGeoch. 1969. The DNA of a minute virus of mice. J. Gen.Virol. 4:37-46.
7. Danna, K. J., G. H. Sack, Jr., and D. Nathans. 1973. Studiesofsimian virus40DNA. VII. Acleavage map of the SV40 genome. J. Mol. Biol. 78:363-376.
8. Eagle, H. 1959. Aminoacidmetabolism inmammalian cellcultures.Science 130:432-437.
9. Gerry, H. W.,T. J. Kelly, Jr., and K. I. Berns. 1973. Arrangement of nucleotide sequences in adeno-associated virus DNA. J. Mol. Biol. 79:207-225. 10. Hoggan, M. D., N. R.Blacklow, and W. P. Rowe. 1966.
Studiesofsmall DNA virusesfound in various adenovi-ruspreparations: physical,biologicaland immunologi-cal characteristics. Proc. Natl. Acad. Sci. U.S.A. 55:1467-1471.
11. Koczot, F.J.,B. J.Carter, C.F.Garon,andJ. A. Rose. 1973. Self-complementarity of terminal sequences withinplusorminusstrands of adenovirus-associated virusDNA. Proc.Natl. Acad.Sci. U.S.A.70:215-219. 12. Mayor, M.D., K. Torikai, J. L.Melnick,and M. Mandel. 1969.Plusand minussingle-stranded DNAseparately encapsidated in adeno-associated satellite virions. Science166:1280-1282.
13. Morrow, J., and P.Berg. 1972.Cleavageof simian virus 40 DNA at a unique site by a bacterial restriction enzyme. Proc. Natl.Acad.Sci. U.S.A.69:3365-3369. 14. Old, R., K. Murray, andG. Roizes. 1975. Recognition
sequenceofrestrictionendonucleaseIllfrom Hemophi-lusinfluenzae.J. Mol. Biol. 92:331-340.
15. Pettersson,U., C. Mulder, H. Delius, and P.Sharp.1973. Cleavage ofadenovirus type 2 DNA into six unique fragments by endonuclease R-R,. Proc. Natl. Acad. Sci. U.S.A.70:200-204.
16. Rose, J. A., K.I.Berns, M. D. Hoggan, and F.J. Koczot. 1969. Evidence for a single-stranded adenovirus-associated virus genome: formation ofaDNAdensity hybridonrelease of viral DNA. Proc.Natl. Acad.Sci. U.S.A. 64:863-869.
17. Smith, H. O., and K. W. Wilcox. 1970. A restriction enzyme from Hemophilus influenzae. I. Purification andgeneral properties.J. Mol. Biol. 51:379-392.
.4
9(
on November 10, 2019 by guest
http://jvi.asm.org/
[image:7.499.66.458.75.155.2]18. Thomas, C. A., Jr., and L. A. MacHattie. 1967. The
anatomyofviral DNAmolecules, p.485-518. In P. D.
Boyer (ed.), Annual review ofbiochemistry, vol. 36. AnnualReviews, Inc., Palo Alto, Calif.
19. Weiss, B., T. R. Live, and C. C. Richardson. 1968.
Enzymatic breakage andjoining ofdeoxyribonucleic
acid. V. Endgrouplabelingandanalysisof
deoxyribo-nucleic acid containingsinglestrandbreaks. J. Biol. Chem.243:4530-4542.
on November 10, 2019 by guest
http://jvi.asm.org/