0022-538X/98/$04.00
1
0
Copyright © 1998, American Society for Microbiology
Human Immunodeficiency Virus Type 1 Subtype F Reverse
Transcriptase Sequence and Drug Susceptibility
CRISTIAN APETREI,
1,2* DIANE DESCAMPS,
1GILLES COLLIN,
1IBTISAM LOUSSERT-AJAKA,
1FLORENCE DAMOND,
1MIHAI DUCA,
2FRANC¸OIS SIMON,
1AND
FRANC¸OISE BRUN-VE´ZINET
1Laboratoire de Virologie, Hoˆpital Bichat-Claude Bernard, 75018 Paris, France,
1and Virus Laboratory,
Microbiology Department, School of Medicine, Gr. T. Popa University, 6600 Iasi, Romania
2Received 21 August 1997/Accepted 15 January 1998
We sequenced and phylogenetically analyzed the reverse transcriptase (RT) regions of the pol genes of 14
human immunodeficiency virus type 1 (HIV-1) isolates from Romanian patients, which were classified as
sub-type F on the basis of env gene structure. The RT sequences showed that the strains clustered phylogenetically
and were equidistant from other HIV-1 subtypes as shown by the neighbor-joining and maximum-likelihood
methods, allowing us to define HIV-1 subtype F according to the pol classification. The subtype F RT sequences
differed from reported group M RT sequences by 10.94% (for nucleotides) and 7.6% (for amino acids).
Pheno-typic analysis of subtype F susceptibility to three classes of antiretroviral compounds showed an increase in the
50% inhibitory concentration of the tetrahydroimidazo[4,5,1-jk][1,4]-benzodiazepin-2-(1H)-one and -thione
(TIBO) derivate R82913 for one strain which was naturally resistant to this compound. This first report of
subtype F pol sequences confirms the perfect correlation between the phylogenetic positions determined by env
and pol analyses and suggests that virus variability might influence the efficacy of antiretroviral treatments.
This finding warrants a global evaluation of the phenotypic and genotypic susceptibility of HIV-1 subtypes to
antiretroviral drugs.
Most human immunodeficiency virus type 1 (HIV-1) drug
susceptibility studies have involved subtype B. Little
informa-tion on the impact of viral diversity on natural susceptibility
to antiretroviral drugs has been reported to date. However,
HIV-1 group O viruses are naturally resistant to nonnucleoside
reverse transcriptase (RT) inhibitors (8), as is HIV-2 (28).
Subtypes are defined on the basis of the env (24, 25) or gag (19)
gene. Most RT sequences reported to date belong to subtype
B strains, which prevail in North America and western Europe
(25), i.e., regions where antiretroviral drugs are developed and
clinical trials are conducted. RT sequences of subtypes A to
D are also available (25, 32, 34). The full sequences of Thai
strains defined as subtype E according to env classification and
defined as subtype A on the basis of the gag sequence (19) also
corresponded to subtype A on the basis of the pol gene (5, 14).
By contrast, full sequence analysis of a subtype G strain ruled
out recombination events in the pol gene (6), in keeping with
a previous report on the RT sequences of subtype G (17). To
date, no information has been published on the pol genes of
the other HIV-1 group M subtypes.
We present the first RT gene sequences and data on the
phenotypic susceptibility to antiretroviral drugs of subtype F
strains from Romanian patients. This is the dominant subtype
in Romanian children and adults (1, 11) and is also a minor
viral form in other countries, such as Brazil (23), Argentina (4),
Cameroon (27), Russia (18), Taiwan (7), Martinique (10),
Cy-prus (16), France (33), Belgium (13), and The Netherlands
(20). Recombinant F/B strains have also been reported (21, 30).
MATERIALS AND METHODS
Study population.We studied 14 HIV-1 subtype F strains isolated from
Ro-manian children (n59) and adults (n55). All but one of the infected children
were nosocomially infected (by injections with nonsterile, reused needles and syringes); the remaining one was vertically infected. Clinical and epidemiological data are described elsewhere, together with virus isolation, env subtype deter-mination, and strain codification (1). None of the patients had received antiret-roviral therapy.
HIV-1 RT sequencing. DNA was extracted with phenol-chloroform from
cocultured peripheral blood mononuclear cells from (PBMC) infected patients, precipitated with ethanol, and quantified spectrophotometrically. The pol gene was then amplified in a nested PCR with outer primers RT-18 and RT out and inner primers RT-19 and RT-20 as previously described (26).
Each nested-PCR product (1,008 bp) was subjected to direct population
se-quencing with sense primer A20(59-ATTTTCCCATTAGTCCTATT-39) and
antisense primer NE120(59-ATGTCATTGACAGTCCAGCT-39). Sequencing
reactions were run with the ABI Prism Dye Terminator Cycle Sequencing Ready Reaction kit with AmpliTaq DNA polymerase (FS; Perkin-Elmer) on an auto-mated sequencer (Applied Biosystems 373A).
Phylogenetic analysis.DNA sequences were analyzed with the
multiple-se-quence editor Clustal W (35) and improved by visual inspection. The semultiple-se-quences were gapstripped and a pairwise matrix based on 591 sites was generated with the DNADIST program of the PHYLIP package, version 3.56 (12). Tree topology was inferred by the neighbor-joining method with the Kimura two-parameter distance matrix (PHYLIP) and a transition/transversion ratio of 2. Bootstrap analysis was performed with the SEQBOOT (100 resamplings), DNADIST, NEIGHBOR, and CONSENSE programs (PHYLIP package). Phylogenetic analysis was also performed by the maximum-likelihood method, using the DNAML program (12). The tree outliers were the HIV-1 group O sequences (MVP5180 and ANT70). In the tree construction we also included the RT sequences of six subtype B HIV-1 isolates (LAI, SF2, MN, OYI, JRFL, and JRCSF), three subtype D sequences (ELI, NDK, and Z2Z6), two subtype A sequences (UG037 and U455), the sequence of an A/G recombinant strain (IBNG) (14), and the sequence of a presumed A/D recombinant strain (MAL) from the Los Alamos National Laboratory database (25). The newly reported sequences for strains CM240 (5), 90CR402, and 93TH253 (14), recombinant A/E strains, were also used in the phylogenetic analyses.
Phenotypic susceptibility assay.The phenotypic susceptibilities of the cellular
HIV-1 subtype F isolates were analyzed in a PBMC assay, taking into account the replication kinetics of each strain (3). After conventional isolation of HIV from frozen PBMC, the cell-free HIV-1 subtype F supernatants corresponding to peak
RT activity were serially diluted (100to 1024) and incubated with fresh normal
phytohemagglutinin-stimulated PBMC. After being washed, the infected cells were placed in 96-well plates containing six serial dilutions of the antiretroviral
* Corresponding author. Mailing address: Laboratoire de Virologie,
Hoˆp. Bichat-Claude Bernard, 46 Rue Henri Huchard, 75018 Paris,
France. Phone: 33 1 40 25 88 94. Fax: 33 1 46 27 02 08. E-mail: francois
[email protected].
3534
on November 9, 2019 by guest
http://jvi.asm.org/
drugs. Each dilution was tested in triplicate. On days 5, 7, and 10, the superna-tants were collected and half the medium was replaced with fresh drug-contain-ing medium. The 50% tissue culture infective dose was assessed by measurdrug-contain-ing RT activity in control drug-free supernatants collected on the same days. At the peak
of RT activity we calculated the drug concentrations inhibiting 50 and 90% (IC50
and IC90, respectively) of the RT activity of 100 50% tissue culture infective
doses. For zidovudine (ZDV), an IC50cutoff of 0.05mM has been defined to
classify the virus isolates as ZDV sensitive or ZDV resistant, based on pheno-typic analyses of several isolates from treated and untreated patients and on comparisons of these results to genotypic data. For the other antiretroviral drugs, it has not been possible to determine any cutoff value. In this study, phenotypic
resistance to these compounds was defined as at least a fivefold increase in IC50s
for the HIV-1 subtype F isolates compared to those for HIV-1 subtype B strains.
Antiretroviral agents.We tested the nucleoside RT inhibitors ZDV
(Well-come, Dartford, United Kingdom) and lamivudine (3TC; Glaxo-Well(Well-come, Dartford, United Kingdom); the nonnucleoside RT inhibitors tetrahydroimi-dazo[4,5,1-jk][1,4]-benzodiazepin-2(1H)-one and -thione (TIBO) derivate R82913 (Janssen, Beerse, Belgium), delavirdine (DLV; Upjohn, Kalamazoo, Mich.), and nevirapine (NVP; Boehringer Ingelheim Pharmaceuticals, Ridge-field, Conn.); and the protease inhibitors saquinavir (SQV; Roche, Welwyn Garden City, United Kingdom) and ritonavir (RTV; Abbott, Abbott Park, Ill.). The purified drugs were kindly provided by the manufacturers.
Nucleotide sequence accession numbers.The nucleotide and amino acid
se-quences of codons 33 to 235 of the RT genes and proteins from the 14 HIV-1 subtype F isolates have been submitted to GenBank (accession no. Y16138 to Y16151).
RESULTS
Genetic analysis.
Figure 1 shows the amino acid alignment
of codons 33 to 235 of the RT genes of the different subtype F
isolates. The subtype F consensus sequence differed from that
of subtype B in 11 residues (V35T, T39A, E40D, D131E,
I135L, S162Y, K173T, Q174K, T200A, Q207A, and R211K).
Two of these mutations (A200 and K211) have been reported
to occur in subtype B isolates. Although not yet reported for
subtype B isolates, one residue (D40) of the subtype F
con-sensus sequence has been described as occurring in concon-sensus
sequences of subtype A. The sequence T173-K174 has been
described for the IBNG subtype A/G isolate (25). Residues
A39, E131, L135, and Y162, which are frequently present in
subtype F strains, have never been encountered in other HIV-1
group M subtypes. None of the sequences analyzed contained
mutations previously linked to resistance to nucleoside or
non-nucleoside RT inhibitors. The sequence of the RO-BCI23
iso-late was more similar to that of the subtype B consensus
se-quence in the first part of the RT gene (subtype B structure in
sequences encoding T39, E40, and I135) but was more variable
than the other subtype F isolates in the second half of the RT
gene (close to the binding pocket). Compared to the subtype B
consensus sequence, the RO-BCI23 protein bore four
muta-tions close to the active site of the RT protein (E169D, K173A,
Q174K, and D177E).
At position 60, all but one of the strains originating in
nosocomially infected Romanian children (BCI7,
BCI8, BCI9, BCI11, BCI12, BCI15, and
RO-BCI16) bore a valine (as in the subtype B consensus sequence),
whereas the strains isolated from adults (BCI17,
RO-BCI18, RO-BCI19, RO-BCI20, and RO-BCI23), a vertically
infected child (RO-BCI13), and a nosocomially infected child
from whom the strain was isolated in 1994 (RO-BCI1) bore an
isoleucine. Conversely, at position 39, all but one (RO-BCI17)
of the strains isolated from adults bore a threonine, like
sub-type B strains, whereas all those isolated from nosocomially
infected children bore an alanine.
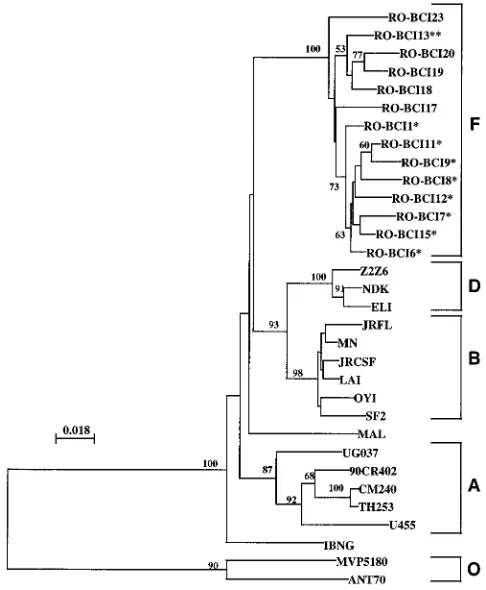
Phylogenetic analysis of HIV-1 RT sequences.
Phylogenetic
analysis of the RT nucleotide sequences by the
neighbor-join-ing and maximum-likelihood methods gave similar results.
FIG. 1. Alignment of the deduced amino acid sequences of HIV-1 subtype F RT and the consensus (Cons) sequences of HIV-1 group M and group O strains. Strains RO-BCI17 to RO-BCI23 were isolated from HIV-1-infected adults.p, established following the analysis of the pol sequences originating from 11 Zimbabwean seroconverters (32).pp, naturally resistant to TIBO derivate.on November 9, 2019 by guest
http://jvi.asm.org/
Phylogenetic trees were constructed by using the 14 Romanian
HIV-1 RT sequences, representative RT sequences of subtype
B strains, and sequences belonging to different subtypes
ob-tained from the database (25). Figure 2 shows the phylogenetic
tree constructed by neighbor joining. High bootstrap values
were obtained at the relevant nodes, indicating that subtypes
B, D, and F each form a consistent clade. Subtype A was
composed of five different strains defined as A or E on the
basis of the env classification, whereas recombinant strains
IBNG and MAL were subtype outliers.
The subtype F phylogenetic tree showed that the strains
from nosocomially infected Romanian children formed a
clus-ter, as was previously observed by analyzing the env sequence
(1). The RT sequence of a strain from a vertically infected
child (RO-BCI13) clustered with the adult sequences. The RT
sequence of strain RO-BCI23 formed a separate branch within
the subtype F tree.
The average intrasubtype F sequence divergence (597
nu-cleotides) was 3.56% (range, 1.54 to 6.36%), and the distance
between the subtype F sequences and the sequences of
oth-er subtypes belonging to group M was 10.94% (range, 8.04
to 13.5%) (P
,
0.001) (Table 1). The divergence among the
subtype F amino acid sequences (197 residues) was 3.7% (range,
1.57 to 6.7%), whereas the mean distance between the subtype
F RT amino acid sequences and other group M sequences was
7.6% (range, 6.00 to 11.59%).
Phenotypic susceptibility.
The IC
50s of nucleoside analogs
ZDV and 3TC were similar to those for wild-type subtype B
field isolates. Although the IC
50s were higher for the subtype F
strains than for the subtype B strains, all the subtype F isolates
were susceptible to protease inhibitors. All were also sensitive
to the nonnucleoside RT inhibitors NVP and DLV.
Suscepti-bility to the third nonnucleoside RT inhibitor, TIBO derivate
R82913, was lower for two isolates (Table 2); one isolate
(RO-BCI1) showed borderline susceptibility, with a moderate
in-crease in IC
50and IC
90(0.30 and 1.1
mM, respectively), the
second strain (RO-BCI23) showed a significant increase in
both IC
50and IC
90(0.53 and 2.02
mM, respectively). This
phenotype was not associated with any of the known mutations
linked to TIBO resistance. However, the RO-BCI23 isolate
showed the most variable RT sequence of the subtype F
iso-lates and presented different residues close to the active site of
the RT. We therefore assessed the phenotypic susceptibilities
of all the available subtype F isolates to TIBO. The TIBO IC
50(mean
6
standard deviation) for 12 subtype F isolates was
0.07
6
0.06
mM (range, 0.01 to 0.2
mM), and the IC
90was 0.4
6
0.27
mM (range, 0.01 to 0.88
mM), meaning that all 12 strains
were susceptible.
DISCUSSION
Studies of HIV’s genetic diversity have shown different rates
of variability for the different viral genes, the most conserved
structural gene being pol (29, 31). Analysis of the pol gene was
not considered relevant for genotyping (36), but several recent
reports have shown that the numerous selection pressures on
the pol gene, generally reflected by synonymous substitutions,
make it suitable for phylogenetic studies (32, 34). The different
pol subtypes described so far correspond to env or gag subtypes
(18, 25, 32, 34).
[image:3.612.49.292.69.364.2]We analyzed the RT coding regions of 14 isolates originating
in different parts of Romania and characterized as subtype F
on the basis of the env sequence (1). Based on a pol nucleotide
and amino acid sequence comparison, these strains clustered
phylogenetically and were equidistant from other HIV-1
sub-types. This first report of subtype F pol sequences confirms the
perfect correlation between the phylogenetic positions
deter-mined by env and gag analysis. The similar results obtained by
the neighbor-joining and maximum-likelihood methods
sup-port the reliability of the phylogenetic tree of HIV-1 RT
se-quences. Furthermore, phylogenetic analysis of the pol gene of
Romanian HIV-1 isolates revealed a cluster similar to that
obtained by analyzing the env gene sequences. The sequences
of isolates from nosocomially infected children formed a
sep-arate branch within the Romanian sequence cluster with a high
FIG. 2. Phylogenetic analysis comparing the RT regions of HIV-1 pol genesfrom different strains. Tree topology was inferred by the neighbor-joining meth-od. The tree was based on an alignment of nucleotides from which columns containing gaps have been deleted (597 nucleotides). The tree was rooted with HIV-1 group O sequences. The numbers given at the branch points are the 50% threshold majority consensus values for 100 bootstrap replicates. Vertical dis-tances are given for clarity. The cluster of sequences from nosocomially infected children (p) had already been observed by analyzing the env gene (1). Strain RO-BCI13 (pp) was isolated from a vertically infected child. Strains RO-BCI17, RO-BCI18, RO-BCI19, RO-BCI20, and RO-BCI23 were isolated from HIV-1-infected adults.
TABLE 1. Nucleotide and amino acid divergences between subtype
F and other group M subtype and group O RT sequences
Isolate group % Divergence from subtype F (range) a
Nucleotides Amino acids
Group M
10.94 (8.04–14.84)
7.6 (6.00–11.59)
Subtype B
10.04 (8.04–13.15)
7.48 (6.00–9.96)
Subtype A
11.98 (9.35–14.84)
7.63 (6.09–10.23)
Subtype D
11.01 (9.55–11.96)
8.36 (6.95–11.59)
Group O
32.09 (29.9–33.52)
19.09 (16.49–21.92)
aAverage intrasubtype F divergence was 3.56% (range, 1.54 to 6.36%) for nucleotides and 3.7% (range, 1.57 to 6.7%) for amino acids.
on November 9, 2019 by guest
http://jvi.asm.org/
[image:3.612.308.549.620.709.2]bootstrap value (i.e., 73), which supports our hypothesis of a
unique introduction of HIV in horizontally infected children
from Romania, probably resulting from the use of an infected
blood product (1).
None of the patients in this study had ever received
antiret-roviral therapy. To investigate the impact of viral variability on
susceptibility to antiretroviral drugs, we analyzed the
pheno-typic susceptibilities of four subtype F isolates to different
classes of antiretroviral drugs. All but two of the strains had
drug susceptibilities similar to those of wild-type subtype B
field isolates in the same assay (Table 2). One isolate
(RO-BCI1) showed borderline susceptibility, i.e., there was a
mod-erate increase in both the IC
50and IC
90of a nonnucleoside RT
inhibitor, TIBO, whereas another isolate (RO-BCI23) showed
significantly diminished susceptibility to TIBO. Although this
compound is not used to treat HIV infection, the phenotypic
susceptibility analysis showed that diversity within group M
might affect drug susceptibility. Nonnucleoside RT inhibitors
are a promising class of antiretroviral compounds for
com-bined therapy because of their low toxicity, facility of use, and
reasonable cost. However, the use of these agents might be
hindered by the existence of naturally resistant variants. HIV-1
group O isolates are naturally resistant to nonnucleoside RT
inhibitors, as is HIV-2 (9). Because point mutations can lead to
marked reductions in drug susceptibility (22), intra- or
inter-subtype pol diversity might be reflected by particular drug
susceptibility profiles. HIV-1 subtype F is only a minor form of
HIV-1 but is widely distributed (4, 7, 10, 13, 16, 18, 20, 21, 23,
27, 33), with major circulation in Romania (1, 11, 15).
Phylo-genetic studies of isolated strains from different countries have
revealed that separate epidemiological events contributed to
the worldwide distribution of this subtype (1, 2, 16, 18). Our
study revealed a natural reduction in susceptibility to a
non-nucleoside RT inhibitor in HIV-1 subtype F strains and
rein-forces the need for global screening for HIV-1 group M
iso-lates by sequencing and phenotyping susceptibility analysis.
ACKNOWLEDGMENTS
This work was supported by Agence Nationale de Recherches sur le
SIDA (ANRS), France (grant 96009). C.A. is an ANRS postdoctoral
fellow. The samples were obtained with the support of the Romanian
Association against AIDS.
REFERENCES
1. Apetrei, C., I. Loussert-Ajaka, G. Collin, F. Letourneur, M. Duca, S.
Sara-gosti, F. Simon, and F. Brun-Ve´zinet.1997. HIV type 1 subtype F sequences
in Romanian children and adults. AIDS Res. Hum. Retroviruses 13:363–365. 2. Bandea, C. I., A. Ramos, D. Pienazek, F. R. Pascu, A. Tanuri, G.
Schochet-man, and M. A. Rayfield.1995. Epidemiologic and evolutionary relationships
between Romanian and Brazilian HIV-1 subtype F strains. Emerg. Infect. Dis. 1:91–93.
3. Brun Ve´zinet, F., D. Ingrand, L. Deforges, K. Gochi, F. Ferchal, M. Schmitt,
M. Jung, B. Masquelier, J. Aubert, C. Buffet-Janvresse, and H. Fleury.1991.
HIV-1 sensitivity to zidovudine: a consensus culture technique validated by genotypic analysis of the reverse-transcriptase. J. Virol. Methods 37:177–188. 4. Campodonico, M., W. Janssens, L. Heyndrickx, K. Fransen, A. Leonaers,
F. F. Fay, M. Taborda, G. van der Groen, and O. H. Fay.1996. HIV type 1
subtypes in Argentina and genetic heterogeneity of the V3 region. AIDS Res. Hum. Retroviruses 12:79–81.
5. Carr, J. K., M. O. Salminen, C. Koch, D. Gotte, A. W. Artenstein, P. A.
Hegerich, D. St. Louis, D. S. Burke, and F. E. McCutchan.1996. Full-length
sequence and mosaic structure of a human immunodeficiency virus type 1 isolate from Thailand. J. Virol. 70:5935–5943.
6. Carr, J. K., M. O. Salminen, P. Lenikki, B. Johansson, D. S. Burke, and F. E.
McCutchan.1996. First full length sequence prototypes of HIV-1 clades C,
E and G: two of three are mosaic genomes, abstr. TuA 100. XIth Interna-tional Conference on AIDS.
7. Chang, K. S. S., C. I. Lin, J. H. Chen, C. H. Shih, H. C. Lin, S. C. Twu, and
M. O. Salminen.1997. HIV type 1 env gene diversity detected in Taiwan.
AIDS Res. Hum. Retroviruses 13:201–204.
8. Descamps, D., G. Collin, I. Loussert-Ajaka, S. Saragosti, F. Simon, and F.
TABLE
2.
IC
50s
and
IC
90s
of
nucleoside
and
nonnucleoside
RT
inhibitors
and
protease
inhibitors
of
HIV-1
subtype
F
isolates,
reference
HIV-1
isolates,
and
HIV-2
Isolate ZDV 3TC DLV NVP TIBO SQV RTV IC 50 ( m M) IC 90 ( m M) IC 50 ( m M) IC 90 ( m M) IC 50 ( m M) IC 90 ( m M) IC 50 ( m M) IC 90 ( m M) IC 50 ( m M) IC 90 ( m M) IC 50 (nM) IC 90 (nM) IC 50 (nM) IC 90 (nM) HIV-1 subtype B a , 0.03 , 0.03 0.05 6 0.03 0.23 6 0.1 0.02 6 0.01 0.10 6 0.07 0.04 6 0.01 0.22 6 0.01 0.09 6 0.05 0.44 6 0.12 11 6 21 9 6 55 5 6 26 150 6 57 HIV-2 ROD , 0.03 0.20 0.02 0.04 . 62.5 . 62.5 . 20 . 20 . 6.25 . 6.25 10 19 80 433 HIV-1 group O b , 0.03 0.04 6 0.03 0.03 6 0.02 0.16 6 0.13 15.5 6 13 31 6 29 , 0.02– . 20 , 0.02– . 20 5.4 6 2.1 5.6 6 1.7 15 6 42 6 6 6 108 6 79 453 6 379 HIV-1 subtype F RO-BCI1 , 0.03 0.05 0.08 0.21 , 0.10 , 0.10 0.03 0.11 0.3 1.10 21 35 65 239 RO-BCI16 , 0.03 0.01 0.04 0.16 , 0.10 0.16 0.02 0.12 0.17 0.27 21 33 108 309 RO-BCI18 , 0.03 0.02 0.04 0.16 , 0.10 , 0.10 0.03 0.13 0.16 0.57 36 48 165 480 RO-BCI23 , 0.03 0.03 0.03 0.10 , 0.10 , 0.10 , 0.002 0.005 0.53 2.02 14 26 109 295 a IC 50 s and IC 90 s were measured for isolates from untreated patients (n 5 25). b Performed on 11 HIV-1 group O isolates as previously described (9).on November 9, 2019 by guest
http://jvi.asm.org/
Brun-Ve´zinet.1995. HIV-1 group O sensitivity to antiretroviral drugs. AIDS
8:977–978.
9. Descamps, D., G. Collin, F. Letourneur, C. Apetrei, F. Damond, I.
Loussert-Ajaka, F. Simon, S. Saragosti, and F. Brun-Ve´zinet.1997. Susceptibility of
human immunodeficiency virus type 1 group O isolates to antiretroviral agents: in vitro phenotypic and genotypic analyses. J. Virol. 71:8893–8898. 10. Desgranges, C., S. Fillon, G. Audoly, C. Neisson-Vernant, O. Bera, M. Ouka,
R. Cesaire, L. Buzelay, and F. Barin.1996. Presence of HIV-1 subtypes B
and F and HTLV-I in HIV/HTLV coinfected individuals of Martinique. J. Acquir. Immune Defic. Syndr. Hum. Retrovirol. 13:468–470.
11. Dumitrescu, O., M. L. Kalish, S. C. Kilks, C. I. Bandea, and J. A. Levy. 1994. Characterization of human immunodeficiency virus type 1 isolates from children in Romania: identification of a new envelope subtype. J. Infect. Dis.
169:281–288.
12. Felsenstein, J. 1993. PHYLIP (phylogeny interference package), version 3.5c. Department of Genetics, University of Washington, Seattle. 13. Fransen, K., A. Buve´, J. N. Nkengasong, M. Laga, and G. van der Groen.
1996. Longstanding presence in Belgians of multiple non-B HIV-1 subtypes. Lancet 347:1403.
14. Gao, F., D. L. Robertson, S. G. Morrison, H. Hui, S. Craig, J. Decker, P. N.
Fultz, M. Girard, G. M. Shaw, B. H. Hahn, and P. M. Sharp.1996. The
heterosexual human immunodeficiency virus type 1 epidemic in Thailand is caused by an intersubtype (A/E) recombinant of African origin. J. Virol. 70: 7013–7029.
15. Holm-Hansen, C., D. Grothues, S. Rusad, B. Rosok, F. R. Pascu, and B.
Asjo¨.1995. Characterization of HIV type 1 from Romanian children: lack of
correlation between V3 loop amino acid sequence and syncytium formation in MT-2 cells. AIDS Res. Hum. Retroviruses 11:597–603.
16. Kostrikis, L. G., E. Bagdades, Y. Cao, L. Zhang, D. Dimitriou, and D. D. Ho. 1995. Genetic analysis of human immunodeficiency virus type 1 strains from patients in Cyprus: identification of a new subtype designated subtype I. J. Virol. 69:6122–6130.
17. Leitner, T., D. Escanilla, S. Marquina, J. Wahlberg, C. Brostro¨m, H. B.
Hansson, M. Uhle´n, and J. Albert.1995. Biological and molecular
charac-terization of subtype D, G and A/D recombinant HIV-1 transmission in Sweden. Virology 209:136–146.
18. Leitner, T., G. Korovina, S. Marquina, T. Smolskaya, and J. Albert. 1996. Molecular epidemiology and MT-2 tropism of Russian HIV type 1 variants. AIDS Res. Hum. Retroviruses 12:1595–1603.
19. Louwagie, J., F. E. McCutchan, M. Peeters, T. P. Brennan, E. Sanders-Buell,
G. A. Eddy, G. van der Groen, K. Fransen, G. D. Gershy-Damet, R. Deleys,
and D. S. Burke.1993. Phylogenetic analysis of gag genes from 70
interna-tional HIV-1 isolates provides evidence for multiple genotypes. AIDS 7: 769–780.
20. Lukhasov, V. V., C. L. Kuiken, K. Boer, and J. Goudsmit. 1996. HIV type 1 subtype in The Netherlands circulating among women originating from AIDS-endemic regions. AIDS Res. Hum. Retroviruses 12:951–953. 21. Marquina, S., T. Leitner, R. D. Rabinovich, J. Benetucci, O. Libonatti, and
J. Albert.1996. Coexistence of subtypes B, F and a B/F env recombinant of
HIV type 1 in Buenos Aires, Argentina. AIDS Res. Hum. Retroviruses 12: 1651–1654.
22. Mellors, J. W., R. F. Schinazi, and B. A. Larder. 1996. Mutations in retroviral genes associated with drug resistance, p. III-206–III-241. In G. Myers, B. Foley, J. W. Mellors, B. Korber, K. T. Jeang, and S. Wain-Hobson (ed.), Human retroviruses and AIDS 1996. A compilation and analysis of nucleic
acid and amino acid sequences. Los Alamos National Laboratory, Los Alamos, N.Mex.
23. Morgado, M. G., E. C. Sabino, E. G. Sapaer, V. Bongertz, L. Brigido,
M. D. C. Guimaraes, E. A. Castilho, B. Galvao-Castro, J. I. Mullins, R. M.
Hendry, and A. Mayer.1994. V3 region polymorphism in HIV-1 from Brazil:
prevalence of subtype B strains divergent from North American/European prototype and detection of subtype F. AIDS Res. Hum. Retroviruses 10: 569–576.
24. Myers, G. 1994. HIV: between past and future. AIDS Res. Hum. Retrovi-ruses 10:1317–1324.
25. Myers, G., B. Foley, J. W. Mellors, B. Korber, K. T. Jeang, and S.
Wain-Hobson (ed.).1996. Human retroviruses and AIDS 1996. A compilation and
analysis of nucleic acid and amino acid sequences. Los Alamos National Laboratory, Los Alamos, N.Mex.
26. Nihjhuis, M., C. A. B. Boucher, and R. Schuurman. 1995. Sensitive proce-dure for the amplification of HIV-1 RNA using combined reverse-transcrip-tion and amplificareverse-transcrip-tion reacreverse-transcrip-tion. BioTechniques 19:178–180.
27. Nkengasong, J. N., W. Janssens, L. Heyndrickx, K. Fransen, P. M. Ndumbe,
J. Motte, A. Leonaers, M. Ngolle, J. Ayuk, P. Piot, and G. van der Groen.
1994. Genotypic subtypes of HIV-1 in Cameroon. AIDS 8:1405–1412. 28. Pauwels, R., K. Andries, J. Desmyter, D. Schols, M. Kukla, H. Breslin, A.
Raeymaeckers, J. Van Gelder, R. Woestenborghs, J. Heykants, K.
Schelle-kens, M. Janssen, E. De Clercq, and P. Janssen.1990. Potent and selective
inhibition of HIV-1 replication in vitro by a novel series of TIBO derivatives. Nature 343:470–474.
29. Quinones-Mateu, M. E., A. Holguin, J. Dopazo, I. Najera, and E. Domingo. 1996. Point mutation frequencies in the pol gene of human immunodefi-ciency virus type 1 are two- to threefold lower than those of env. AIDS Res. Hum. Retroviruses 12:1117–1128.
30. Sabino, E. C., E. G. Shpaer, M. G. Morgado, B. T. M. Korber, R. S. Diaz, V.
Bongertz, S. Cavalcante, B. Galva˜o-Castro, J. I. Mullins, and A. Mayer.
1994. Identification of human immunodeficiency virus type 1 envelope genes recombinant between subtypes B and F in two epidemiologically linked individuals from Brazil. J. Virol. 68:6340–6346.
31. Seibert, S. A., C. Y. Howell, M. K. Hughes, and A. L. Hughes. 1995. Natural selection on the gag, pol, and env genes of human immunodeficiency virus 1 (HIV-1). Mol. Biol. Evol. 12:803–813.
32. Shafer, R. W., J. A. Eisen, T. C. Merigan, and D. A. Katzenstein. 1997. Sequence and drug susceptibility of subtype C reverse transcriptase from human immunodeficiency virus type 1 seroconverters in Zimbabwe. J. Virol.
71:5441–5448.
33. Simon, F., I. Loussert-Ajaka, F. Damond, S. Saragosti, F. Barin, and F.
Brun-Ve´zinet.1996. HIV type 1 diversity in Northern Paris, France. AIDS
Res. Hum. Retroviruses 12:1427–1433.
34. Soto-Ramirez, L. E., S. Tripathy, B. Renjifo, and M. Essex. 1996. HIV pol sequences from India fit distinct subtype pattern. J. Acquir. Immune Defic. Syndr. Hum. Retrovirol. 13:299–307.
35. Thompson, J., D. Higgins, and T. Gibson. 1994. CLUSTAL W: improving the sensitivity of progressive multiple sequence alignment through sequence weighting, position-specific gap penalties and weight matrix choice. Nucleic Acids Res. 22:4673–4680.
36. WHO Network for HIV Isolation and Characterization. 1994. HIV type 1 variation in World Health Organization-sponsored vaccine evaluation sites: genetic screening, sequence analysis, and preliminary biological character-ization of selected viral strains. AIDS Res. Hum. Retroviruses 10:1327–1343.