Copyright © 1994, American SocietyforMicrobiology
Human
Immunodeficiency
Virus
Type
1 RNA
Expression
by
Four
Chronically Infected Cell
Lines Indicates
Multiple
Mechanisms
of Latency
SALVATORET.BUTERA,1* BEVERLY D.
ROBERTS,'
LEELAM,2THOMAS HODGE,2 ANDTHOMAS M. FOLKS'RetrovirusDiseasesBranch, DivisionofViral and RickettsialDiseases,1andImmunologyBranch, Divisionof HIV/AIDS, 2 National CenterforInfectious Diseases, CentersforDisease
Control andPrevention,Atlanta, Georgia 30333 Received25February1993/Accepted 17December1993
Recentinformationhassuggestedthatposttranscriptionalmechanisms,wherebyhumanimmunodeficiency
virus type 1 (HIV-1) RNA exists as multiply spliced transcripts withoutpromoting an accumulation of the larger messages, are responsible for maintaining a stable state ofnonproductive viral expression or viral latency.To testtheuniversality oftheseobservations,wecomparedthepatterns of viralRNAsplicingand the frequencies of cells actually harboring HIV-1 RNA in four
chronically
HIV-1-infected cell lines (Ul [promonocytic],ACH-2 [Tlymphocytic], OM-10.1 [promyelocytic],andJ1.1 [Tlymphocytic]).In uninduced UlandACH-2 cultures,ahighfrequencyof cells(approximatelyoneinsix)contained HIV-1 RNA butmainly asmultiplysplicedtranscripts, again supportingaposttranscriptional mechanism maintainingvirallatency. Insharpcontrast, only 1 in50 cellsinuninducedOM-10.1andJ1.1culturescontainedHIV-1RNA,indicating aprimarytranscriptional mechanismcontrolling viral expressionin these cells.Furthermore,those OM-10.1 andJ1.1cells thatdid contain viralRNA werein astateofproductiveHIV-1expressionmarkedbythepresence of both spliced and unspliced transcripts. Even though the total absence ofviral RNA in the majority of OM-10.1 and J1.1 cells indicated a state ofabsolute latency, treatment with tumor necrosis factor alpha induced transcription of HIV-1RNA innearly 100% ofthe cells in allfour ofthechronicallyinfected cultures. Tumor necrosis factoralphainductionofUl,ACH-2,and OM-10.1 cultures resulted inaninitialaccumulation ofmultiply spliced HIV-1 RNA followed byatransitiontothelargerunsplicedviralRNAtranscripts.This RNA splice transition was less apparent in the J1.1 cell line. These results demonstrate that host cell-specific transcriptional andposttranscriptional mechanismsareimportantfactors in the controlofHIV-1latency. During the extended clinicallyasymptomaticperiodfollow-ing humanimmunodeficiency virus type 1 (HIV-1) infection, the opportunities for medical intervention are potentially immense (7). However, such intervention will require a de-tailed understanding of the various mechanisms that control HIV-1latency and expressionat acellular level(1). Advances in this areahave been made by studyingchronically infected cell models in which constitutiveHIV-1 expression is minimal butcanbeinduced physiologicallywithcytokines (2).
Afterintegration,HIV-1 geneexpressioncanbedivided into early and late stages. Initially, the predominance of viral mRNAismultiply splicedto2.0-kbtranscripts that encode the regulatory proteins (including Tat and Rev) necessary to propagateviral expression (8, 11,15).Atransition thenoccurs to permit the accumulation of singly spliced (4.3-kb) and full-length (9.3-kb) RNAspecies encoding the viral structural proteins andproviding the genomic RNA for budding virions
(8, 11). The mechanisms that control this early-to-late-stage transition in viral RNA splicing is reported to involve the action of the Rev protein in the protection and transport of full-length HIV-1 RNA(9, 13).
In chronically infected Ul and ACH-2 cultures, the consti-tutive pattern of HIV-1 RNA splicing previously showed a predominanceofthe2.0-and 4.3-kb viral messages coincident
*Corresponding author. Mailing address: Retrovirus Diseases
Branch,Centers for Disease Control and Prevention, 1600 Clifton Rd., NE,MS-G19, Atlanta, GA 30333. Phone: (404) 639-1024. Fax: (404) 639-1174.
with a relative absence of full-length message (14, 21). In addition,aninitial accumulation ofmultiply spliced viral RNA after induction of these cultures that mimicked the early-to-late-stage transition of HIV-1replicationwasobserved. There-fore,thenonproductivepatternofconstitutive viral expression in these cultures was appropriately described as blocked early-stage latency (22), whereby HIV-1 RNA is present but selectivelysplicedtorestrictproductiveviralexpression.These findings suggested that aprimary posttranscriptional mecha-nism was responsible for maintaining nonproductive HIV-1 expression and that a threshold level of Rev protein was necessary to drive productive viral expression (20, 21). Addi-tionalsupportforthe Rev threshold hypothesis has arisen from observations ofanidenticalHIV-1 RNAsplicepatternduring infectionwithrevdeletion mutants (8) and that multiple Rev protein monomers mustbindtotheRev response elementof the viral RNAto achievefull functional activity (12).
To testthe universalityofposttranscriptional regulation of HIV-1 expression,we comparedthepatterns ofHIV-1 RNA
splicingandthefrequencyof cellsharboring viralRNAinfour independently derived, chronically infected cell lines. The promonocytic Ul (10), promyelocytic OM-10.1 (3), T-lympho-cytic ACH-2(5),andT-lymphocyticJ1.1(19) lineswerecloned
followinganacuteHIV-1 (LAI)infection of their parental cell lines(U937, HL-60,A3.01, andJurkat, respectively). All four infected lines harbor one or more integrated provirionsand
constitutively express very low amounts of HIV-1 proteins when maintainedin RPMI 1640mediumcontaining 10%fetal bovineserum. However, HIV-1 expression canbe induced in 2726
on November 9, 2019 by guest
http://jvi.asm.org/
NOTES 2727 each cell culture using recombinant human tumor necrosis
factor
alpha (TNF-ot).
Todetermine thefrequencyofcellsharboring viral RNA in uninduced andTNF-aK-inducedculturesatthesingle-cell level, reverse transcriptase-dependent PCR (RT-PCR) was per-formed on
limiting
cell dilutions. Chronically infected cells werecultured in the presence orabsence of TNF-a for 24 h, andthen serial twofolddilutionsof the cells into their respec-tiveuninfectedparentalcell lineswere prepared so that each dilution contained a total of 106 cells. Cell dilutions wereperformed prior
to RNA extraction to ensure that the fre-quencyresultswouldnotbeartifactually influencedby individ-ual cellsharboring
a relative abundance of viral RNA. Total RNAwasextractedfromeachinfected cell dilutionby using acommercially
available kit(Clontech
Laboratories, Inc., PaloAlto, Calif.), digested
with DNase (Promega Corp., Madison,Wis.)
for 30 min at37°C,
chloroform-isoamyl alcoholex-tracted,
andethanolprecipitated.
The RNApelletwasresus-pended
in 4 ,ul ofdiethyl
pyrocarbonate-treated water andsubjected
to reversetranscription
at 42°C, using random hexamerprimers,
as directedby
the kit's manufacturer (Per-kin-ElmerCetus, Norwalk, Conn.).
Half of the resultant cDNAproduct
wasamplified
ina100-,ul reaction volumeby usingthegag-based primers
SK38and SK39(16)
asdescribedpreviously(23),
and the other halfwasamplified
by using the 5' long terminal repeat(LTR)-based primers
FPL1 and FPL2 asdescribed
previously
(14).
Tenmicroliters of each of the finalamplification
products
wasanalyzed
by agarose gelelectro-phoresis
andSouthernhybridization
to32P-end
labeledprobe SK19(16)
forgag-based primer
reactions orLTRP3(14)
for LTR-basedprimer
reactions. The amplified products of gag-basedprimer
reactionswere furtherquantitated by luminom-etry afterliquid-phase
hybridization
with HIV-1gag-specific,
acridinium
ester-conjugated probes
asspecified
bythemanu-facturer
(Gen-Probe,
Inc., SanDiego, Calif.).
Detection of all viral RNA
transcripts,
regardless of thesplicing
event,was achievedby
using
PCRprimers
andprobespecific
for sequences within the 5' LTR(14, 15).
Asshown inFig. 1,
thefrequency
of uninduced Ul and ACH-2 cellsexpressing
HIV-1 RNAwasrelatively high,
approximately20%(>1
in6).
However,
by comparison,
thefrequency
of cells in uninduced OM-10.1 and J1.1 culturestranscribing
LTR-con-taining
messageswasdramatically lower, approximately
1 in 50.Long
exposuresof thehybridization
reactionwereusedtodeterminethe final end
point
ofreactivity depicted
inFig.
1. Because of the technicalvariability
inherentto dilutionson asingle-cell
level andRT-PCR,
three additional cell titrations wereindependently prepared
andamplified.
Inevery case,thefrequency
of cellspositive
forLTR-containing
messages in uninduced OM-10.1 cultures was at most 1 in50,
while thefrequency
remained muchhigher
in uninduced Ul cultures(data
notshown). Thus,
thevastmajority
ofuninduced OM-10.1 and J1.1 cells containnodetectable viralRNA,
indicating
a state of absolute
latency
on a cellularlevel,
while a muchhigher frequency
of uninduced Ul and ACH-2 cells harbor HIV-1LTR-containing
RNA. Dilutional RT-PCR was alsoperformed
24 hafter TNF-otinduction,
andin allfour of the celllines,
TNF-a treatment resulted in an accumulation ofLTR-containing
viralRNAinnearly
100% of the cells(Fig. 1).
Asimilar
approach
was usedtodetermine thefrequency
of cellsexpressing full-length
HIV-1 RNA except thatprimers
and
probes
targeting
the gagregion
wereused.Becausethe gagregion
of the viral RNA is removedduring
splicing (8),
gag-based primers
willamplify only full-length
HIV-1 RNA.Amplification products
from the HIV-1 gagregion
of theexpected
molecular sizewereconfirmedby
Southernhybrid-Medium +TNF-a
O O Oen Ie 0 a° n n o
-I
l;i.
0. 0 l
Ol
I.
FIG. 1. Limiting-dilutionRT-PCR of four chronically infected cell lines eitheruninduced (Medium) or after HIV-1 induction (+TNF-ot).
HIV-1 inductionwasperformed by using human recombinantTNF-cK
(Genzyme Corp., Cambridge, Mass.)at 20 U/ml forOM-10.1 cultures and at 100 U/ml forUl, J1.1, and ACH-2 cultures for 24 h. The number of infectedcells per culture is indicated above each lane. Total cellularRNAwaspurified fromeach culture, DNase treated, reverse transcribed, and PCR amplified by using LTR-based primers and probe for detection of all HIV-1 RNA. Amplified products were detectedbySouthern hybridizationand analyzed by PhosphorImager (MolecularDynamics, Sunnyvale, Calif.) analysis. A series of control reactions was performed to verify that the PCR-amplified products were reverse transcriptase dependent (not shown). In each case, amplified products ofthe expectedmolecular sizes were generated; spurious bands were presumably due to partial RNA degradation. Extendedautoradiographicexposuresof each blot were performed to determinethecellularfrequency of detection (markedoneach panel by an arrowhead). Results are representative of five independent experiments.
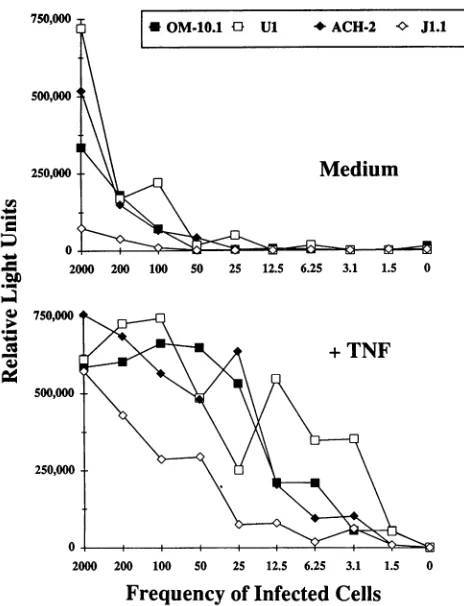
ization andwerequantitated by luminometry using acridinium ester-conjugated,gag-specific probes (Fig. 2). All four chroni-callyinfected cultures when maintained in medium showed a frequency of approximately 1 in 50 cells containing gag-encoding viral RNA. However, after TNF-a treatment, the frequency of cells expressing full-length viral RNA increased tonearly100% in all four cultures(Fig. 2), indicating complete conversiontoproductive HIV-1expression.
Thefrequencyof RT-PCR-positive cells withuseof LTR-based primers was compared with that obtained with use of gag-based primers (Table 1). In uninduced Ul and ACH-2 cultures,LTR-based RT-PCR detectedacellularfrequencyof approximately20% (>1 in 6), much higherthan the cellular frequencydetectedby gag-basedamplification (approximately
1 in 50). Therefore, the viral RNA pattern observed with uninduced Ul and ACH-2 cells by RT-PCR mimicked the blocked early-stage latency pattern reported by standard Northern(RNA)blotanalysis(14, 21)in thatahigh frequency of cellsexpressedsplicedHIV-1 RNA in the relative absence of full-length transcripts. In uninduced OM-10.1 and J1.1 cultures, the cellularfrequency observed byboth LTR-based andgag-based RT-PCRwas identical,
approximately
1 in 50. Thus, the small proportion of uninduced OM-10.1 and J1.1 cells that didcontain viral RNAwereina state ofproductive expression, marked by the presence of bothspliced
and full-lengthtranscripts.
However, the vastmajority
of unin-duced OM-10.1 and J1.1 cells contained no detectable viral VOL.68, 1994on November 9, 2019 by guest
http://jvi.asm.org/
[image:2.612.335.547.77.252.2]2728 NOTES
* OM-10.1 £1 Ul *ACH-2 v J1.1
Medium
Ul
9- _ 0
OM-10.1
9 v Cl
0-c 9.3 kb
< 4.3 kb >
c 2.0kb >
< 9.3 kb >
4.3kb
C2.0 kb >
9,
Jc m#14 co cm coV0Ov co C4 v
_ _ C - - N
ACH-2 J1.1
250,000
0 2
2000 200 100 50 25 12.5 6.25 3.1 1.5 0
Frequency
of Infected Cells
FIG. 2. Limiting-dilution RT-PCR of four chronicallyinfected cell
lines eitheruninduced (Medium)orafterHIV-1induction(+ TNF)as described forFig.1exceptthatRT-PCRwasperformedwithgag-based primers. Amplified products of the expected molecular sizes were
verified by Southern hybridization and quantitated byluminometry using acridinium ester-conjugated probes (Gen-Probe). Results are
reported asrelative light units for each frequency of infected cells.
Resultsarerepresentative oftwoindependent experiments.
RNA,andblockedearly-stage latencywasnotevident in these cultures. After TNF-o induction, HIV-1 RNA could be de-tectedtothesingle-celllevel inall four celllineswhenanalyzed with eitherset of RT-PCRprimers (Table 1).
FIG. 3. Kinetic induction of HIV-1 expression from four
chroni-callyinfected cell lines afterTNF-cx treatment.HIV-1-specific RNA wasdetectedfrom 25 pgof totalcellularRNAperlane for Ul and 10
,ugof total cellular RNAperlanefor the otherthree cell lines. The
quantity and integrity of the separated RNA was confirmed by ethidium bromide staining and UV visualization of the ribosomal
bands(not shown).Aradiolabeled HIV-1 5' LTRprobewasused in
hybridization, andtheimagewasdeveloped by autoradiography (7-day exposure). For each cell line, a representative blot depicting the patternobservedin severalindependent experimentsis shown. HIV-1
expression wasverified in cell-free culture supernatants by p24
en-zyme-linked immunosorbent assay (Coulter Immunologics, Hialeah, Fla.)orbyreverse transcriptaseassay(29),withinductionafter 72 h
being>100-fold in OM-10.1cultures,>50-fold inU1cultures,and
12-to15-fold in J1.1 and ACH-2 cultures.
We then examined the kinetic pattern of HIV-1 RNA splicing during TNF-ct induction at the population level by standard Northern blot analysis (Fig. 3). Total RNA was
purifiedfrom guanidine thiocyanate cell lysates (26),
quanti-TABLE 1. Numbersof cellsexpressingHIV-1 RNAinuninduced andTNF-oa-inducedchronicallyinfectedculturesa
Reaction
No.of OM-10.1 ACH-2 Ul J1.1
infected cells Medium +TNF-a Medium
+TNF-ot
Medium +TNF-a Medium +TNF-aLTR gag LTR gag LTR gag LTR gag LTR gag LTR gag LTR gag LTR gag
2,000 + + + + + + + + + + + + + + + +
200 + + + + + + + + + + + + + + + +
100 + + + + + + + + + + + + + + + +
50 + + + + + + + + + + + + + + + +
25 - - + + + - + + + - + + - - + +
12.5 - - + + + - + + + - + + - - + +
6.25 - - + + + - + + + - + + - - + +
3.125 - - + + - - + + - - + + - - + +
1.56 - - + + - - + + - - + + -
-0 - - - _ _ _ - - _
a Culturescontaining known numbers of infectedcells were prepared in106uninfectedparentalcellsand subjected toRT-PCR,using either LTR-based primers(for
thedetectionofall viralRNAtranscripts)or gag-basedprimers(for thedetectionof full-lengthviralRNA). Results are reported as a positive or negative RT-PCR
reactionafterspecificSouthernhybridizationfor the LTR reactions orluminometryfor the gagreactions.
750,000
500,000
250,000
Cl
o
2000 200 100 50 25 12.5 6.25 3.1 1.5
nA
'a
;0-.3
*>.
co
PS
750,000
500,000
0
J. VIROL.
on November 9, 2019 by guest
http://jvi.asm.org/
[image:3.612.58.290.75.378.2] [image:3.612.54.557.547.698.2]NOTES 2729
tated, and electrophoresed through formaldehyde-containing agarosegels (3). After Northern transfer, HIV-1-specificRNA wasdetected by hybridization using a32P-labeled2.5-kb XbaI-PstI fragment of pHXB2 (25), containing the 5' LTR, as described previously (4). The viral RNA patternobserved at the population level foreach culture beforeTNF-ainduction (T-0) accurately reflected thepattern observed at the cellular level by RT-PCR. Accordingly, uninduced Ul cultures dis-played a viral RNA pattern dominated by the spliced tran-scripts with an absence of full-length message (Fig. 3). The same was true for uninduced ACH-2 cultures when the T-0 pattern was enhanced by longer autoradiographic exposures (data not shown). This pattern is evidently due to the high frequency of cells displaying block early-stage latencyin these cultures. In contrast, the viral RNA pattern from uninduced OM-10.1 and J1.1 cultures had evidence offull-lengthHIV-1 RNA. This pattern is apparently contributed by the small percentageof cells that do containviral RNA andareinastate ofproductive expression as evidencedby RT-PCR.
Viral induction in each cell linewasevident byan accumu-lation of HIV-1-specific mRNA after 4 to 8 h of TNF-a treatment (Fig. 3). InUl, ACH-2, and OM-10.1 cultures, the initial viral RNA thataroseafterinductionwaspredominantly spliced to the 2.0-kb messages encoding the regulatory pro-teins. In these cultures, a transition to the singly spliced and full-length messages occurred within 12 h of
induction,
after which all three viral RNA bands continued to accumulate. Laser densitometryverified the initial accumulation of 2.0-kb viral RNAand thetransitiontothelargermessagesduringthe time course of TNF-ao induction in these cultures(data
notshown). The initial accumulationof the
multiply
spliced
mes-sages and the early-to-late-stage transition in these cultures supportthe possibility thata threshold levelof Revactivity
is requiredtodriveHIV-1expressionafter activation(12,
14, 20, 21). The kinetic viral RNA pattern observed in J1.1 cultures after inductionwasdifferent in thatnoinitialpredominance
of the multiple spliced RNAspecieswas observed(Fig. 3).
This may indicate a lower threshold requirement for Rev in these cellsor ahigher endogenouslevel of host factors necessary for Rev activity (27, 28).The mechanisms controlling viral quiescence among these cell systems appear to exist on at least twodistinct levels. As observedin a
proportion
ofUl and ACH-2cells,
onelevel of control existsposttranscriptionally,
since HIV-1 RNA is tran-scribed butprocessedviasplicing
tomaintainanonproductive
viralstate.This blockedearly-stagelatency
patternwasevident at the cellular levelby
RT-PCR and at the culture levelby
standard Northern
analysis.
Theposttranscriptional
control mechanisms of Ul and ACH-2 cells may involve additional host cell factors essential for Revactivity
(27, 28)
that may need toaccumulateafter cellularstimulationtodriveproduc-tive HIV-1
expression. Alternatively,
viralquiescence
in OM-10.1 and J1.1 cellsisdue tostricttranscriptional
control.This wasclearlyevidentonacellular levelby
RT-PCR because the vastmajorityofuninduced cellscontained no detectable viral RNA. On apopulation
level,
the small percentage(approxi-mately
2%)
of cellsconstitutively
expressing
HIV-1 gave evidence offull-length
viral RNAby
Northern
analysis.
Cellu-lar factors may also be critical formaintaining
the state of latency in OM-10.1 and J1.1 cells. Thisregulation
could potentiallyinvolve factorsworking
atthe level of theintegrated
provirus that prevent
transcription
or an absence ofpositive
regulatoryfactors necessaryto driveviral
transcription.
Furthermore,
multiple
mechanisms may becontrolling
HIV-1
expression
within Ul and ACH-2cultures,
sinceonly
approximately 20% of the uninduced cells exhibited the
blocked
early-stage
pattern oflatency.
By RT-PCR analysis, themajority
ofuninduced Ul and ACH-2 cellsappeared tocontain noviral RNA and maybe
subject
to control mecha-nisms similartothose exhibitedby OM-10.1 andJ1.1cultures. Theability
ofUl andACH-2cellsto transcribeHIV-1 RNA mayoccurduring
adiscretestageof the cellcycleorbe under the control of other mechanismsinfluencing
geneactivity,
therefore
permitting only
aproportion
of theuninduced cells toharbor viral RNAatanyonetime. Thesamemaybetruefor OM-10.1 and J1.1 cultures becauseapproximately
2% ofthe cellsproductively
express HIV-1 RNAandprotein
atanygiven
time.
The relative absenceof
full-length
HIV-1 RNA message in uninduced Ul andACH-2cells suggests thatalowconstitutive level ofRevprotein
may be aposttranscriptional
mechanismresponsible
formaintaining nonproductive
viralexpression
(12, 14, 21).
Our results with OM-10.1 and J1.1 cells suggest thatmechanismsatthe level oftranscription
mayalsocontrol constitutive HIV-1expression.
Because cell linesdisplaying
different mechanisms of controlover
latency
each underwentaselective
splice
switch to theregulatory
messages upon viralinduction,
theaccumulation ofRevtoacritical threshold level remains apotential unifying
mechanism for the activation of HIV-1expression.
Thesamemaybe truefor the J1.1cellline,
although
the critical Revthreshold appearstobe lower duetocell
line-specific
differences.Thecontribution ofHIV-1
latency
on acellularleveltothe clinicalprogression
toAIDS remainspoorly
defined.Although
active HIV-1
replication
canbe detectedatall stagesof AIDSprogression,
including
theclinically
asymptomatic phase
(17,
18),
alarge
numberofinfected cells within thelymph
nodes harbor HIV-1 DNA in the absence of detectable viral RNA(6).
Thisdiscrepancy
between viral DNA and RNA suggestsastateof absolute viral
latency
and is reminiscent ofthe pattern observed inthe OM-10.1 and J1.1 cell lines.Fromourin vitro observations with these celllines,
itmight
beexpected
that when these infectedlymph
node cells in vivo encounter the proper combination of extracellularstimuli,
a conversion toactive viral
transcription
can occur.Furthermore,
the blockedearly-stage latency
pattern observed with the Ul and ACH-2 celllines has also been detected in theperipheral
blood cells of HIV-1-infected individuals(24).
Thesefindings
suggest that HIV-1latency
on acellular level isanimportant
aspect of the clinical disease and that HIV-1 activation fromlatency
may contribute tovirus disseminationand diseaseprogression.
We report on the HIV-1 RNA patterns in four
indepen-dently
derived, chronically
infected cell linesduring
uninduced and induced viralexpression.
These cell systems represent at least two different and distinct mechanismsgoverning
HIV-1expression
with variousdegrees
of cellular and viral control and willserve asimportant
systemsofcomparison.
Itwill beof continued interest and medicalimportance
toinvestigate
the molecular mechanisms involved incontrolling
the state ofrepressed
viralexpression
and thepathways
responsible
for HIV-1 activation.WethankP.Guentherfortechnical involvement with the RT-PCR assays and M. Saltarelli and C. Hart for critical reviews of the manuscript.
REFERENCES
1. Bednarik, D. P., and T. M. Folks. 1992. Mechanisms of HIV-1 latency.AIDS6:3-16.
2. Butera,S.T.,and T. M. Folks.1992.
Application
of latent HIV-1 infectedcellular models totherapeutic
intervention. AIDS Res. HumanRetroviruses 8:991-995.3. Butera,S.T.,V. L.Perez,B.-Y.Wu,G.J.Nabel,and T. M. Folks. VOL.68, 1994
on November 9, 2019 by guest
http://jvi.asm.org/
1991. Oscillation of the human immunodeficiency virus surface receptoris regulated by the state of viral activation inaCD4+cell model ofchronic infection.J.Virol.65:4645-4653.
4. Butera, S.T., B. D. Roberts, and T. M. Folks.1993.Regulationof HIV-1 expression by cytokine networks in a CD4+ model of chronic infection. J. Immunol. 150:625-634.
5. Clouse, K. A., D. Powell, I.Washington,G. Poli,K.Strebel,W. Farrar, B.Barstad, J. Kovacs, A.S.Fauci, and T. M. Folks. 1989. Monokineregulation ofHIV-1expressionin achronically infected human T-cell clone. J. Immunol. 142:431-438.
6. Embretson, J., M. Zupancic, J. L. Ribas,A.Burke, P.Racz, K. Tenner-Racz, and A. T. Haase.1993. Massive covertinfection of helper T lymphocytes and macrophages by HIV during the incubationperiodof AIDS. Nature(London)362:359-362. 7. Fauci, A. S., S. M. Schnittman, G. Poli, S. Koenig, and G.
Pantaleo. 1991.Immunopathogenic mechanismsin human immu-nodeficiencyvirus (HIV) infection.Ann. Intern. Med. 114:678-693.
8. Feinberg, M. B., R. F. Jarrett, A. Aldovini, R. C. Gallo, and F. Wong-Staal. 1986. HTLV-III expression and production involve complexregulationatthe levels ofsplicingandtranslationofviral RNA. Cell46:807-817.
9. Felber, B. K., M. Hadzopoulou-Cladaras, C. Cladaras, and G. N. Pavlakis. 1989.Therevprotein ofHIV-1affects thestability and transportoftheviral mRNA. Proc. Natl.Acad. Sci.USA 86:1495-1499.
10. Folks, T., J. Justement, A. Kinter, C. A. Dinarello, andA. S. Fauci. 1987. Cytokine induced expression of HIV-1 in a chronically infected promonocytic cell line. Science238:800-802.
11. Kim, S., R. Byrn, J. Groopman, and D. Baltimore.1989.Temporal aspects ofDNAand RNAsynthesis during human immunodefi-ciency virus infection: evidence for differentialgeneexpression.J. Virol. 63:3708-3713.
12. Malim, M. H., and B. R. Cullen. 1991. HIV-1 structural gene expressionrequires the binding ofmultiplerev monomers tothe viralRRE:implicationsfor HIV-1 latency. Cell65:241-248. 13. Malim, M. H., J. Hauber, S. Le, J. V. Maizel, and B. R. Cullen.
1989. The HIV-1 rev transactivator acts through a structured target sequence to activate nuclear export of unspliced viral mRNA. Nature(London)338:254-257.
14. Michael, N. L., P. Morrow, J. Mosca, M. Vahey, D. S. Burke, and R. R.Redfield. 1990.Induction of humanimmunodeficiency virus type 1expressioninchronically infectedcellsis associated primar-ily withashift inRNAsplicingpatterns.J.Virol.65:1291-1303. 15. Muesing, M. A., D. H. Smith, C. D. Cabradilla, C. V. Benton, L. A.
Lasky, and C. J.Capon. 1985.Nucleic acidstructureand expres-sion of the human AIDS/lymphadenopathy retrovirus. Nature (London)313:450-458.
16. Ou, C.-Y., S. Kwok, S. W. Mitchell, D. H. Mack, J. J. Sninsky, J. W. Krebs, P.Feorino, D. Warfield, and G. Schochetman. 1988. DNA amplificationfordirect detection of HIV-1 in DNA of peripheral bloodmononuclearcells. Science 239:295-297.
17. Pantaleo, G., C. Graziosi, and A. S. Fauci. 1993. The immuno-pathogenesis ofhumanimmunodeficiencyvirus infection. N. Engl.
J.Med. 328:327-335.
18. Pantaleo, G.,C. Graziosi, J. F. Demarest, L.Butini, M. Montroni, C. H.Fox, J. M. Orenstein,D. P. Kotler,and A. S. Fauci. 1993. HIVinfection is active andprogressive in lymphoid tissue during theclinically latent stage of disease. Nature (London) 362:355-358.
19. Perez, V. L., T. Rowe, J. S.Justement, S. T. Butera, C. H. June, and T. M. Folks. 1991. An HIVinfectedTcell clone defective in IL-2 productionand Ca"+ mobilization following CD3 stimulation.J. Immunol. 147:3145-3148.
20. Pomerantz, R., T. Seshamma, and D. Trono. 1992. Efficient replication of human immunodeficiency virus type 1 requires a threshold level ofrev:potentialimplications for latency. J. Virol. 66:1809-1813.
21. Pomerantz,R., D. Trono, M. B.Feinberg,and D. Baltimore. 1990. Cells nonproductively infected with HIV-1 exhibit an aberrant patternof viralRNAexpression:amolecular model for latency. Cell 61:1271-1276.
22. Pomerantz, R.J., 0. Bagasra,and D. Baltimore. 1992. Cellular latency of human immunodeficiency virus type-1. Curr. Opin. Immunol. 4:475-480.
23. Rogers, M. F., C.-Y. Ou, M. Rayfield, P. A. Thomas, E. E. Schoenbaum, E. Abrams, K. Krasinski, P. A. Selwyn, J. Moore, A. Kaul, K. T.Grimm, M.Bamji, G. Schochetman, and the New York City Collaborative Study of Maternal HIV Transmission and Montefiore Medical Center HIV Perinatal Transmission Study Group. 1989. Use of the polymerase chain reaction for early detection of theproviralsequencesof human immunodeficiency virus in infants born to seropositive mothers. N. Engl. J. Med. 320:1649-1654.
24. Seshamma, T., 0. Bagasra, D. Trono, D. Baltimore, and R. J. Pomerantz. 1992. Blocked early-stage latency in the peripheral blood cells of certain individuals infected with human immuno-deficiency virus type 1. Proc. Natl. Acad. Sci. USA 89:10663-10667.
25. Shaw,G. M., B. H.Hahn,S. K.Arya,J. E.Groopman,R.C.Gallo, and F. Wong-Staal. 1984. Molecular characterization ofhuman T-cell leukemia (lymphotropic) virus type III in the acquired immunedeficiency syndrome. Science226:1165-1171.
26. Strohman, R. C., P. S.Moss, J.Micou-Eastwood, D.Spector,A. Przybyla, and B. Paterson. 1977. Messenger RNA for myosin polypeptides: isolation from single myogenic cell cultures. Cell 10:265-273.
27. Trono,D., and D.Baltimore.1990.Ahuman cell factor is essential forHIV-1 Revaction. EMBOJ.9:4155-4160.
28. Vaishnav,Y.N., M.Vaishnav,and F.Wong-Staal. 1991. Identifi-cation andcharacterization of a nuclear factor that specifically bindstotherevresponseelement(RRE) of human immunodefi-ciency virustype 1 (HIV-1).NewBiol. 3:142-150.
29. Willey,R.L., D. H.Smith, L. A.Lasky,T. S.Theodore, P. L. Earl, B.Moss, D. J.Capon, and M. A. Martin. 1988. In vitro mutagen-esisidentifies a regionwithin the envelope gene of the human immunodeficiency virus that is critical for infectivity. J. Virol. 62:139-147.