Iron-sulfur cluster protein NITROGEN FIXATION S-LIKE 1 and its interactor
1
FRATAXIN function in plant immunity
2
Jose Pedro Fonseca1, Hee-Kyung Lee1, Clarissa Boschiero1, Marcus Griffiths1, Seonghee Lee1,2,
3
Patrick Zhao1, Larry M. York1, Kirankumar S. Mysore1†
4
5
1 Noble Research Institute LLC, Ardmore, OK, USA 6
2 Current address: Gulf Coast Research and Education Center, University of Florida, Wimauma, 7
FL, USA
8
†Corresponding Author: [email protected] 9
10
Short title: Iron-sulfur cluster proteins function in defense
11
12
One-sentence summary: Iron-sulfur (Fe-S) cluster proteins play a role in plant innate immunity
13
by regulating plant defense-related genes.
14
15
16
17
18
19
20
21
22
23
AUTHOR CONTRIBUTIONS
24
JPF and KSM designed the research; JPF performed all experiments and analyzed data; JPF and
25
CB analyzed genomics data, PZ provided bioinformatics support; MG, LMY and JPF performed
26
and analyzed shoot respiration assays, JPF performed microscopy; HL and SL provided technical
27
support; JPF and KSM analyzed data and wrote the manuscript.
28
29
ABSTRACT
30
Iron-sulfur (Fe-S) clusters are inorganic cofactors that are present in all kingdoms of life as part
31
of a large number of proteins involved in several cellular processes, including DNA replication
32
and metabolism. In this work, we demonstrate an additional role for two Fe-S cluster genes in
33
biotic stress responses in plants. Eleven Fe-S cluster genes, including the NITROGEN FIXATION
34
S (NIFS)-LIKE 1 (NFS1) and its interactor FRATAXIN (FH), when silenced in Nicotiana
35
benthamiana, compromised nonhost resistance to Pseudomonas syringae pv. tomato T1.
36
NbNFS1 expression was induced by pathogens and salicylic acid. Arabidopsis thalianaatnfs and
37
atfh mutants, with reduced AtNFS1 or AtFH gene expression, respectively, showed increased
38
susceptibility to both host and nonhost pathogen infection. Arabidopsis AtNFS1 and AtFH
39
overexpressor lines displayed decreased susceptibility to infection by host pathogen P. syringae
40
pv. tomato DC3000. The AtNFS1 overexpression line exhibited constitutive upregulation of
41
several defense-related genes and enrichment of gene ontology terms related to immunity and
42
salicylic acid responses. Our results demonstrate that NFS1 and its interactor FH are involved
43
not only in nonhost resistance but also in basal resistance, suggesting a new role of the Fe-S
44
cluster pathway in plant immunity.
45
46
47
48
49
50
51
52
53
54
55
INTRODUCTION
57
Iron-sulfur (Fe-S) clusters are versatile cofactors formed by iron atoms and inorganic sulfide.
58
Several plant metabolic pathways and proteins require Fe-S clusters for proper functioning
59
(Johnson et al., 2005; Balk and Shaeddler, 2014). Biosynthesis of Fe-S clusters requires a
60
complex machinery. Until now, around 46 genes involved in Fe-S cluster biosynthesis have been
61
identified in Arabidopsis thaliana (Balk and Pilon, 2011). Fe-S clusters are generally linked to
62
the polypeptide chain via the interaction of the iron atoms with cysteine residues (Balk and
63
Lobreaux, 2005). Three types of Fe-S cluster biosynthetic systems have been described so far:
64
(1) the nitrogen fixation (NIF) system from azothropic bacteria; (2) the sulfur mobilization
65
(SUF) system found in plastids; and (3) the iron-sulfur cluster (ISC) system, commonly found in
66
bacteria and mitochondria of eukaryotic cells (Lill, 2009; Balk and Pilon, 2011). There are
67
common steps to all systems: first, the release of sulfur from cysteine to form alanine; second,
68
the transfer of sulfur to scaffold proteins where Fe-S clusters are assembled; the third and the last
69
step is transfer of the mature Fe-S cluster to an apoprotein. Each of these three steps are
70
mechanistically conserved by the requirement for a cysteine desulfurase (Jonhson et al., 2005).
71
The main function of Fe-S proteins is electron transfer through the different oxidation states of
72
iron that can generate redox potential (Balk and Lobreaux, 2005). Several Fe-S cluster proteins
73
contribute to electron transfer in the respiratory electron transport chain (Lill, 2009; Couturier et
74
al., 2013). They are also involved in other cellular processes, including redox and catalytic
75
activity through their enzymes (e.g. aconitase which can convert citrate to isocitrate; Tong and
76
Rouault, 2007) and regulatory proteins (Fe-S clusters are oxygen stress sensors in bacteria; Kiley
77
and Beinert, 2003). Several transcription factors in bacteria, such as iron-sulfur cluster regulator
78
(IscR) and nitrite-sensitive transcription repressor (NsrR), have Fe-S cluster cofactors that sense
79
changes in superoxide, Fe-S clusters and oxygen contents (Rouault , 2006). Defects in Fe-S
80
cluster protein biogenesis have a drastic impact in several metabolic pathways for all organisms,
81
including amino-acid biosynthesis, gene expression, DNA replication, DNA repair, protein
82
translation, amino acid metabolism, photosynthesis, respiration and development (Balk and
83
Schaedler, 2014; Beinert, 2000, Lill, 2009; Lill and Muhlenhoff, 2008; Rouault , 2006; Rouault ,
84
2012; Stehling and Lill, 2013). It has been shown in mammals that Thio modification of transfer
RNAs require the mitochondrial cysteine desulfurase, NITROGEN FIXATION S-LIKE 1
86
(NFS1), as a sulfur donor (Nakai et al., 2017; Braymer and Winge, 2018; Mueller et al., 2006).
87
In Arabidopsis, there are two NFS genes coding for cysteine desulfurases: a mitochondrial
88
(AtNFS1) and a chloroplastic one (AtNFS2), which share only 18% amino acid identity among
89
them. AtNFS1 and AtNFS2 both have cysteine desulfurase activity required for the mobilization
90
of sulfur from cysteine that is vital for the biosynthesis of Fe-S clusters in their respective
sub-91
cellular compartments (Frazzon et al., 2007; Leon et al., 2002; Turowski et al., 2012). However,
92
only the mitochondrial AtNFS1 was shown to be required for cytosolic Fe-S enzyme activity
93
(Bernard et al., 2013). The function of many Fe-S proteins, including AtNFS1, is essential for
94
normal plant development in Arabidopsis. Transfer (T)-DNA knockout mutants of AtNFS1 are
95
embryo lethal thus making it challenging for genetic analysis of AtNFS1 (Frazzon et al., 2007;
96
Couturier et al., 2013; Balk and Shaeddler, 2014). AtNFS1 is thought to provide sulfur for the
97
biogenesis of Fe-S clusters in plants (Balk and Lobreaux, 2005). It can catalyze the conversion of
98
cysteine to alanine, forming sulfane sulfur as a by-product (Balk and Pilon, 2011; Balk and
99
Lobreaux, 2005). Arabidopsis AtNFS1 overexpression lines have increased levels of Fe and S
100
while mutant plants had lower amounts of S further reinforcing the role of AtNFS1 in Fe and S
101
homeostasis (Armas et al., 2019). The Arabidopsis AtNFS1 protein has been expressed and
102
purified from Escherichia coli and is shown to catalyze the release of sulfide from cysteine
103
(Frazzon et al., 2007). Another study confirmed a cysteine desulfurase activity for AtNFS1 by
104
showing that it can catalyze L-alanine production in increasing L-cysteine concentrations
105
(Turowski et al., 2012). The same study also showed that AtNFS1 binds to an Arabidopsis
106
FRATAXIN homolog (AtFH) using in vitro pull down, and that addition of AtFH protein
107
increases the catalytic activity of AtNFS1 by three-fold. FH is a structurally well-conserved
108
protein across different kingdoms of life (Gibson et al., 1996). AtFH showed dual localization in
109
mitochondria and chloroplasts in plants and is known to bind to iron molecules and to be
110
involved in iron homeostasis, Fe-S cluster biosynthesis and mitochondrial energy metabolism
111
(Turowski et al., 2015). In yeast, Frataxin1 (YFH1) is associated with mitochondrial membranes
112
and is involved in mitochondrial iron homeostasis. The yeast yfh1 mutant has growth defects,
113
accumulates mitochondrial iron, exhibits a higher sensitivity to oxidative stress, and has reduced
114
mitochondrial respiration (Ristow et al., 2000). In humans, deficiency in FH function due to a
115
point mutation in the FH gene has been related to a degenerative disease known as Friedreich’s
ataxia that leads to premature death (Babcock et al., 1997). In Arabidopsis, atfh knockout
117
mutants were shown to be lethal, but a few AtFH knockdown lines are viable and showed
118
retarded growth, reduced fresh weight and seed number (Busi et al., 2006; Balk and Shaeddler,
119
2014).
120
In plants, the biological role of Fe-S proteins in genotoxic stress and abiotic stress is gradually
121
emerging (Liang et al., 2014; Inigo et al., 2016). There are no reports thus far that directly show
122
Fe-S cluster genes’ involvement in plant biotic stress resistance/tolerance. Identification of genes
123
and pathways involved in plant disease resistance is important to develop disease-resistant crops
124
either by breeding or biotechnological approaches. One of the problems of using classical
125
resistance (R) genes in crops to confer disease resistance is their durability because R genes are
126
often effective against a specific pathogen strain. Nonhost resistance (NHR) on the other hand
127
can be used to confer broad and durable disease resistance (Mysore and Ryu, 2004; Gill et al.,
128
2015; Lee et al., 2016; Lee et al., 2017; Niks and Marcel, 2009). NHR is a type of resistance
129
shown by all plant species against most potential pathogens (nonhost pathogens) and results
130
usually in compromised or failure to establish virulence (Heath, 2000; Mysore and Ryu, 2004).
131
Several genes involved in NHR have also been used to confer resistance against economically
132
important diseases in crop plants (reviewed in Fonseca and Mysore, 2019).
133
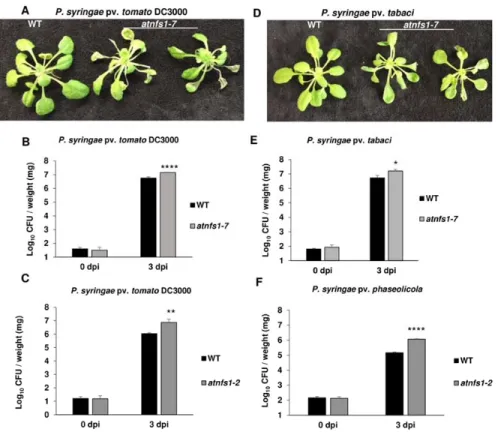
In this study, we demonstrate the role of Fe-S cluster genes in NHR and plant immunity. We
134
identified 11 Fe-S cluster genes to play a role in NHR by virus-induced gene silencing (VIGS) in
135
Nicotiana benthamiana. We further characterized the role of the mitochondrial AtNFS1 and its
136
interactor AtFH in NHR in Arabidopsis using mutants and overexpression lines that exhibited
137
enhanced susceptibility and increased disease resistance, respectively, against host and nonhost
138
pathogens. This is the first report showing the role of Fe-S cluster genes in combating plant
139
biotic stresses.
140
141
RESULTS
143
Virus-induced gene silencing of the NbNFS1 gene results in increased susceptibility to 144
nonhost bacterial pathogens in N. benthamiana 145
A forward genetics approach using the Tobacco rattle virus (TRV)-based VIGS system to
146
silence genes induced by a mixed cDNA elicitor library (del Pozo et al., 2004; Anand et al.,
147
2007; Senthil-Kumar and Mysore, 2014; Senthil-kumar et al., 2018) has been used to
148
successfully identify several genes involved not only in NHR but also in host resistance such as
149
gene-for-gene and pathogen-associated molecular patterns (PAMP)-mediated immunity (Rojas et
150
al., 2012; Senthil-Kumar et al., 2013; Kaundal et al., 2017; Nagaraj et al., 2016). One of the
151
cDNA clones identified in this screening, NbME26D10 (https://vigs.noble.org), displayed
152
homology to the AtNFS1 gene from Arabidopsis. AtNFS1 is annotated to encode a long
153
(AtNFS1-1) and a short (AtNFS1-2) isoform protein, which likely originated due to alternate
154
splicing. The AtNFS1 amino acid sequence is well conserved across plants, animals and even
155
prokaryotes as shown by multiple alignment (Supplemental Figure S1).
156
To further confirm that NFS1 plays a role in NHR, two VIGS constructs were generated with
157
distinct regions of the NbNFS1 coding sequence (NbNFS1-a; 522-822 bp and NbNFS1-b;
120-158
420 bp), for the downregulation of NbNFS1 in N. benthamiana (Supplemental Figure S2A).
159
The TRV2:GFP construct was used as a control (GFP has no sequence similarity to plant
160
genomic DNA and thus will not cause gene silencing) to rule out the possibility of TRV or
161
Agrobacterium tumefaciens infection alone causing the observed phenotype. NbNFS1 transcript
162
levels were significantly downregulated by VIGS (Supplemental Figure S2B).
163
TRV2:PHYTOENE DESATURASE (PDS) was used as a positive control for VIGS
164
(Supplemental Figure S3). Three weeks after silencing, fully expanded N. benthamiana leaves
165
above the TRV inoculation site were infiltrated with nonhost pathogen Pseudomonas syringae
166
pv. tomato T1 expressing GFPuv (encodes GFP that is optimized for maximal fluorescence 167
when excited by UV light; Wang et al., 2007) using a needleless syringe. We monitored
168
multiplication of P. syringae pv. tomato T1 under UV light. Multiplication of GFPuv-labeled
169
bacteria was more pronounced in NbNFS1-silenced plants compared to control plants two days
170
after pathogen inoculation (Fig. 1A). Bacterial multiplication was quantified at 0, 1, 2 and 3 days
171
after infection. NbNFS1-silenced plants were significantly more susceptible compared to the
unsilenced control plants upon P. syringae pv. tomato T1 infection at 1, 2 and 3 days after
173
pathogen infection (Fig. 1B). NbNFS1-silenced plants, when compared to the control, were also
174
significantly more susceptible to another nonhost pathogen, P. syringae pv. glycinea at 1, 2 and 3
175
days after pathogen infection (Supplemental Figure S3C). To investigate whether the NbNFS1
176
transcript levels were induced upon pathogen infection, we monitored transcript levels in
wild-177
type (WT) N. benthamiana plants at 0 h, 8 h, 1 d and 3 d after spray inoculation with the nonhost
bacterial pathogen P. syringae pv. tomato T1 and the host pathogen P. syringae pv. tabaci.
179
NbNFS1 expression was significantly induced after infection with both host and nonhost
180
pathogens at three days after infection (Fig. 1C, D). NbNFS1 transcripts were quantified in
181
unsilenced (TRV2:GFP) and NbNFS1-silenced plants after treatment with salicylic acid (SA), a
182
key phytohormone involved in plant defense. Interestingly, NbNFS1 transcripts were
183
significantly upregulated upon 1 mM SA treatment at 12 hours post infection in control plants
184
while in silenced plants, NbNFS1 transcripts remained unaltered. This indicates that NbNFS1
185
could be involved in the SA-mediated plant defense pathway (Fig. 1E). We also measured the
186
expression of the SA-mediated defense marker gene, NbPATHOGENESIS RELATED1 (PR1),
187
between control and silenced NbNFS1 plants and found it to be significantly reduced in
NbNFS1-188
silenced plants at basal levels without any pathogen treatment (Fig. 1F). Collectively, the above
189
results suggest a role for NbNFS1 in nonhost defense responses in plants and the SA signaling
190
pathway. Since NbNFS1-silenced plants showed disease susceptibility to bacterial nonhost
191
pathogens, we hypothesized that other Fe-S pathway genes may also play a role in NHR.
192
Silencing of Fe-S cluster genes in N. benthamiana compromises nonhost resistance 193
One of the Fe-S genes, NbNFS1, when silenced, resulted in enhanced susceptibility to nonhost
194
pathogens. Therefore, we sought to investigate if other Fe-S cluster pathway genes also play a
195
role in plant defense responses. To do this, we obtained gene sequences for all curated Fe-S
196
cluster genes available in Arabidopsis (Balk and Lobreaux, 2014; Balk and Pilon, 2011). We
197
identified N. benthamiana homologs using a cut-off e-value of 1e-10 for these genes using
198
BLASTX and N. benthamiana Genome v1.0.1 predicted protein database from the Sol Genomics
199
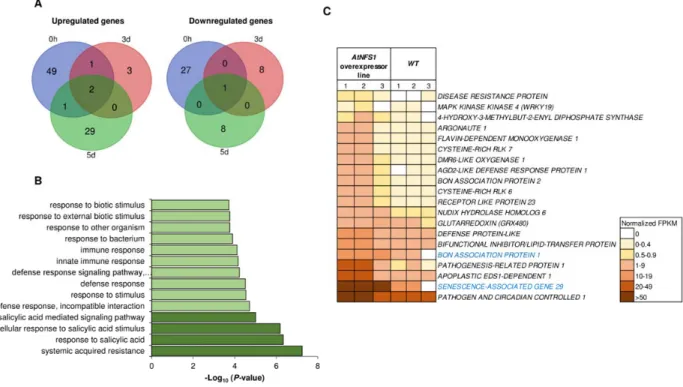
Network (SGN) website (https://solgenomics.net/) (Supplemental Table S1). We then used the
200
SGN VIGS Tool (Fernandez-Pozo et al., 2015) to select a fragment length of around 300 bp for
201
each N. benthamiana Fe-S cluster homolog and designed primers for Gateway cloning into the
202
TRV2-VIGS vector (Supplemental Table S1).
203
Twenty three Fe-S cluster genes were cloned into the TRV2-VIGS binary vector and
204
transformed into A. tumefaciens strain GV2260. VIGS was performed individually for each of
205
the 23 genes in N. benthamiana. Three-weeks later, silenced plants were inoculated with a
206
nonhost pathogen as described above for NbNFS1-silenced plants. Eleven out of 23 Fe-S cluster
207
genes, when silenced, compromised NHR (Table 1 and Supplemental Figure S4).
Interestingly, a gene that encodes a known interactor of NFS1, NbFH (Turowski et al., 2012),
209
when silenced, also compromised NHR against P. syringae pv. tomato T1 at 2 and 3 days after
210
pathogen infection (Fig. 2). Some of the other Fe-S cluster genes that, when silenced,
211
compromised NHR at 1, 2 and 3 days after pathogen infection include two mitochondrial
212
ferredoxin genes that display high sequence identity (87%) between them: MITOCHONDRIAL
213
FERREDOXIN 1 (MFDX1) and MITOCHONDRIAL FERREDOXIN 2 (MFDX2). A third
214
mitochondrial ferredoxin gene, PYRIDINE NUCLEOTIDE-DISULFIDE OXIDOREDUCTASE
215
(ADXR), also displayed compromised NHR when silenced. All three mitochondrial ferredoxins
216
are thought to be involved in electron transfer reactions (Takubo et al., 2003). Other Fe-S cluster
217
silenced genes that result in increased susceptibility to nonhost pathogens are
218
GLUTAREDOXIN16 (GRX16), which encodes a plastidial Fe-S cluster assembly machinery
219
enzyme with oxidoreductase activity that can potentially mediate the reduction of disulfide bonds
220
(Garg et al., 2010), and CYTOSOLIC IRON-SULFUR PROTEIN ASSEMBLY 1 (CIA1), which
221
encodes a cytosolic Fe-S cluster protein (Balk et al., 2005; Luo et al., 2012) (Supplemental
222
Table S1 and Supplemental Figure S4). These results suggest that Fe-S cluster proteins play a
223
role in plant defense especially for NHR.
224
The Fe-S cluster biosynthesis genes AtNFS1 and AtFH contribute to Arabidopsis resistance 225
against host and nonhost pathogens
226
To determine if the role of Fe-S cluster genes in NHR is conserved in other plant species, we
227
obtained Arabidopsis mutants for AtNFS1 and AtFH. We obtained an AtNFS1 heterozygous
mutant from the GABI-KAT collection with a T-DNA insertion located on exon 2 (GABI-KAT
229
211c08), referred to here as atnfs1-7, and a homozygous SALK line with an insertion located in
230
the 5’ untranslated region (UTR) (SALK_083681C), referred to as atnfs1-2 (Supplemental
231
Figure S5). Since complete loss of NFS1 leads to lethality or severe developmental defects with
232
no viable seed production, we could not obtain a homozygous atnfs1 complete knockout line
233
(Couturier et al., 2013; Balk and Shaeddler, 2014). The atnfs1-7 heterozygous mutant showed
234
reduced AtNFS1 expression (Supplemental Figure S5) and shorter primary root length when
235
compared to WT Col-0 plants (Supplemental Figure S6B). The homozygous atnfs1-2 mutant
236
had reduced AtNFS1 expression (Supplemental Figure S5). Both atnfs1-7 and atnfs1-2 mutants
were inoculated with a host pathogen P. syringae pv. tomato DC3000 and the nonhost pathogens
238
P. syringae pv. tabaci and P. syringae pv. phaseolicola by flood inoculation (Ishiga et al., 2011,
239
2017), and samples were collected at 0 h and 3 days after infection for bacterial quantification. In
240
agreement with our previous N. benthamiana NbNFS1-silenced plants (Fig. 1), both atnfs1
241
mutants were also susceptible to a nonhost pathogen 3 days after pathogen infection (Fig. 3E
242
and F). Both mutants were also susceptible to the host pathogen P. syringae pv. tomato DC3000
243
3 days after pathogen infection when compared to the WT plants (Fig. 3B, C). The observed
244
difference in Log10 colony-forming units (CFU) between the atnfs1-7 mutant and the WT (Fig. 245
3B, E), for example, is equivalent to a three-fold-change increase for the atnfs1-7 mutant. The
246
Arabidopsis atnfs1-7 mutant also showed disease-related symptoms such as necrosis and wilting
247
three days after infection compared to WT plants (Fig. 3A, D). Similar to NbNFS1 (Fig. 1), the
248
AtNFS1 transcript was also significantly induced by the nonhost pathogens and 1 mM SA in WT
249
Col-0 (Supplemental Figure S7).
250
The susceptibility of two atfh mutants to host and nonhost pathogens was also tested. The
251
homozygous mutant line atfh1 (SALK_054434C), with a T-DNA insertion located in the 3’UTR,
252
and atfh2 (SALK_021263C, 5’UTR) were identified and obtained (Supplemental Figure S8).
253
Similar to AtNFS1, complete loss of AtFH function in Arabidopsis leads to plant lethality (Busi
254
et al., 2006;Couturier et al., 2013; Balk and Shaeddler, 2014). Reverse transcription quantitative
255
PCR (RT-qPCR) analysis showed that expression of AtFH in the atfh1 and atfh2 mutant lines is
256
significantly reduced compared to the WT control (Supplemental Figure S7). We performed a
257
pathogen flood inoculation assay using both athf1 and atfh2 mutants and WT control plants upon
258
infection with the nonhost pathogen P. syringae pv. phaseolicola and the host pathogen P.
259
syringae pv. tomato DC3000. Similar to NbFH VIGS-silenced plants (Fig. 2), the Arabidopsis
260
atfh1 and atfh2 mutant lines were highly susceptible to nonhost pathogens 3 days after pathogen
261
infection when compared to WT Col-0 (Fig. 4A, B). In addition, both atfh1 and atfh2 mutants
262
also supported more host bacterial multiplication when compared to WT Col-0 at 3 days after
263
pathogen infection (Fig. 4C). Differences in Log10 CFU observed between the atfh mutants and 264
the WT is equivalent to a two-fold change increase for the atfh mutant. (Fig. 4D). Similar to the
265
atnfs1-7 mutant, atfh1 mutant seedlings (10 days old) also showed slight developmental defects,
266
such as shorter roots at the seedling stage, when compared to WT plants (Supplemental Figure
267
S6A). No significant difference in shoot biomass could be observed for the atfh1 and atnfs1-7
mutants and the WT in adult plants (23 days old, Supplemental Figure S9). These results
269
suggest that both AtNFS1 and AtFH play a role in NHR and basal resistance in Arabidopsis.
270
AtNFS1 and AtFH overexpression lines showed enhanced resistance to infection with the 271
host pathogen P. syringae pv. tomato DC3000 272
The recent annotation of the Arabidopsis genome (Araport11) shows three possible variant
273
coding sequences for AtNFS1; two long isoforms AtNFS1.1 and AtNFS1.3 with 1,362 base pairs
274
(bp) that show identical sequences to each other; and a short isoform AtNFS1.2 with 978 bp and
275
missing 83 amino acid residues from its N-terminal region compared to AtNFS1.1
276
(Supplemental Figure S10). AtNFS1.3 is possibly a redundant form and therefore was excluded
277
from this study. Both the long and short AtNFS1 isoforms were amplified from cDNA
278
(Supplemental Figure S10).
In order to further confirm AtNFS1’s role in disease resistance, Arabidopsis lines overexpressing
280
either the long AtNFS1.1 isoform or the short AtNFS1.2 isoform were generated. The expression
281
of both AtNFS1.1 and AtNFS1.2 was significantly upregulated by ~10 fold in their respective
282
overexpression lines (Supplemental Figure S11). Three- to four-week-old AtNFS1.1 (
NFS1.1-1-283
OX and NFS1.1-2-OX) and AtNFS1.2 (NFS1.2-7-OX and NFS1.2-18-OX) overexpression lines
284
and the WT were flood inoculated (Ishiga et al., 2011) with the host pathogen P. syringae pv.
285
tomato DC3000. Both AtNFS1.1 and AtNFS1.2 overexpression lines were less susceptible to host
286
pathogen infection 3 days after infection in relation to the WT control plants, indicating that both
287
the AtNFS1 short and long isoforms have a role in disease resistance (Fig. 5A, B). Additionally,
288
the overexpression lines also showed less disease symptoms, such as wilting and water soaked
289
lesion formation, compared to WT plants upon leaf infiltration using a needleless syringe with P.
290
syringae pv. tomato DC3000 (CFU = 8 x 106) 3 days after infection (Fig. 5C). We also tested the 291
growth of the host virulent pathogen P. syringae pv. maculicola ES4326 in the short isoform
292
overexpressor lines (NFS1.2-7-OX and NFS1.2-18-OX) and WT plants using the flood
293
inoculation method. As shown in Supplemental figure S12, the AtNFS1 overexpression lines
294
exhibited enhanced resistance to P. syringae pv. maculicola ES4326 3 days after pathogen
295
infection (Supplemental Figure S12) in a similar manner to the resistance observed against P.
296
syringae pv. tomato DC3000 shown above.
297
In addition to AtNFS1 overexpression lines, we also generated two Arabidopsis AtFH
298
overexpression lines (FH-16-OX and FH-8-OX). Upon flood inoculation of both FH-OX lines
299
with the host pathogen P. syringae pv. tomato DC3000, similar to AtNFS1-OX lines, both
AtFH-300
OX lines showed reduced disease symptoms and accumulated less bacteria when compared to
301
WT Col-0 3 days after pathogen infection (Fig. 5D). We did not observe any obvious phenotypic
302
developmental changes for the AtNFS1 and AtFH overexpression lines. These results indicate
303
that both isoforms of AtNFS1 and AtFH are involved in basal resistance against P. syringae pv.
304
tomato DC3000 in Arabidopsis.
305
AtNFS1 and AtFH overexpression or knockdown does not significantly impair shoot 306
respiration or biomass in adult Arabidopsis plants
307
Both AtNFS1 and AtFH localize in the mitochondria and full knockout lines have impaired
308
growth causing embryo lethality possibly by affecting plant respiration. Knockdown mutants
with reduced AtNFS1 or AtFH gene expression, however, are viable and generate healthy plants.
310
In order to test a possible effect of the AtNFS1 and AtFH mutations or overexpression on
311
mitochondrial respiration rate, dark respiration of the entire shoot (hereafter shoot respiration)
312
was chosen as the most relevant metric in order to exclude non-mitochondrial photorespiration
313
(Graham, 1980). We measured shoot biomass and CO2 flux in the WT, AtNFS1 or AtFH 314
knockdown lines, AtNFS1 or AtFH overexpression lines, and a E1-2-OXOGLUTARATE
315
DEHYDROGENASE knockout mutant (ogdh2-2, SALK_055824C) as a control (Supplemental
316
Figure S13), which has been previously shown to have a reduced respiration rate
(Condori-317
Apfata et al., 2019). No significant differences were observed in the shoot respiration rate or
318
biomass among all genotypes tested compared to WT plants, with the exception of the control
319
ogdh2-2 mutant (Fig. 6). This suggests that the disease phenotypes observed in the AtNFS1 or
320
AtFH knockdown and overexpression lines are not due to an indirect effect from changes in
321
respiration rates or plant growth.
AtNFS1 is ubiquitously expressed in several organs in Arabidopsis 323
To study the expression pattern of AtNFS1 in various Arabidopsis tissues, a 906 bp promoter
324
fragment (from +7 CDS to -991 bp) of AtNFS1 was amplified by PCR, and cloned into a binary
325
vector for generation of a β-glucuronidase (GUS) reporter construct, pNFS1:GUS. The
326
expression pattern of the GUS gene under the control of the AtNFS1 promoter was determined by
327
GUS histochemical staining of 2-week-old homozygous stable transgenic plants expressing
328
pNFS1:GUS. AtNFS1 is highly expressed in all plant tissues according to publicly available gene
329
expression data from Genevestigator (Figure 7C, D). Consistent with the gene expression data,
330
we observed GUS expression in most plant organs including leaf vasculature, leaf veins, the
331
hypocotyl, primary and lateral roots, emerging lateral roots, and at the root tips (Figure 7A). We
332
further confirmed whether AtNFS1 expression is induced by a host pathogen by infecting
333
pNFS1:GUS expressing plants with P. syringae pv. tomato DC3000. Surprisingly, we did not
334
observe any difference in GUS staining in the leaves but we did observe increased GUS staining
335
in the roots of plants drenched with the bacterial pathogen suspension (Figure 7B).
336
AtNFS1 localizes to mitochondria
Sequence prediction analysis of AtNFS1 indicated a mitochondrial localization (score of 0.83)
338
but also a possible chloroplastic localization, albeit with a low score (0.16) according to
339
Multiloc2 (Blum et al., 2009). AtNFS1 was previously shown to localize to mitochondria
340
although no mitochondrial marker was used (Frazzon et al., 2007). In order to confirm the
341
subcellular localization of AtNFS1, this time using a mitochondrial co-localization protein
342
marker, and to rule out a possible chloroplast localization, we fused the CDS of AtNFS1
343
(AtNFS1.1) with GFP and cloned this into an A. tumefaciens binary vector. AtNFS1:GFP was
344
transiently expressed in N. benthamiana along with a plant mitochondrial marker, mCherry
345
(Nelson et al., 2007). Two days after the co-infiltration, leaves were observed under a confocal
346
microscope and co-localization of AtNFS1:GFP and mCherry could be observed specifically in
347
the mitochondria but not in the chloroplast (Supplemental Figure S14).
348
Overexpression of AtNFS1 results in upregulation of several defense-related genes 349
To understand the mechanism of reduced susceptibility to host pathogens in AtNFS1
350
overexpression lines, we used the NFS1.2-18-OX line as a representative line to perform
RNA-351
seq upon P. syringae pv. tomato DC3000 infection. The aerial part of 5-week-old Arabidopsis
352
seedlings was collected from three plants (n = 3) each for WT and NFS1.2-18-OX line at
353
different time points (0, 3, and 5 dpi) upon flood-inoculation with P. syringae pv. tomato
DC3000 (CFU =1.6 x 105). Total RNA was isolated from these samples. Individual RNA-seq
355
libraries were prepared, and uniquely indexed for Illumina sequencing. Differentially expressed
356
genes (DEGs) were identified using the criteria, Log2 fold change > 1 and < -1; FDR < 0.05, for 357
the NFS1.2-18-OX line and the WT upon P. syringae pv. tomato DC3000 infection. Based on
358
these criteria, a total of 129 DEGs were identified for all time points between the NFS1.2-18-OX
359
line and the WT (Fig. 8A; Supplemental Table S2). A total of 85 upregulated DEGs for all time
360
points were identified in the NFS1.2-18-OX line compared to the WT (Fig. 8A; Supplemental
361
Table S3). Subsequently, these 85 upregulated DEGs were functionally classified according to
362
gene ontology (GO) terms using AgriGO2 (Tian et al., 2017) (Supplemental Table S4). To
363
identify the overrepresented functional categories, a singular enrichment analysis (SEA) was
364
performed. SEA compares each annotated gene to all annotated expressed genes. In total, 46 GO
365
terms displayed overrepresentation for biological processes (FDR < 0.001) (Supplemental
Figure S15A; Supplemental Table S4). Interestingly, 12 overrepresented GO terms belong to
367
defense-related processes such as immune response, innate immune response, response to biotic
368
stimulus, systemic acquired resistance and SA-mediated signaling pathway. We also found 29
369
GO terms overrepresented (FDR < 0.05) for the same dataset of 85 upregulated DEGs using
370
Panther (Mi et al., 2019) (Fig. 8B; Supplemental Table S5). Similar to AgriGO2, the same
371
defense-related GO pathways were found to be enriched using Panther such as response to SA,
372
systemic acquired resistance and immune responses (Fig. 8B). In addition to defense-related
373
processes, we also observed overrepresentation of GO terms related to Fe-S clusters such as
374
metal ion binding, cofactor binding, and adenyl nucleotide binding (Supplemental Table S4;
375
Supplemental Figure S15B). Additionally, using AgriGO2 and Panther, we also identified
376
overrepresented GO terms from other biological processes that may also play a role in plant
377
defense such as response to oxygen and response to stress (Supplemental Tables S4 and S5).
378
Strikingly, a total of 53 upregulated DEGs were identified at the 0 h basal control (fold change
379
>1; FDR < 0.05) in the NFS1.2-18-OX line compared to the WT (Supplemental Table S6).
380
Interestingly, most of the defense-related DEGs upregulated at all time points in the
NFS1.2-18-381
OX line compared to the WT belonged to the 0 h time point (18 out of 20; Fig. 8C).
382
Additionally, a total of 28 downregulated DEGs were identified at the 0 h time point between the
383
NFS1.2-18-OX line and the WT (Fig. 8A; Supplemental Table S7) but no molecular functions
384
related to defense responses were found, and other GO terms like hydrolase activity and binding
385
were identified from these DEGs.
386
We validated the differential expression of seven upregulated defense-related genes (0 h time
387
point) from our RNA-seq dataset by RT-qPCR (Supplemental Figure S16) and confirmed their
388
increased expression in the NFS1.2-18-OX line compared to the WT in the absence of pathogen
389
infection. Overall, these results suggest that the overexpression of AtNFS1 constitutively
390
upregulates several defense-related genes. Even though several defense-response related genes
391
were constitutively upregulated in the NFS1 overexpressor line compared to the WT, we did not
392
see any developmental defects like stunting or yield penalty phenotypes associated with it.
393
394
DISCUSSION
396
Fe-S proteins carry out the synthesis and assembly of Fe-S clusters mainly in plastids,
397
mitochondria, and in the cytosol of plants. Fe-S clusters are protein cofactors involved in
398
catalysis, electron transport and sensing of ambient conditions required for several plant
399
physiological processes such as amino acid metabolism, DNA replication and development
400
(Couturier et al., 2012). In this study, we show a new role for the Fe-S cluster pathway in plant
401
innate immunity. Using a VIGS-mediated forward genetics screening in N. benthamiana, we
402
identified the CYSTEINE DESULFURASE coding gene, NbNFS1, which, when silenced,
403
compromised NHR against the bacterial pathogens tested. Expression of NbNFS1 was induced
404
by the host pathogen, nonhost pathogens and SA. NbPR1 expression was decreased at 0 h after
405
infection in the NbNFS1-silenced plants compared to the unsilenced control. Both atnfs1-7 and
406
atnfs1-2 mutants from Arabidopsis also showed increased susceptibility to host and nonhost
407
pathogen infection compared to WT plants. Arabidopsis AtNFS1 overexpressors exhibited
408
resistance against the host pathogen P. syringae pv. tomato DC3000. AtNFS1 transcript was
409
significantly induced upon nonhost pathogen infection and SA treatment in WT plants similarly
410
to N. benthamianaNbNFS1. This result is consistent with an earlier report showing induction of
411
soybean (Glycine max) GmNFS1 expression in roots upon treatment with SA (Heis et al., 2011).
412
Furthermore, we showed that the nuclear-encoded AtNFS1:GFP co-localized in mitochondria
413
along with the mitochondria tracker mCherry. This rules out a possible plastidial localization for
414
AtNFS1 and further confirms previous studies (Frazzon et al., 2007).
415
In addition to NbFS1, we show that 11 out of 23 other Fe-S cluster genes tested also play a role
416
in NHR in N. benthamiana. Along with NFS1, we also further characterized its interactor, FH
417
(Frazon et al., 2007; Turowski et al., 2015), for its role in basal and NHR in N. benthamiana and
418
Arabidopsis. Amongst the 11 Fe-S cluster genes found to be involved in NHR, here we highlight
419
a few for their possible roles in plant defense. Mitochondrial ferredoxins 1 and 2 (NbMFDX1 and
420
NbMFDX2) and a ferredoxin reductase (NbMFDXR) are involved in a variety of redox reactions
421
in the cell, electron transfer, and reducing cytochrome C in the presence of nicotinamide adenine
422
dinucleotide phosphate (NADPH) (Takubo et al., 2003). These cellular processes have been
423
shown to be important for plant defense responses such as hypersensitive response (Dayakar et
424
al., 2003). We also identified a member of the glutaredoxin family, GLUTARREDOXIN16
(NbGRX16), which resulted in increased susceptibility to nonhost pathogens when silenced.
426
Glutaredoxins are known to mediate redox regulation of proteins because of their capacity to
427
catalyze disulfide bonds. GLUTAREDOXIN480 (GRX480 encoded by At1g28480) has been
428
shown to interact with TGACG SEQUENCE-SPECIFIC BINDING PROTEIN (TGA)
429
transcription factors in a yeast two-hybrid screening, using TGA as bait (Ndamukong et al.,
430
2007). TGA transcription factors are well known regulators of PR genes since they are known to
431
bind the defense nonexpresser of PR gene1 (NPR1) protein (Kesarwani et al., 2007). Further, it
432
was shown that GRX480 transcripts were significantly induced by SA (Ndamukong et al., 2007).
433
We also investigated the role of the AtNFS1 interactor protein, AtFH, in disease resistance.
434
Reduced expression of FH in N. benthamiana and Arabidopsis made the plants more susceptible
435
to both host and nonhost pathogens. Overexpression of AtFH in Arabidopsis reduced bacterial
436
accumulation of the host pathogen P. syringae pv. tomato DC3000 when compared to WT Col-0.
437
FHs in general are known to bind iron molecules and are involved in iron homeostasis,
438
respiration, Fe-S cluster biogenesis and redox control (Turowski et al., 2012).
439
Transcriptome analysis of the AtNFS1 overexpressor line compared to the WT identified several
440
upregulated DEGs involved in plant defense responses in the AtNFS1 overexpressor line
441
compared to the WT control. Several GO terms belonging to plant immunity were found to be
442
significantly overrepresented in the upregulated gene list (Supplemental Table S8). For
443
example, the pathogenesis-related gene PR1, PATHOGEN AND CIRCADIAN CONTROLLED 1
444
(PCC1) that is upregulated in response to both virulent and avirulent strains of P. syringae pv.
445
tomato (Mir et al., 2013) and the systemic acquired resistance related MONOOXYGENASE 1
446
(FMO1) were upregulated in the AtNFS1 overexpressor line compared to the WT (Fig. 8C;
447
Supplemental Table S3). Amongst the top five upregulated DEGs with the highest fold change,
448
we found the genes encoding cysteine-rich receptor-like kinases (CRKs) 6 and 7, which are
449
involved in pathogen-induced activation of defense responses against host pathogen infection
450
(Yeh et al., 2015). Most defense-related upregulated DEGs identified here belong to the 0 h time
451
point, suggesting that AtNFS1 has a role in basal disease resistance. We did not observe any
452
significant differences in terms of developmental phenotype or size between the AtNFS1
453
overexpressor lines and WT plants. This is probably because the upregulation of defense genes
454
was moderate (< 5 fold) in contrast to the constitutive expresser of PR genes (cpr) mutants, for
455
example, which have constitutive expression of PR genes causing severe stunting (Clarke et al.,
1998; Bowling et al., 1997). This moderate upregulation of defense-related genes may suggest
457
that NFS1 overexpression would be a good potential strategy for generating disease-resistant
458
crops.
459
In order to address a possible pleiotropic effect in AtNFS1 or AtFH overexpressor or mutant lines
460
resulting in disease resistant or susceptible phenotypes, we tested whether knockdown mutants
461
and overexpressors of AtFH and AtNFS1 differ in shoot respiration or growth. We found no
462
significant differences in shoot respiration or fresh shoot biomass in adult plants among all the
463
lines tested. This suggests that the observed disease phenotypes in AtNFS1 or AtFH
464
overexpressor or mutant lines are not an indirect effect due to changes in respiration rates due to
465
mitochondrial defects, but likely stem from immune regulation changes. The lack of difference
466
observed in respiration rates and fresh biomass for adult plants between all genotypes are in
467
agreement with previous shoot biomass data collected for the AtNFS1 overexpressor or
468
knockdown lines in Arabidopsis (Armas et al., 2019).
469
In agreement with the upregulation of NFS1 transcripts both in Arabidopsis and N. benthamiana
470
after SA treatment, we also found several genes involved in SA signaling to be upregulated in
471
the AtNFS1 overexpression line compared to the WT (Supplemental Tables S3 and S7;
472
Supplemental Figure S15A). The gene AGD2-LIKE DEFENSE RESPONSE PROTEIN 1
473
(ALD1) was induced by 5.5 fold at the 0 h time point compared to the WT in the AtNFS1
474
overexpressor line. ALD1 has been shown to be required for the accumulation of SA besides
475
affecting basal plant defense responses (Cecchini et al., 2015). The above mentioned PCC1 (Mir
476
et al., 2013)and the chloroplastic Fe-S cluster binding ferredoxin
4-HYDROXY-3-METHYLBUT-477
2-ENYL DIPHOSPHATE SYNTHASE (HDS) are SA-responsive genes and the later has been
478
shown to modulate SA-mediated disease resistance (Gil et al., 2005) (Supplemental Tables S3
479
and S8). These data suggest that AtNFS1 may play a role in SA-mediated plant defense
480
signaling.
481
In conclusion, the genetic and transcriptomic data, in combination with disease phenotype assays
482
presented here suggest that the Fe-S cluster interactor proteins NFS1 and FH are involved not
483
only in NHR but also in basal resistance. We propose a new role for Fe-S cluster genes in plant
484
immunity responses that has been overlooked until now. Our results also suggest that NFS1, its
interactor FH, and possibly other Fe-S cluster genes could be new targets for crop breeding and
486
genetic engineering to generate durable disease-resistant varieties.
487
488
489
MATERIALS AND METHODS
490
Plant materials
491
Arabidopsis thaliana plants were grown on ½ MS media in a controlled chamber maintained at a
492
22 °C constant temperature and 50% relative humidity with a 12 h / 12 h day / night cycle. For
493
experiments involving soil-grown plants, agar germinated plants were transferred to pots
494
containing Metro-Mix 360 two weeks after germination and maintained at the same conditions as
495
described above in a growth chamber. Nicotiana benthamiana plants were grown in pots with
496
Metro-Mix 360 in the greenhouse under a 16 h / 8 h day / night photoperiod. Mutants
497
characterized in this study are as follows: SALK_054434C (atfh1), SALK_021263C (atfh2),
498
Gabi-Kat_211c08 (atnfs1-7), SALK_083681C (atnfs1-2) and SALK_055824C (ogdh2-2). Seeds
499
were obtained from the ABRC stock (https://abrc.osu.edu/) and belong to the SALK T-DNA
500
collection (http://signal.salk.edu/cgi-bin/tdnaexpress; Alonso et al., 2003). The Arabidopsis
501
atnfs1-7 mutant was obtained from the Gabi-Kat collection (https://www.Gabi-Kat .de/). All
502
SALK and Gabi-Kat lines were grown in ½ MS media plates containing kanamycin (50 µg/mL)
503
and sulfadiazine (8 µg/mL), respectively, and genotyped using PCR to confirm homozygosity
504
(Supplemental Table S9).
505
Cloning and Arabidopsis transformation
506
The coding sequence (CDS) of AtNFS1 and AtFH were amplified by PCR from Arabidopsis
507
cDNA (1 µg) with Phusion High-Fidelity DNA Polymerase (New England Biolabs, Ipswich,
508
MA), purified using QIAquick PCR Purification Kit (Qiagen, Valencia, CA) to remove any
509
primer dimers and incubated with Invitrogen BP clonase to generate an entry vector
510
(pDONR207) by transformation of TOP10 (Invitrogen, Carlsbad, CA) competent cells. The
511
product from BP reaction was introduced into the following binary vectors containing the
512
Cauliflower mosaic virus promoter (CaMV 35S) that drives the gene construct: pMDC32 for
overexpression lines (NFS1-OX) and pMDC83 (AtNFS1:GFP) via LR reaction (LR clonase,
514
Invitrogen) and then transformed into Agrobacterium tumefaciens strain GV3101 for stable
515
transformation in Arabidopsis using the floral dip method (Clough and Bent, 1998). All cloned
516
genes were checked by sequencing using Sanger sequencing. The 907 bp promoter region of
517
AtNFS1 was amplified by PCR from genomic DNA (50 ng) and cloned into the GUS reporter
518
binary vector pMDC162 by GATEWAY cloning and stably-transformed into Arabidopsis in the
519
same way as described above for GUS histochemical studies. All primers used in this work can
520
be found in Supplemental Table S1.
521
Virus-induced gene silencing (VIGS)
522
Fragments from Fe-S cluster genes (~300 bp) were amplified by PCR from N. benthamiana
523
cDNA and cloned (GATEWAY) into entry vector pMDC207 followed by LR cloning into the
524
VIGS TRV2 vector in the same way as described in the above section. Plasmids were
525
transformed into A. tumefaciens strain GV2260 for VIGS. A normalized library obtained from a
526
mixed elicitors-treated N. benthamiana leaves (Anand et al., 2007; Senthil-kumar et al., 2014)
527
was used to screen genes involved in NHR.
528
VIGS of Fe-S cluster genes was performed in N. benthamiana as described previously
(Senthil-529
Kumar and Mysore, 2014; Chung et al., 2004). Briefly, A. tumefaciens strain GV2260 carrying
530
TRV2 with the gene of interest (GOI) to be silenced (TRV2:GOI), TRV2:GFP, TRV2:PDS
531
(Phytoene Desaturase) were grown overnight in LB media with appropriate antibiotics. Cultures
532
were harvested by centrifugation and re-suspended in infiltration buffer (10 mM MES, 150 µM
533
acetosyringone, pH 5.6). Samples were incubated for three hours, and re-suspended in a 10 mM
534
MES buffer before infiltration. Suspensions of TRV2:GOI and TRV1 were mixed in a 1:1 ratio
535
to a final optical density (OD600nm) of 0.4 before infiltration into three week-old N. benthamiana 536
plants and allowed another three weeks for silencing to occur. TRV2:PDS was used as an overall
537
positive control for every VIGS-silencing experiment and TRV2:GFP was used as the
538
unsilenced control.
539
Pathogen infection assays
540
We used the flood inoculation method that mimics natural infection for our disease assays in
541
Arabidopsis (Ishiga et al., 2011; Uppalapati et al., 2008; Ishiga et al., 2017). In brief,
old plants grown in ½ MS media were incubated for 5 minutes with 40 mL of a bacterial
543
suspension with 0.015% (v/v) Silwet L-77, containing either one of the following strains:
544
Pseudomonas syringae pv. tomato DC3000 (host), P. syringae pv. tabaci (nonhost) and P.
545
syringae pv. phaseolicola (nonhost) at the concentrations of 8x104, 1.6x106, and 8x105 colony
546
forming units (CFU), respectively. Disease symptoms were observed at three or five days after
547
infection. We collected infected tissues at 0 h, 1 day, and 3 days after the infection and weighed
548
each plant for normalization of CFU. Four plants per time point (n = 4) per genotype were used.
549
Whole seedlings were placed in tubes containing grinding beads and 200 µL of sterile water and
550
ground for 1 min to obtain a homogeneous suspension. Suspensions were 10-fold serially diluted
551
for plating. Aliquots of 10 µL were then plated on KB medium with the appropriate selection
552
antibiotic and incubated at 28 °C for two days after which CFU were counted (Fonseca et al.
553
2015; Durrant et al., 2007). For disease symptom visualization, we used a CFU of 8x106 and
554
infiltrated fully grown leaves of 4 week-old soil-grown Arabidopsis plants using a needleless
555
syringe. Disease symptoms were observed and recorded 24 h to 48 h after infiltration.
556
For pathogen infection assays in N. benthamiana, VIGS-silenced and control plants were
557
infiltrated with a needleless syringe at the abaxial part of the leaf with a bacterial suspension
558
containing the following nonhost pathogens: P. syringae pv. tomato T1 and P. syringae pv.
559
glycinea at 8x105 CFU. We collected tissues at 0, 1, 2 and 3 days after infection. Two leaf-disks
560
of 10 mm diameter were collected per plant from at least 5 plants (n = 5) using a 0.5 cm2 borer.
561
Leaf discs were placed in tubes containing grinding beads and 500 µL of sterile distilled water
562
and ground to obtain a homogeneous suspension. Suspensions of 20 µL were transferred to a
563
plate containing 180 µL of sterile distilled water and 10× serial dilutions were made. Aliquots of
564
10 µL were then plated on KB medium with the appropriately selected antibiotic and incubated
565
at 30°C for two days, after which CFU were counted. We monitored bacterial growth using
566
GFPuv-labeled P. syringae strains two to three days after infection in N. benthamiana. Statistical
567
analyses were done using Student’s t test of the differences between two means of
log-568
transformed data or by one-way ANOVA using the GraphPad Prism software.
569
Subcellular localization
570
To confirm mitochondrial localization of AtNFS1, we co-infiltrated N. benthamiana leaves with
571
A. tumefaciens strain GV2260 carrying a binary vector pMDC83 that has the CaMV 35S
promoter driving AtNFS1-GFP and another A. tumefaciens GV2260 strain with the red
573
fluorescent organelle marker protein mCHERRY fused to a mitochondrial targeting sequence
574
from yeast (Saccharomyces cerevisiae), cytochrome oxidase IV (Nelson et al., 2007). Briefly, A.
575
tumefaciens strains carrying the above mentioned constructs were grown overnight and
576
centrifuged at 13,000 rpm to pellet bacteria. The pellet was re-suspended and incubated in
577
induction buffer (10 mM MES, 150 µM acetosyringone, pH 5.6) for 4 hours with gentle shaking.
578
Samples were centrifuged and the pellet was re-suspended in 10 mM MES buffer to a final
579
OD600nm = 0.15 each. Both A. tumefaciens strains were mixed at a 1:1 ratio and used to infiltrate 580
leaves of four-week-old greenhouse-grown N. benthamiana plants using a needleless syringe.
581
Two days after infiltration, the infiltrated area of the leaves were observed with an inverted Leica
582
TCS-SP8-X point scanning confocal laser scanning microscope (Leica Microsystems GmbH,
583
Mannheim, Germany) equipped with a 40x water-immersion objective. GFP was excited with
584
the 488 nm line of the white light laser and emission was detected at 493-551 nm. mCherry was
585
excited with the 587 nm line of the Argon laser and emission was detected at 604-661 nm.
586
Histochemical GUS staining
587
GUS histochemical staining of Arabidopsis transgenic lines expressing pNFS1:GUS was done
588
using the chromogenic substrate X-gluc according to Hull and Devic (1995). Whole Arabidopsis
589
plants or specific tissue were submerged in a 2 mM X-gluc solution in 100 mM sodium
590
phosphate, pH 7.0, 10 mM EDTA, 1 mM K3Fe(CN)6 and 0.1% (v/v) Triton X-100, applied
591
vacuum for 2 min, and incubated at 37 °C overnight. The following day, the tissues were cleared
592
by successive washes in 70% (v/v) ethanol to remove chlorophyll for visualization. Images of
593
GUS staining were taken by the Olympus SZX12 Stereo Microscope (Olympus Corporation,
594
Tokyo, Japan) using the transmitted light.
595
596
RNA extraction and reverse transcription quantitative PCR (RT-qPCR)
597
Total RNA was isolated from 50 mg of leaves using RNeasy Mini Kit (Qiagen), followed by
598
TURBO DNase treatment (Invitrogen). At least 1 µg of RNA was used for cDNA synthesis
599
using SuperScript III reverse transcriptase (Invitrogen) according to the manufacturer's
600
instructions. Amplicons were amplified from cDNA using gene specific primers (Supplemental
601
Table S1), and their relative gene expression values were normalized to the AtACTIN2 (ACT2)
gene for Arabidopsis samples and the NbACTIN1 gene for N. benthamiana using the
603
comparative CT method (Livak and Schmittgen, 2001). qPCR was performed in a Bio-Rad
604
instrument (Eppendorf, Hauppauge, NY) using SYBR Green (Applied Biosystems, Foster City,
605
CA) to monitor dsDNA synthesis. The program used to perform reactions was as follows: 95 °C
606
for 5 mins, followed by 40 cycles of extension at 72 °C for 20 seconds, annealing at 60 °C for 1
607
min. To assess amplicon quality the melting curve was generated at 65 °C until 95 °C with 0.5
608
°C increments for 5 seconds.
609
RNA-seq experiment
610
Total RNA from Arabidopsis was isolated from 60 mg of leaf tissue using Spectrum Plant Total
611
RNA Kit (Sigma-Aldrich), and residual DNA was removed from each RNA sample by DNase
612
treatment using TURBO DNA-free Kit (Thermo Fisher Scientific, Waltham, MA ) and then
613
purified by RNeasy MinElute Cleanup Kit (Qiagen). The quality and profile of the RNA samples
614
were checked on an Agilent 2100 Bioanalyzer (Agilent Technologies) using an Agilent RNA
615
6000 Nano Kit (Agilent Technologies). The RNA samples were quantified on Qubit 2.0
616
Fluorometer using Qubit RNA BR Assay Kit (Invitrogen). RNA-seq libraries were prepared
617
using TruSeq Stranded mRNA Sample Prep Kits (Illumina). Briefly, mRNA was purified from 1
618
µg of total RNA, fragmented and converted to double-stranded DNA for sequencing. Individual
619
libraries were uniquely indexed using TruSeq RNA Single Indexes (Illumina) and pooled in an
620
equimolar ratio. The pooled libraries were sequenced on a Hiseq4000 sequencing machine
621
(Illumina).
622
The sequencing quality of the reads generated were examined by FastQC software (v0.10.1)
623
(http://www.bioinformatics.babraham.ac.uk/projects/fastqc/) and only high quality samples were
624
used. The filtered clean reads were mapped to the Arabidopsis genome (Araport11, release 2016
625
downloaded from https://www.araport.org/) to estimate raw counts and effective read lengths by
626
Salmon tool v0.7.2 (Patro et al., 2017). All samples displayed a high mapping rate to the
627
Arabidopsis genome (> 95%). Raw counts from Salmon were normalized across all samples by
628
median normalization, and differential expression was estimated using the DESeq2 module from
629
the Bioconductor R package (Love et al., 2014) and an in-house Perl script. Gene expression was
630
quantified as FPKM (fragments per kilobase of exon per million fragments mapped) values, and
631
identified differentially expressed genes (DEGs) were required to have a false-discovery rate
(FDR) < 0.05. DESeq2 software (Love et al., 2014) was further used to perform differential
633
expression analysis, which included dynamic filtering of low expressed genes to prevent
634
spurious identification of DEGs. Hierarchical tree graphs for biological process and molecular
635
function using overrepresented GO terms from all DEGs were obtained by singular enrichment
636
analyses (SEA), using AgriGO2 (Tian et al., 2017). GO terms were declared enriched when FDR
637
< 0.001. Overrepresented GO terms using the same DEGs dataset was confirmed using Panther
638
default FDR < 0.05 (Mi et al., 2019).
639
Shoot respiration
640
Arabidopsis WT (Col-0), ogdh2-2, atfh1, FH-16-OX, atnfs1-7, NFS1.2-18-OX and NFS1.1-1-OX
641
seeds were germinated and seedlings were grown in solid ½ MS-agar media. After five days of
642
growth, evenly germinated seedlings were transferred to a Falcon 6-well culture plate (Corning,
643
Arizona, USA) with each well containing liquid ½ MS media. Nine plants were pooled together
644
per well to make a biological replicate with a total of 5 biological replicates per genotype. After
645
another 18 days of growth on liquid media, the plants were transferred to darkness 1 hour before
646
CO2 flux measurements by wrapping an entire plate in aluminum foil. Shoots were severed from 647
the roots, then the fresh weight of the shoots was recorded before transferring the shoots into a
648
light- and air-tight chamber having a volume of 7.5 cm3. The respiration measurements were
649
performed using a LI-850 CO2/H20 gas analyzer (LI-COR Biosciences, NE, USA) in a dark
650
room with a supplemental photosynthetically inactive yellow LED light. The chamber was
651
connected to the LI-850 instrument, buried in a bead bath with the temperature set at room
652
temperature (23 °C), and the CO2 flux was measured with an observation duration of 2 minutes 653
30 seconds using LI-8x0 v1.02 software. Specific shoot respiration rate (nmol CO2 gfw-1 s-1) was 654
calculated from the raw CO2 flux data using R version 3.6.0 where a linear regression was used 655
to calculate the CO2 flux rate with dead band set at 60 seconds, and then divided by total shoot 656
fresh weight.
657
658
Accession Numbers
659
Sequence data from this article can be found in the Arabidopsis Genome Initiative or
660
EMBL/GenBank databases under the following accession numbers: AtNFS1 (AT5G65720),