In Vivo
Characterization of the Nonessential Budding Yeast Anaphase-Promoting
Complex/Cyclosome Components Swm1p, Mnd2p and Apc9p
Andrew M. Page*
,†Vicky Aneliunas,
†John R. Lamb
‡,1and Philip Hieter
†,2*Program in Biochemistry, Cellular, and Molecular Biology, The Johns Hopkins University School of Medicine, Baltimore, Maryland 21205, ‡Department of Molecular Biology and Genetics, The Johns Hopkins University School of Medicine, Baltimore, Maryland 21205 and
†Michael Smith Laboratories, University of British Columbia, Vancouver, British Columbia V6T 1Z4, Canada
Manuscript received December 17, 2004 Accepted for publication April 6, 2005
ABSTRACT
We have examined thein vivorequirement of two recently identified nonessential components of the budding yeast anaphase-promoting complex, Swm1p and Mnd2p, as well as that of the previously identified subunit Apc9p. swm1⌬ mutants exhibit synthetic lethality or conditional synthetic lethality with other APC/C subunits and regulators, whereasmnd2⌬mutants are less sensitive to perturbation of the APC/C. swm1⌬mutants, but not mnd2⌬mutants, exhibit defects in APC/C substrate turnover, both during the mitotic cell cycle and in␣-factor-arrested cells. In contrast,apc9⌬mutants exhibit only minor defects in substrate degradation in␣-factor-arrested cells. In cycling cells, degradation of Clb2p, but not Pds1p or Clb5p, is delayed inapc9⌬. Our findings suggest that Swm1p is required for full catalytic activity of the APC/C, whereas the requirement of Mnd2p for APC/C function appears to be negligible under standard laboratory conditions. Furthermore, the role of Apc9p in APC/C-dependent ubiquitination may be limited to the proteolysis of a select number of substrates.
U
BIQUITIN-mediated proteolysis (Ciechanover In addition to APC11 and APC2, the APC/Ccom-prises 11 other subunits (PageandHieter1999;Peters 1994;Hochstrasser 1996) of cell cycle
regula-tors is required for cell cycle transitions. In eukaryotes, 1999; Zachariae andNasmyth 1999). Structural ho-mologs of these subunits are absent from other SCF-like the polyubiquitination and subsequent degradation of
Pds1p/securin and mitotic cyclins is essential for the complexes and their functions are poorly understood. Homologs of APC1, APC4, APC5, CDC26, and DOC1 metaphase-to-anaphase transition and exit from mitosis.
are found throughout the eukaryotic kingdom, as are The anaphase-promoting complex/cyclosome (APC/C),
the three tetratricopeptide repeat (TPR) proteins CDC16, a large and complex member of the RING-finger family
CDC23, and CDC27. A fourth TPR protein, APC7, appears of E3 ubiquitin ligases, catalyzes the polyubiquitination
to be present only in multicellular eukaryotes, whereas of these and other substrates. RING-finger proteins have
Apc9p has been identified only in several fungi. In bud-been shown to promote substratein vitroubiquitination
ding yeast, APC/C function is essential for viability in in conjunction with E2s and a ubiquitin-activating system
otherwise normal cells, although the APC/C is dispens-and are believed to recruit ubiquitin-conjugating enzymes
able for cell survival in cells lacking both Pds1p and Clb/ to their cognate E3 complexes (Jacksonet al.2000). In
CDK activity. Deletion of most core APC/C subunits is
Skp1/cullin-homology/F-box (SCF) and VHL/Elongin
therefore lethal, but APC/C subunits Doc1p, Cdc26p, BC complexes, evidence suggests that the RING-H2
pro-and Apc9p are encoded by nonessential genes. tein ROC1/Rbx1p recruits E2s to cullins (Kamuraet
Regulation of mitotic APC/C activity is controlled by
al.1999;Skowyraet al.1999). In the case of the APC/C,
two specificity factors, Cdc20p and Cdh1p, both of APC2 contains a cullin-homology domain on its carboxy
which contain WD-40 repeats (Schwabet al.1997; Vis-terminus (Yu et al. 1998;Zachariae et al. 1998) and,
intinet al.1997;Fanget al.1998). Cdc20p and Cdh1p like other members of the cullin family, is thought to
may bind directly to APC/C substrates and recruit them act as a scaffold that promotes substrate ubiquitination
to the core complex, although the biochemical details by bringing E2/RING-H2 complexes into close
proxim-of how substrates are recognized by the APC/C are ity with substrate/specificity factor complexes (Yuet al.
poorly understood (BurtonandSolomon2001; Hili-1998;Gmachlet al. 2000;Leversonet al.2000;Tang
otiet al.2001;Pflegeret al.2001;Schwabet al.2001).
et al.2001).
At the metaphase-to-anaphase transition, APCCdc20p
cata-lyzes the ubiquitination of Pds1p, releasing Esp1p/sep-arase, which then cleaves Scc1p/cohesin to initiate sis-1Present address:Rosetta Inpharmatics, Seattle, WA 98109.
ter-chromatid separation and the onset of anaphase 2Corresponding author:Michael Smith Laboratories, 2185 East Mall,
Vancouver, BC V6T 124, Canada. E-mail: [email protected] (ZachariaeandNasmyth1999;Amon2001;
flanked byⵑ500 bp upstream and 250 bp downstream into
Fix2001;Nasmythet al.2001). APCCdc20palso
ubiquiti-theClaI-NotI sites of pRS426. See Table 1 for a list of strains
nates Clb5p and a fraction of Clb2p (Shirayamaet al.
and plasmids used in this article.
1999;Yeonget al.2000;Irniger2002) and the resulting Galactose shutoff assays:Cells were grown to midlog phase decrease in B-type cyclin activity, coupled with release at the permissive temperature (25⬚or 30⬚) in SC media lacking
uracil, tryptophan, or leucine, with 2% raffinose,
supple-of the phosphatase Cdc14p from the nucleolus, activates
mented with an additional 1⫻adenine, uracil, and/or
trypto-Cdh1p. As cells exit from mitosis, APCCdh1pubiquitinates
phan as appropriate. After washing cells twice with ddH2O,
the remaining B-type cyclins and other components of
cells were resuspended in the same media and arrested in
the mitotic apparatus such as Ase1p, Kip1p, Cin8p, and ␣-factor forⵑ3.5 hr or until⬎90% of cells had mating projec-Cdc5p (Juanget al.1997;Charleset al.1998;Shira- tions. Following␣-factor arrest, cultures were standardized to
similar densities. For experiments conducted at temperatures
yama et al. 1998; Gordon and Roof 2001;
Hilde-higher than the arrest temperature, cultures were shifted to
brandtandHoyt2001), as well as Cdc20p itself (Prinz
the nonpermissive temperature for 15 min before being
in-et al.1998;Gohet al.2000).
duced with 2% galactose for 30 min. After galactose induction,
Recently several groups have identified Swm1p and glucose was added to 2% and cycloheximide to 1 mg/ml to Mnd2p as new nonessential budding yeast APC/C com- repress transcription and translation from the GAL1
pro-moter. For each time point, volume-adjusted aliquots were
ponents by immunoprecipitation and mass
spectrome-removed from the culture, immersed in 1⫻ice-cold
SC-Ura-try of the purified complex (Yoonet al.2002;Hall et
Trp with no carbon source and rapidly centrifuged. Cell pellets
al.2003;Schwickartet al.2004). Structural homologs
were washed with 1 ml ice-cold ddH2O and transferred to
of both proteins were also identified in fission yeast Eppendorf tubes. After recentrifugation, cell pellets were im-APC/C complexes (Yoon et al. 2002). SWM1was pre- mediately frozen at⫺70⬚.
Flow cytometry:Typically 1 ml of midlog-phase cell culture
viously shown to be required for meiotic spore wall
was harvested for analysis. For cultures grown in synthetic
maturation, whereasMND2was first identified as being
complete media, 5–10l of 10 mg/ml bovine serum albumen
required for propermeioticnucleardivision in a survey
(NEB) was often added to assist initial centrifugation of cells.
of genes whose deletion causes meiotic defects (Ufano Cell pellets were washed with 1 ml ice-cold 0.2mTris pH 7.5
et al.1999;Rabitschet al.2001). Little is known about and then resuspended in 70% ethanol in 0.2mTris pH 7.5 the contribution of either protein to APC/C function. and fixed overnight at 4⬚. Fixed cells were treated with 1 mg/ ml RNAse 0.2mTris pH 7.5 forⵑ4 hr at 37⬚and were then
In this work we assess thein vivorequirement of Swm1p
incubated overnight at 50⬚ after addition of 5–10 l of 20
and Mnd2p, as well as that of Apc9p, another
nonessen-mg/ml Proteinase K. Cells were stained overnight in 5 g/
tial APC/C subunit for which limited functional data ml propidium iodide in 0.2
mTris pH 7.5 at 4⬚and were lightly
exist. sonicated before analysis on a Becton Dickinson FACSSort.
Cell cycle arrest-release experiments:We constructed strains containing APC/C substrates epitope tagged on the carboxy terminus with either 3XHA or 13XMYC in wild-type, swm1⌬, MATERIALS AND METHODS
mnd2⌬, orapc9⌬backgrounds. Cultures were grown in YPD to midlog phase at 25⬚, harvested, and washed twice with
Yeast genetic methods and media:Standard yeast growth
me-dia (YPD) and dropout meme-dia [synthetic complete (SC)] was ddH2O. Cells were then resuspended in YPD with 1⫻ ␣-factor and arrested for ⵑ2 hr or until⬎90% of cells had mating used as described inRoseet al.(1990). Yeast transformations
were performed according to the method ofGietzet al.(1995). projections. Cultures were reharvested and washed twice with ddH2O and then resuspended in YPD and released into the Loss of viability was assessed by replating on YPD at 25⬚ and
scoring microcolonies after overnight growth. Rescue ofswm1⌬ cell cycle at 25⬚after standardizing for culture density. At 15-min intervals, 1 ml of culture was harvested for protein pellets apc9⌬andswm1⌬cdc26⌬synthetic lethality (Table 5) was
accom-plished by mating YAP2888 (swm1⌬::HIS3MX6 pRS316-SWM1) and 1 ml was harvested for FACS analysis.
Protein extracts and Western blot analysis:Except for immu-with YVA433 (apc9⌬::KanMX) or YVA435 (cdc26⌬::KanMX)
con-taining pRS424, pSG2 (pRS424-SWM1), pAP35 (pRS424-CDC16), noprecipitations, whole-cell lysate was prepared by the method described in Goh et al.(2000). Typically, 20–40 l/sample pAP36 (pRS424-CDC23), or pAP37 (pRS424-CDC27). Diploids
were sporulated and Ura⫹ Trp⫹ His⫹G418-resistant spores were loaded on SDS-PAGE gels. When using the wet transfer method, we transferred to PVDF for 4–5 hr at 750 mA, whereas were assessed for their ability to grow on 5-FOA. In some cases,
Trp⫺Ura⫹His⫹G418-resistant spores were tested directly by when using the semidry method we transferred to nitrocellu-lose for 1 hr at 12 V. Immunoblot analysis was typically con-retransforming in the 2vector bearing the relevant gene and
assessing the ability of Trp⫹transformants to grow on 5-FOA ducted as follows: Membranes were preblocked for 30 min in block buffer (5% milk/1⫻ Tris-buffered saline TBS/0.05% media.
Plasmids:pOCF30 (pRS416-GAL1-PDS1) (Cohen-Fixet al. Tween-200, followed by incubation with the primary antibody overnight in the same buffer. Blots were washed with wash 1996) was kindly provided by Doug Koshland. pAP62 was
constructed by subcloning anNheI-PstI fragment containing buffer (1⫻ TBS/0.05% Tween-20, supplemented with up to 0.1% SDS). Membranes were typically incubated with second-the ASE1-3XMYC cassette of pPB338 (generously provided
by David Pellman) into the SpeI-PstI sites of pRS414-GAL1 ary antibody in block buffer for 4 hr at room temperature, followed by extensive washing with wash buffer prior to devel-(Mumberget al.1995). pMT634 (pGAL1-CLN2-HA URA3 LEU2
CEN) (Willemset al.1996) was generously provided by Mike opment.
Immunoprecipitations:Glass-bead lysate was prepared as Tyers. pSG2 was constructed by subcloning aBamHI-NdeI
cas-sette containing theSWM1ORF flanked byⵑ175 bp upstream follows from log-phase cultures grown in YPD. Typically 250– 500 ml of midlog-phase culture were harvested by centrifuga-and 225 bp downstream into pRS424 from the original
TABLE 1
Strains and plasmids
Strain/plasmid Genotype Source
Strain
YPH499 MATahis3-⌬200 ade2-101 ura3-52 lys2-801 leu2-⌬1 trp1⌬63 This study YST74 MATahis3-⌬200 ade2-101 ura3-52 lys2-801 leu2-⌬1 trp1⌬1 cdc23-54 Lambet al.(1994) YJL96 MATahis3-⌬200 ade2-101 ura3-52 lys2-801 leu2-⌬1 trp1⌬63 swm1⌬::HIS3 This study YAP645 MATahis3-⌬200 ade2-101 ura3-52 lys2-801 leu2-⌬1 trp1⌬63 apc11⌬::HIS3 This study
leu2⌬1::apc11-13 pOCF30
YAP928 MATahis3-⌬200 ade2-101 ura3-52 lys2-801 leu2-⌬1 trp1⌬63 This study SWM1-13XMYC; HIS3MX6
YAP1146 MATahis3-⌬200 ade2-101 ura3-52 lys2-801 leu2-⌬1 trp1⌬63 pOCF30 This study YAP1148 MATahis3-⌬200 ade2-101 ura3-52 lys2-801 leu2-⌬1 trp1⌬63 swm1⌬::HIS3 This study
pOCF30
YAP1232 MATahis3-⌬200 ade2-101 ura3-52 lys2-801 leu2-⌬1 trp1⌬63 PDS1-13XMYC; TRP1 This study YAP1273 MATahis3-⌬200 ade2-101 ura3-52 lys2-801 leu2-⌬1 trp1⌬63 pMT634 This study YAP1275 MATahis3-⌬200 ade2-101 ura3-52 lys2-801 leu2-⌬1 trp1⌬63 swm1⌬::HIS3 pMT634 This study YAP1277 MATahis3-⌬200 ade2-101 ura3-52 lys2-801 leu2-⌬1 trp1⌬63 cdc34-2 pMT634 This study YAP1294 MATahis3-⌬200 ade2-101 ura3-52 lys2-801 leu2-⌬1 trp1⌬63 CLB5-3XHA; TRP1 This study YAP1543 MATahis3-⌬200 ade2-101 ura3-52 lys2-801 leu2-⌬1 trp1⌬63 CLB2-3XHA; TRP1 This study YAP1646 MATahis3-⌬200 ade2-101 ura3-52 lys2-801 leu2-⌬1 trp1⌬63 ASE1-3XHA; TRP1 This study YAP1949 MATahis3-⌬200 ade2-101 ura3-52 lys2-801 leu2-⌬1 trp1⌬63 pAP62 This study YAP1951 MATahis3-⌬200 ade2-101 ura3-52 lys2-801 leu2-⌬1 trp1⌬63 swm1⌬::HIS3 pAP62 This study YAP1955 MATahis3-⌬200 ade2-101 ura3-52 lys2-801 leu2-⌬1 trp1⌬63 apc11⌬::HIS3 This study
leu2⌬1::apc11-13 pAP62
YAP2004 MATahis3-⌬200 ade2-101 ura3-52 lys2-801 leu2-⌬1 trp1⌬63 CDC5-3XHA; TRP1 This study YAP2305 MATahis3-⌬200 ade2-101 ura3-52 lys2-801 leu2-⌬1 trp1⌬63 mnd2⌬::KanMX pOCF30 This study YAP2307 MATahis3-⌬200 ade2-101 ura3-52 lys2-801 leu2-⌬1 trp1⌬63 mnd2⌬::KanMX pAP62 This study YAP2340 MATahis3-⌬200 ade2-101 ura3-52 lys2-801 leu2-⌬1 trp1⌬63 swm1⌬::HIS3MX6 This study YAP2343 MATahis3-⌬200 ade2-101 ura3-52 lys2-801 leu2-⌬1 trp1⌬63 swm1⌬::HIS3MX6 This study YAP2346 MATahis3-⌬200 ade2-101 ura3-52 lys2-801 leu2-⌬1 trp1⌬63 swm1⌬::HIS3MX6 This study
pOCF30
YAP2410 MATahis3-⌬200 ade2-101 ura3-52 lys2-801 leu2-⌬1 trp1⌬63 CLB2-3XHA; TRP1 This study swm1⌬::HIS3MX6
YAP2419 MATahis3-⌬200 ade2-101 ura3-52 lys2-801 leu2-⌬1 trp1⌬63 CLB2-3XHA; TRP1 This study mnd2⌬::KanMX
YAP2428 MATahis3-⌬200 ade2-101 ura3-52 lys2-801 leu2-⌬1 trp1⌬63 swm1⌬::HIS3MX6 This study mad2⌬::TRP1
YAP2432 MAT␣his3-⌬200 ade2-101 ura3-52 lys2-801 leu2-⌬1 trp1⌬63 swm1⌬::HIS3MX6 This study mad2⌬::TRP1
YAP2441 MAT␣his3-⌬200 ade2-101 ura3-52 lys2-801 leu2-⌬1 trp1⌬63 swm1⌬::HIS3MX6 This study bub2⌬::TPR1
YAP2443 MATahis3-⌬200 ade2-101 ura3-52 lys2-801 leu2-⌬1 trp1⌬63 swm1⌬::HIS3MX6 This study bub2⌬::URA3
YAP2481 MATahis3-⌬200 ade2-101 ura3-52 lys2-801 leu2-⌬1 trp1⌬63 apc1-21; trp1⌬::HIS3 This study pOCF30
YAP2485 MATahis3-⌬200 ade2-101 ura3-52 lys2-801 leu2-⌬1 trp1⌬63 apc1-21; trp1⌬::HIS3 This study pAP62
YAP2771 MATahis3-⌬200 ade2-101 ura3-52 lys2-801 leu2-⌬1 trp1⌬63 apc9⌬::TRP1 pOCF30 This study YAP2775 MATahis3-⌬200 ade2-101 ura3-52 lys2-801 leu2-⌬1 trp1⌬63 cdc26⌬::TRP1 pOCF30 This study YAP2821 MATahis3-⌬200 ade2-101 ura3-52 lys2-801 leu2-⌬1 trp1⌬63 PDS1-13XMYC; TRP1 This study
swm1⌬::HIS3MX6
YAP2824 MATahis3-⌬200 ade2-101 ura3-52 lys2-801 leu2-⌬1 trp1⌬63 PDS1-13XMYC; TRP1 This study mnd2⌬::KanMX
YAP2827 MATahis3-⌬200 ade2-101 ura3-52 lys2-801 leu2-⌬1 trp1⌬63 CLB5-3XHA; TRP1 This study swm1⌬::HIS3MX6
YAP2830 MATahis3-⌬200 ade2-101 ura3-52 lys2-801 leu2-⌬1 trp1⌬63 CLB5-3XHA; TRP1 This study mnd2⌬::KanMX
YAP2861 MATahis3-⌬200 ade2-101 ura3-52 lys2-801 leu2-⌬1 trp1⌬63 ASE1-3XHA; TRP1 This study swm1⌬::HIS3MX6
YAP2864 MATahis3-⌬200 ade2-101 ura3-52 lys2-801 leu2-⌬1 trp1⌬63 CDC5-3XHA; TRP1 This study swm⌬::HIS3MX6
TABLE 1
(Continued)
Strain/plasmid Genotype Source
YAP2873 MATahis3-⌬200 ade2-101 ura3-52 lys2-801 leu2-⌬1 trp1⌬63 CLB5-3XHA; TRP1 This study swm1⌬::HIS3MX6 pds1⌬::KanMX
YAP2934 MATahis3-⌬200 ade2-101 ura3-52 lys2-801 leu2-⌬1 trp1⌬63 CLB2-3XHA; TRP1 This study apc9⌬::KanMX
YAP2937 MATahis3-⌬200 ade2-101 ura3-52 lys2-801 leu2-⌬1 trp1⌬63 PDS1-13XMYC; TRP1 This study apc9⌬::KanMX
YAP2941 MATahis3-⌬200 ade2-101 ura3-52 lys2-801 leu2-⌬1 trp1⌬63 CLB5-3XHA; TRP1 This study apc9⌬::KanMX
YVA387 MAT␣his3-⌬200 ade2-101 ura3-52 lys2-801 leu2-⌬1 trp1⌬63 mad2⌬::TRP1 This study YVA388 MAT␣his3-⌬200 ade2-101 ura3-52 lys2-801 leu2-⌬1 trp1⌬63 mad2⌬::TRP1 This study YVA389 MAT␣his3-⌬200 ade2-101 ura3-52 lys2-801 leu2-⌬1 trp1⌬63 bub2⌬::URA3 This study YVA390 MAT␣his3-⌬200 ade2-101 ura3-52 lys2-801 leu2-⌬1 trp1⌬63 bub2⌬::URA3 This study YVA1048 MATahis3-⌬200 ade2-101 ura3-52 lys2-801 leu2-⌬1 trp1⌬63 apc1-21; TRP1 This study YVA1098 MATahis3-⌬200 ade2-101 ura3-52 lys2-801 leu2-⌬1 trp1⌬63 apc1-21; trp1⌬::HIS3 This study YVA1114 MATahis3-⌬200 ade2-101 ura3-52 lys2-801 leu2-⌬1 trp1⌬63 mnd2⌬::KanMX This study YVA1130 MATahis3-⌬200 ade2-101 ura3-52 lys2-801 leu2-⌬1 trp1⌬63 MND2-13XMYC; TRP1 This study
Plasmid
pMT634 pGAL1-CLN2-HA URA3 LEU2 CEN M. Tyers
pSG2 pRS424-SWM1 S. Givan
pVA117 pRS426-MND2 This study
pOCF30 pRS416-GAL1-PDS1 D. Koshland
pAP62 pRS414-GAL1-ASE1-3XMYC This study, derived from
pPB338 from D. Pellman
with 2g/ml aprotinin, 0.5g/ml leupeptin, 0.7g/ml pep- was subcloned into pRSETB and expressed in bacteria. The resulting 6-HIS fusion protein was purified over a Ni ⫹ 2 statin A, 1 mmPMSF, or with a EDTA-free complete protein
inhibitor cocktail tablet (Roche). A volume of glass beads column and injected into rabbits to produce polyclonal anti-bodies (Covance).
approximately equal to the volume of cells harvested was added and cells were subjected to 16 cycles of vortexing by hand for 30 sec followed by a 30-sec incubation on ice. After
centrifugation for 10 min at 4⬚at 13,000 rpm in a JA-20 rotor RESULTS (Beckman, Fullerton, CA), lysate was transferred to Eppendorf
tubes and cleared for 5–10 min at 14,000 rpm at 4⬚ in an Identification of SWM1andMND2:To identify new
Eppendorf centrifuge. Cleared lysate was transferred to Ep- components and regulators of the budding yeast ana-pendorf tubes, to which were added 50–100 l of anti- phase-promoting complex, we conducted screens for mycconjugated beads (Covance). Immunoprecipitations were
high-copy suppressors of several temperature-sensitive
conducted overnight at 4⬚ on an end-over-end rocker. The
mutants in previously identified APC/C components.
beads were pelleted in an Eppendorf centrifuge at 2000 rpm
In separate multi-copy suppression screens we identified
with tubes rotated 180⬚between spins to enhance pelleting.
After removal of the supernatant, beads were washed five times SWM1as a dosage suppressor ofcdc23-54andMND2as in 1 ml ice-cold extract buffer, with each wash comprising 3 a dosage suppressor ofapc1-21.cdc23-54is a previously min of agitation on the end-over-end rocker, followed by the
described allele ofCDC23(Sikorskiet al.1993) whereas
same centrifugation regimen. After the final wash, 250–300
apc1-21 is a new allele of APC1 constructed by PCR l of 2⫻Laemmli lacking-mercaptoethanol was added to
mutagenesis. Upon shift to 37⬚,apc1-21predominantly
the bead pellet, which was then incubated at 42⬚for 5–10 min.
The beads were then transferred to an NA-45 spin column arrests as large-budded cells with short mitotic spindles
and spun at⫺7500 rpm for 5 min, followed by addition of and DAPI-staining masses at the bud neck, an arrest -mercaptoethanol. Samples were boiled for 3 min and then similar to other temperature-sensitive mutants in essen-immediately frozen at ⫺70⬚. Antibodies against Cdc16p,
tial APC/C components (Figure 1A).apc1-21mutants are
Cdc27p, and Cdc23p are described inLambet al.(1994).
also defective in APC/C substrate proteolysis in␣
-factor-Immunofluorescence:Cells were fixed for 90 min with
one-tenth volume 37% formaldehyde at 37⬚, washed one to two arrested cells (see Figure 3). A 2plasmid containing
times in 1 ml SK (1 msorbitol, 50 mmKPO4pH 7.5) and SWM1(pSG2) suppressed the temperature sensitivity of resuspended in SK. Cells were stained for microtubules with cdc23-54 at 35⬚, and a 2 plasmid containing MND2 rat antitubulin (Yol 1/34) and fluorescein-conjugated
goat-(pVA117) suppressed the temperature sensitivity of
anti-rat. (GAN-F).
apc1-21 at 33⬚ (Figure 1B). Consistent with previous
re-Anti-Apc2p antibodies: A 348-bp BamHI-KpnI fragment
Figure1.—Identification ofSWM1andMND2as new budding yeast APC/C components. (A) Terminal arrest phenotype of apc1-21. Upon shift to 37⬚for 3 hr, the majority ofapc1-21cells (YVA1098) arrest with large buds, short mitotic spindles, and DAPI-staining masses at or near the bud neck. (B)SWM1andMND2were identified as multicopy suppressors ofcdc23-54and apc1-21, respectively, in independent suppressor screens. A 2vector containing theSWM1open reading frame suppresses the temperature sensitivity ofcdc23-54(YST74) at 35⬚and a 2vector containingMND2suppresses the temperature sensitivity of apc1-21(YVA1048) at 33⬚. (C) Flow cytometry analysis. A wild-type strain (YPH499),swm1⌬(YAP2340), andmnd2⌬(YVA1114) were cultured at 25⬚and shifted to 37⬚for 3 hr. Aliquots were harvested and analyzed by flow cytometry or fixed and stained with antitubulin (Yol 1/34) and DAPI. At 25⬚,swm1⌬has a significant 2N accumulation, whereas the profile ofmnd2⌬is similar to that of wild type. Upon shift to 37⬚for 3 hr, the 2N accumulation ofswm1⌬, but notmnd2⌬, is further exacerbated. (D) Upon shift to 37⬚, the number of large-budded cells with short mitotic spindles at or near the bud neck is approximately threefold higher inswm1⌬compared to wild type.
and Mnd2p in APC/C complexes (Yoon et al. 2002; (up to 12 hr) does not cause significant loss of viability inswm1⌬mutants (data not shown).mnd2⌬mutants in Hallet al.2003;Schwickartet al.2004), both
Swm1p-13XMYC and Mnd2p-Swm1p-13XMYC immunoprecipitated the S288c background are not temperature sensitive at 37⬚. Levels of both Swm1p and Mnd2p proteins re-other APC/C subunits (see supplemental Figure S1, A
and B, at http://www.genetics.org/supplemental/). mained steady and did not fluctuate during the mitotic cell cycle (see supplemental Figure S2 at http://www.
swm1⌬andmnd2⌬deletion phenotypes:Deletion of
swm1⌬in the S288c background is viable from 25⬚ to genetics.org/supplemental/).
Swm1p but not Mnd2p is required for efficient turn-35⬚, butswm1⌬strains grow poorly at 37⬚, arresting after
one to five cell divisions.swm1⌬mutants grown at 25⬚ over of specific APC/C substrates: To assess the role of Swm1p and Mnd2p in APC/C substrate turnover, we in liquid culture are characterized by an accumulation
in G2/M DNA content, which is exacerbated after shift- monitored the degradation of several APC/C substrates
in␣-factor-arrested cells, where the APC/C is constitu-ing to 37⬚for 3 hr, whereas the FACS profile ofmnd2⌬
cultures does not significantly change (Figure 1C). In tively active against its known substrates (Amon et al.
1994;Irniger andNasmyth 1997; Juanget al. 1997;
swm1⌬cultures at 37⬚, we typically observed a threefold
increase in the number of large-budded cells with short Charles et al. 1998). We expressed Pds1p or Ase1p-3XMYC from the GAL1 promoter in cells arrested in mitotic spindles and DAPI-staining masses at the bud
neck, whereasmnd2⌬strains did not accumulate large- G1with␣-factor and then repressed protein expression
with the addition of glucose and cycloheximide. Both budded cells with short spindles upon shift to 37⬚
Figure2.—Swm1p is required for APC/C activ-ity in␣-factor-arrested cells. (A) Pds1p wild-type (YAP1146), swm1⌬(YAP1148), or apc11-13 (YAP-645) strains bearing pOCF30 (pRS416-GAL1-PDS1) were cultured to midlog phase at 25⬚ in SC-Ura 2% Raf supplemented with additional adenine and tryptophan and arrested for 3.5 hr with 5g/ ml ␣-factor at 25⬚. Cultures were shifted to 37⬚ for 15 min and then induced with 2% galactose for another 30 min at 37⬚. Two percent glucose and 1 mg/ml cycloheximide was added to repress transcription and translation of the GAL1 pro-moter, and aliquots were harvested for protein extracts and flow cytometry every 15 min. The “Raf” time point was harvested immediately be-fore shift to 37⬚ and the “Gal” time point was harvested immediately before addition of glucose and cycloheximide. Western blots were probed with anti-Pds1p (C210) and reprobed with anti-Cdc28p as a loading control. (B) Wild-type (YAP1949), swm1⌬ (YAP1951), and apc11-13 (YAP1955) cul-tures all bearing pAP62 ( pRS414-GAL1-ASE1-3XMYC) were cultured in SC-Trp 2% Raf supple-mented with additional adenine and uracil and arrested as in A. Western blots were probed with anti-MYC (9E10). An anti-Apc2p (BC039) cross-reacting band is used as a loading control. The asterisk represents an unknown anti-Apc2p cross-reacting band used as a loading control. (C) Wild type (YAP1146) andswm1⌬(YAP1148) were cul-tured to midlog phase in SC-Ura 2% Raf supple-mented with additional adenine and tryptophan at 30⬚and arrested with 5g/ml␣-factor for 3.5 hr. Cultures were induced with 2% galactose for 30 min and 2% glucose and 1 mg/ml cyclohexi-mide was added to repress GAL1 transcription and translation. Aliquots for cell pellets and flow cytometry were harvested every 15 min and processed as in A. (D) YAP1273 (wild type), YAP1275 (swm1⌬), and YAP1277 (cdc34-2), all bearing pMT634 (pGAL1-CLN2-HA), were grown to midlog phase in SC-Ura 2% Raf supplemented with extra adenine and tryptophan. Cultures were arrested for 3.5 hr in 0.1mhydroxyurea at 25⬚, then 2% galactose was added, and cultures were shifted to 37⬚for 45 min. After galactose induction of Cln2p-3XHA, 2% glucose and 1 mg/ml cycloheximide were added to repressGAL1transcription and translation and aliquots were harvested for cell pellets and flow cytometry every 15 min. The “Raf” time point was harvested immediately before galactose induction and shift to 37⬚. Whole-cell lysate was prepared as described inGohet al.(2000). Western blots were probed with anti-HA (12CA5) and reprobed with anti-Cdc28p as a loading control.
and do not break through into S-phase. Maintenance of to degrade Pds1p, with a significant portion of the protein still remaining at the 60-min time point (Figure 2C; see G1arrest was verified by FACS analysis in all experiments
(data not shown). In wild-type strains, both Pds1p and also Figure 4).
To rule out a role for Swm1p in general ubiquitin-Ase1p-3XMYC are nearly undetectable 15–20 min after
repression of their expression (Figure 2, A and B). In mediated proteolysis, we assessed the role of Swm1p in the turnover of the G1cyclin Cln2p. Cln2p degradation
contrast, in swm1⌬ the accumulated Pds1p persisted
throughout the duration of the 60-min chase period, requires both SCFGrr1 and Cdc34p, an SCF-associated
ubiquitin-conjugating enzyme (E2) (Deshaies et al.
similar to the positive control,apc11-13, with little
degra-dation in either strain. For Ase1p-3XMYC, in bothswm1⌬ 1995;Willemset al.1996;KishiandYamao1998; Pat-ton et al. 1998). Cln2p is unstable in hydroxyurea-andapc11-13, Ase1p-3XMYC accumulated to higher
lev-els than in wild type, but its level decreased slowly over arrested cells, when the APC/C is inactive against its known substrates (Willemset al.1996). Wild-type,swm1⌬, the course of the chase period. In our assays we observed
only partial stabilization of ectopically expressed Ase1p- and cdc34-2 cells were arrested in hydroxyurea and Cln2p-3XHA was expressed from theGAL1 promoter. 3XMYC, consistent with the observations ofThornton
andToczyski(2003) who reported incomplete stabili- Thirty minutes after repression of theGAL1promoter,
the bulk of Cln2p-3XHA was degraded in both wild type zation of Ase1p in Apc⫺strains and proposed the
Figure3.—Mnd2p is dispensable for APC/C activity in ␣-factor-arrested cells. (A) Wild-type (YAP1146),mnd2⌬(YAP2305), andapc1-21 (YAP-2841) strains containing pOCF30 were grown to midlog phase at 25⬚in SC-Ura 2% Raf media sup-plemented with additional adenine and trypto-phan. Cultures were arrested at 25⬚with 5g/ml ␣-factor for 3.5 hr and then shifted to 37⬚for 15 min. Two percent galactose was added to induce PDS1expression for 30 min at 37⬚. Two percent glucose and 1 mg/ml cycloheximide were then added to repressGAL1transcription and transla-tion, and aliquots for cell pellets and flow cytome-try were harvested every 20 min and processed as in Figure 2A. (B) Wild-type (YAP1949), mnd2⌬ (YAP2307), and apc1-21 (YAP2485) strains con-taining pAP62 were grown to midlog phase at 25⬚ in SC-Trp 2% Raf media supplemented with additional uracil and adenine. Cultures were arrested with 5g/ml␣-factor at 25⬚for 3 hr and 45 min and then shifted to 37⬚ for 15 min. Cultures were induced with 2% galactose for 30 min at 37⬚, and then 2% glucose and 1 mg/ml cycloheximide was added to repressGAL1transcription and translation. Aliquots for cell pellets and flow cytometry were harvested every 20 min and processed as in Figure 2A. The asterisk represents an unknown anti-Apc2p cross-reacting band used as a loading control.
dase-ubiquitin-fusion construct, which is dependent on andapc9⌬, after an initial drop in Pds1p levels, a fraction of Pds1p remains undegraded for the duration of the the N-end rule pathway for its proteolysis (Bachmair
et al.1986;Varshavsky1996), was also rapidly degraded time course. Compared to theswm1⌬mutant, in which a substantial portion of Pds1p remains undegraded after in a swm1⌬ strain (data not shown). Collectively, our
results suggest that Swm1p is specifically required for 60 min, in the apc9⌬ mutant only a small fraction of Pds1p is refractory to degradation during the entire APC/C-dependent ubiquitination and is not required
for general ubiquitin-mediated proteolysis. time course. Similar to swm1⌬, in the cdc26⌬ mutant, Pds1p accumulates to higher levels than in the wild-type In contrast to the APC/C substrate stabilization
exhib-ited byswm1⌬,mnd2⌬mutants failed to stabilize either strain, but its levels diminish gradually over the course of the experiment (Figure 4B). Our results demonstrate Pds1p (Figure 3A) or Ase1p-3XMYC (Figure 3B). Both
substrates were rapidly degraded after 20 min to levels that, at permissive temperatures, nonessential APC/C subunits exhibit substantial differences in the degrada-similar to that of the wild-type strain, whereas in an
apc1-21mutant, levels of both substrates persisted through- tion of ectopically expressed Pds1p and that the contri-bution of each subunit to overall APC/C activity varies out the duration of the time course. Our results
dem-onstrate that Swm1p, but not Mnd2p, is required for significantly.
swm1⌬ mutants but not mnd2⌬ mutants exhibit de-efficient APC/C function in␣-factor-arrested cells.
In galactose shutoff experiments using Pds1p as an fects in APC/C substrate degradation through the cell cycle:APC/CCdc20pactivity is required for the timely
deg-APC/C substrate,swm1⌬mutants consistently exhibited
stabilization of ⵑ50–70% of the initially expressed radation of Pds1p, Clb5p, and a fraction of the total pool of Clb2p (Gohet al.2000;Irniger2002), and in Pds1p protein. This stabilization was not temperature
dependent as we observed this phenomenon at both the strains where Clb2p is degradable by APC/CCdh1p,
dele-tion of bothCLB5andPDS1makes Cdc20p dispensable permissive temperature forswm1⌬growth (30⬚) and the
nonpermissive temperature (37⬚). To ascertain whether for cellular viability (Shirayamaet al.1999;Waschand
Cross2002; ThorntonandToczyski 2003). Having
other nonessential APC/C subunits exhibit this
phe-nomenon, we conducted similar experiments inapc9⌬ demonstrated a requirement for Swm1p, but not for Mnd2p, for full APC/C activity against ectopically ex-andcdc26⌬. In the S288c background, apc9⌬mutants
are viable at 37⬚, whereascdc26⌬mutants grow normally pressed substrates in␣-factor-arrested cells, we next ex-amined whether strains lacking either Swm1p or Mnd2p at 30⬚ but fail to grow at 33⬚ and above. In galactose
shutoff assays conducted at 30⬚, we observed substantial are defective in APC/C proteolysis of tagged substrates expressed from their endogenous promoters over the differences in the degradation pattern of Pds1p among
swm1⌬,cdc26⌬, and apc9⌬ mutants (Figure 4A). From course of the cell cycle. In both the wild-type strain and
swm1⌬, Clb2p-3XHA appeared 45 min after release from quantification of the amount of Pds1p in the “Gal” time
point of each strain in several experiments, we estimate ␣-factor. In the wild-type strain, Clb2p-3XHA levels de-clined 105 min after release as the cells entered ana-that, inswm1⌬andcdc26⌬, the starting amount of Pds1p
tion was delayed by ⵑ15 min with the Clb5p-3XHA reaching its lowest level at 105 min. The delay in Clb5p-3XHA degradation was not due to the failure ofswm1⌬
to efficiently degrade Pds1p as the timing of Clb5p-3XHA proteolysis was still delayed by 15 min in the
swm1⌬pds1⌬double mutant (Figure 5C).
We also tested the requirement of Swm1p in the deg-radation of two APC/CCdh1psubstrates, Cdc5p-3XHA and
Ase1p-3XHA. Like Clb2p, in theswm1⌬mutant Cdc5p-3XHA levels remained steadily high throughout the cell cycle (Figure 5E), whereas Ase1p-3XHA exhibited a pro-teolysis defect but was not completely stabilized (Figure 5D). Our results demonstrate that Swm1p is required for the timely degradation of both APC/CCdc20p and
APC/CCdh1psubstrates and that swm1⌬mutants exhibit
significant proteolysis defects even at permissive temper-atures.
We conducted similar experiments to assess the tim-ing of APC/C substrate degradation inmnd2⌬mutants. In contrast toswm1⌬, we did not observe any significant differences in the timing of onset of Clb2p-3XHA (Fig-ure 6A), Pds1p-13XMYC (Fig(Fig-ure 6B), and Clb5p-3XHA (Figure 6C) degradation. These results are consistent with our failure to observe stabilization of ectopically expressed APC/C substrates in␣-factor-arrestedmnd2⌬
cultures and suggest that Mnd2p is largely dispensable
Figure4.—Deletions ofSWM1,APC9, andCDC26have
sub-for APC/C substrate proteolysis in the absence of other
stantially different APC/C activity in ␣-factor-arrested cells.
APC/C defects.
(A) Wild-type (YAP1146),swm1⌬(YAP2346),apc9⌬(YAP2771),
and cdc26⌬(YAP2775) strains bearing pOCF30 were grown Timing of Clb2p but not Pds1p or Clb5p degradation to midlog phase at 25⬚in SC-Ura 2% Raf supplemented with is delayed inapc9⌬mutants:Given our failure to observe additional tryptophan and adenine. Cells were arrested with significant stabilization of ectopically expressed Pds1p 5g/ml␣-factor at 25⬚for 3 hr and 45 min and then shifted
in apc9⌬ cultures arrested in ␣-factor, we assessed the
to 30⬚for 15 min. Cultures were induced with 2% galactose
timing of APC/C substrate degradation in apc9⌬over
for 30 min at 30⬚and then 2% glucose and 1 mg/ml
cyclohexi-mide was added to repressGAL1transcription and translation. the mitotic cell cycle. Onset of Clb2-3XHA degradation
Maintenance of␣-factor-arrested cells was verified by flow cytom- (Figure 7A) was consistently delayed 15 min in apc9⌬, etry (data not shown). Western blots were probed with
anti-but the amount of Clb2p-3XHA eventually declined to
Pds1p (C210, D. Koshland) and reprobed with anti-Cdc28p as
the same level as that in the wild-type strain. In contrast,
a loading control. (B) Pds1p levels were quantified on a
Bio-the timing of Pds1p-13MYC and Clb5p-3XHA
degrada-Rad (Richmond, CA) Fluor-S multi-imager.
tion inapc9⌬was nearly identical to that of wild type, with the disappearance of both proteins occurring by 105 min (Figure 7, B and C). Our findings suggest that ure 5A). For Pds1p-13XMYC, we observed initial accu- the requirement for Apc9p in APC/C function may be mulation of Pds1p-13XMYC protein 30 min after release limited to the degradation of only some APC/C sub-from ␣-factor in wild type and initiation of Pds1p- strates and not others. Alternatively, APC/C complexes 13XMYC turnover atⵑ90 min, with the bulk of Pds1p- lacking Apc9p may retain sufficient activity to degrade 13XMYC protein having been degraded by 105 min. Pds1p and Clb5p efficiently, but not Clb2p.
In swm1⌬however, Pds1p-13XMYC levels decline only Spindle elongation is delayed in swm1⌬ and apc9⌬
slightly during the onset of anaphase, with substantial mutants: In our arrest-release experiments, we consis-levels persisting at 90 and 105 min. Furthermore, we con- tently observed inswm1⌬, and to a lesser extent inapc9⌬, sistently observed undegraded Pds1p-13XMYC in␣-factor- a delay in reaccumulation of 1N content in FACS pro-arrested cells and at the 15-min time point (Figure 5B). files. Typically 1N content began to reappear at 120 min In the case of Clb5p, we observed significant accumu- in wild-type and mnd2⌬ strains but not until 135 min lation of Clb5p-3XHA protein 15 min after release from inswm1⌬andapc9⌬. To ascertain whether any of these ␣-factor in both wild type andswm1⌬. In the wild-type mutants are delayed in early or late anaphase, we ar-strain, Clb5p-3XHA levels began to decline 75 min after rested cultures in␣-factor, as in Figure 2, and released release from␣-factor and reached their lowest level at them into the mitotic cell cycle at 25⬚. In swm1⌬and
Figure5.—Continued.
delayed byⵑ15 min compared to wild-type andmnd2⌬. tative growth, in three other nonessential budding yeast APC/C subunits:apc9⌬,cdc26⌬, anddoc1⌬.swm1⌬was By 105 min, the number of short spindles in apc9⌬was
comparable to that of wild type andmnd2⌬, but inswm1⌬ also synthetically lethal withcdc23-1andcdc23-54, as well as theCDC28-VFmutant, which reduces APC/C activity a significant number of cells had short spindles at 120
min (Figure 7D). Likewise, onset of spindle elongation and has been shown to be synthetically lethal with other APC/C mutants (Rudneret al. 2000). In addition, we (Figure 7E) occurred between 75 and 120 min for wild
type andmnd2⌬and between 90 and 120 min forapc9⌬. assessed the genetic interactions ofswm1⌬with a dele-tion in eitherUBC4orCLB2. Ubc4p is an E2-ubiquitin-Inswm1⌬, appearance of elongated spindles was delayed
by ⵑ15 min with the peak occurring at 105–120 min. conjugating enzyme that can activate the APC/Cin vitro
(Charles et al. 1998) and ubc4⌬ mutants have been These results are consistent with our observations that
swm1⌬mutants are defective in the degradation of a broad shown to be synthetically lethal with the apc mutants
cdc23-1anddoc1⌬(Irnigeret al.1995, 2000). In addi-spectrum of both APC/CCdc20pand APC/CCdh1psubstrates,
whereasapc9⌬mutants exhibit only a delay in the degrada- tion to being an APC/C substrate, Clb2p directs the phosphorylation of APC/C TPR subunits by its partner tion of Clb2p and not Pds1p or Clb5p.
Genetic analysis of swm1⌬, mnd2⌬, and apc9⌬ mu- kinase Cdc28p (RudnerandMurray2000), andclb2⌬
is synthetically lethal withcdc23-1(Irnigeret al.1995). tants:We examined the genetic interactions ofswm1⌬
with mutations in other APC/C components and regula- Both swm1⌬ ubc4⌬ and swm1⌬ clb2⌬ double mutants were viable, but exhibited conditional synthetic lethality tors (Table 2).swm1⌬exhibited synthetic lethality with
in-Figure6.—Deletion ofMND2does not affect degradation of APC/C substrates during the mi-totic cell cycle.CLB2-3XHA(YAP1543) and CLB2-3XHA mnd2⌬ (YAP2419) (A), PDS1-13XMYC (YAP1232) andPDS1-13XMYC mnd2⌬(YAP2824) (B), andCLB5-3XHA(YAP1294) andCLB5-3XHA mnd2⌬(YAP2830) (C) were arrested with␣-factor and released into the cell cycle as described in Figure 5A. Deletion ofMND2did not substantially affect the timing of Clb2p-3XHA, Pds1p-13XMYC, or Clb5p-3XHA degradation. Asterisks in B repre-sent anti-Cdc28p cross-reacting bands used as a loading control.
teraction withclb5⌬. We also assessed genetic interac- acts in parallel with the APC/C to inhibit cyclin B-CDK activity at exit from mitosis (Mendenhall1993; Dono-tions betweenswm1⌬and the APC/C specificity factors.
swm1⌬was synthetically lethal with thecdc20-1mutation, van et al. 1994; Nugroho and Mendenhall 1994; Schwobet al. 1994). sic1⌬mutations are synthetically butswm1⌬cdh1⌬double mutants were viable at
Figure7.—APC/C substrate degradation in apc9⌬. CLB2-3XHA(YAP1543) andCLB2-3XHA apc9⌬(YAP2934) (A),PDS1-13XMYC(YAP1232) and PDS1-13XMYC apc9⌬(YAP2937) (B), and CLB5-3XHAandCLB5-3XHA apc9⌬(YAP2941) (C) were arrested with␣-factor and released into the cell cycle as described in Figure 5A.apc9⌬ delayed the onset of Clb2p-3XHA proteolysis but did not significantly affect the timing of PDS1-13XMYC and Clb5p-3XHA degradation. Aster-isks in B represent anti-Cdc28p cross-reacting bands. (D and E) Wild-type (YPH499),swm1⌬ (YAP2340),apc9⌬(YAP2930), andmnd2⌬ (YVA-1114) were grown to midlog phase in YEPD, arrested with␣-factor, and released into the cell cycle as described in Figure 5A. Samples were harvested and fixed for immunofluorescence and stained with antitubulin (Yol 1/34) and DAPI. Short and long mitotic spindles were scored for each time point (n⫽100).
et al. 2001; Cross2003), as well as with temperature- lethality withdoc1⌬at 33⬚. However, we failed to observe genetic interactions amongapc9⌬andubc4⌬,cdh1⌬, and sensitive mutations inCDC23and APC2(Schwabet al.
1997; Krameret al. 1998). Crosses between sic1⌬and mnd2⌬.apc9⌬in combination withcdc26⌬was also viable and did not lower the nonpermissive temperature of the
swm1⌬strains generated viablesic1⌬swm1⌬double
mu-tants, although ⵑ40% ofsic1⌬swm1⌬ spores failed to cdc26⌬ single mutant. APC9 and SIC1 are somewhat closely linked (ⵑ35 kb apart), but in crosses between germinate. Survivingsic1⌬swm1⌬double mutants were
viable at 35⬚. sic1⌬andapc9⌬, we were able to recover a few haploid
double-mutant spores that were viable at 35⬚.
mnd2⌬mutants exhibited genetic interactions with a
narrower spectrum of APC/C components and regula- Deletion of mad2⌬ partially suppresses the poor growth ofswm1⌬:We examined the genetic relationship tors thanswm1⌬mutants (Table 2).mnd2⌬was
syntheti-cally lethal withapc1-21and reduced the nonpermissive of Swm1p and Mnd2p both to the spindle checkpoint, which inhibits APC/CCdc20pbefore the onset of anaphase
temperatures ofcdc26⌬to 30⬚anddoc1⌬to 35⬚,
respec-tively.apc2-402 mnd2⌬double mutants grew somewhat in response to spindle damage or lack of microtubule attachment to kinetochores (Straight and Murray slower at 33⬚ than apc2-402 single mutants. However,
mnd2⌬ did not exhibit significant genetic interactions 1997;SkibbensandHieter1998;ShahandCleveland 2000;Gorbsky2001;Yu2002), and to the mitotic exit with swm1⌬, apc9⌬, ubc4⌬, cdh1⌬, and CDC28-VF and
did not lower the nonpermissive temperature ofcdc20-1 checkpoint, which inhibits release of Cdc14p from the nucleolus and Cdh1p activation until the spindle is orapc11-13(Table 3). These findings suggest either that
the loss of Mnd2p is not particularly deleterious to properly positioned (Li 1999; Bloecher et al. 2000;
Pereira et al. 2000; Wang et al. 2000; Adames et al.
APC/C function in general or that only a limited
num-ber of APC/C subunits depend on Mnd2p for their full 2001;Lew andBurke2003). swm1⌬mutants respond normally to benomyl whereasmnd2⌬mutants are mildly activity.
apc9⌬mutations (Table 4) exhibited synthetic lethal- sensitive (data not shown). Neitherswm1⌬normnd2⌬
Figure7.—Continued.
chromosome fragment.swm1 mad2⌬andswm1⌬bub2⌬ wild-typeSWM1gene was confirmed by PCR and by the observation that swm1⌬ apc9⌬ 2-CDC27 and swm1⌬
double mutants were viable at 35⬚(Table 2), andmnd2⌬
mad2⌬andmnd2⌬bub2⌬mutants grew well at 37⬚(Ta- cdc26⌬ 2-CDC16 strains grow poorly at 30⬚, which is a permissive temperature for all three single mutants ble 3). Deletion ofswm1⌬ormnd2⌬did not significantly
affect the benomyl sensitivity of eithermad2⌬orbub2⌬ (Table 5). In addition, the temperature sensitivity of
apc9⌬cdc26⌬at 33⬚and 35⬚was rescued by overexpres-mutants. However,mad2⌬, but notbub2⌬, partially
sup-pressed the poor growth phenotype ofswm1⌬mutants sion of CDC16, but notCDC23,CDC27, orSWM1. Our findings show that the lethality of swm1⌬ apc9⌬ and at 37⬚(see supplemental Figure S3 at http://www.gene
tics.org/supplemental/). This result suggests that the swm1⌬cdc26⌬can be rescued by overexpression of spe-cific TPR proteins.
apcdefect ofswm1⌬mutants is partially relieved when the inhibitory effect of Mad2p on the APC/C is abro-gated.
DISCUSSION Overexpression of specific TPR proteins rescues the
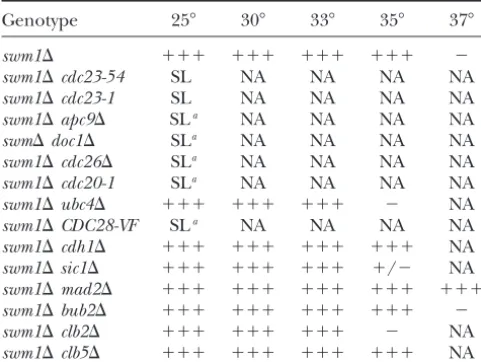
synthetic lethality ofswm1⌬apc9⌬and swm1⌬cdc26⌬ In wild-type cells, the ubiquitin-ligase activity of the anaphase-promoting complex against B-type cyclins and double mutants:To better understand the nature of the
lethal defects ofswm1⌬apc9⌬andswm1⌬cdc26⌬double Pds1p is essential for normal progression from meta-phase to anameta-phase and into G1 of the next cell cycle.
mutants, we examined whether overexpression of any
of the three TPR proteins could rescue the inviability Nevertheless, a surprising number of APC/C subunits are not essential for cell viability in budding yeast under of either double mutant. The temperature sensitivity of
cdc26⌬single mutants was rescued by a 2plasmid con- normal growth conditions, and their contribution to APC/C functionin vivothus far has been poorly charac-tainingCDC16, but not CDC23, or CDC27 (Figure 8).
Overexpression ofAPC2,APC11,CDC20,CDH1,APC5, terized. In this work we have shown thatswm1⌬,apc9⌬, and mnd2⌬ each have different effects on the
degra-APC9,APC1, or SWM1likewise failed to rescuecdc26⌬
(data not shown). We observed rescue ofswm1⌬apc9⌬ dation of APC/C substrates. We identified Swm1p and Mnd2p as new components of the budding yeast double mutants specifically by a 2 plasmid carrying
CDC27, but not CDC16 or CDC23, whereas only 2- APC/C by screening for multicopy suppressors of exist-ingapcmutants. We have demonstrated that at
TABLE 3 TABLE 2
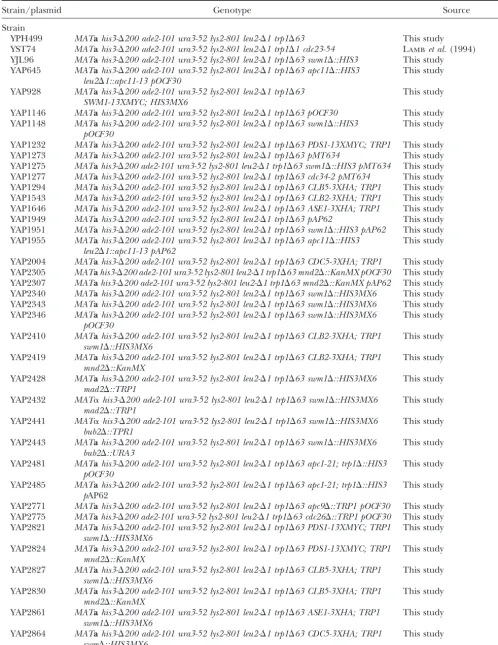
swm1⌬genetics mnd2⌬genetics
Genotype 25⬚ 30⬚ 33⬚ 35⬚ 37⬚ Genotype 25⬚ 30⬚ 33⬚ 35⬚ 37⬚
swm1⌬ ⫹⫹⫹ ⫹⫹⫹ ⫹⫹⫹ ⫹⫹⫹ ⫺ mnd2⌬ ⫹⫹⫹ ⫹⫹⫹ ⫹⫹⫹ ⫹⫹⫹ ⫹⫹⫹
mnd2⌬apc1-21 SLa NA NA NA NA
swm1⌬cdc23-54 SL NA NA NA NA
swm1⌬cdc23-1 SL NA NA NA NA mnd2⌬swm1⌬ ⫹⫹⫹ ⫹⫹⫹ ⫹⫹⫹ ⫹⫹⫹ ND mnd2⌬apc9⌬ ⫹⫹⫹ ⫹⫹⫹ ⫹⫹⫹ ⫹⫹⫹ ND swm1⌬apc9⌬ SLa NA NA NA NA
swm⌬doc1⌬ SLa NA NA NA NA mnd2⌬cdc26⌬ ⫹⫹⫹ ⫺ NA NA NA
mnd2⌬doc1⌬ ⫹⫹⫹ ⫹⫹ ⫹ ⫺ NA swm1⌬cdc26⌬ SLa NA NA NA NA
swm1⌬cdc20-1 SLa NA NA NA NA mnd2⌬apc2-402 ⫹⫹⫹ ⫹⫹⫹ ⫹ NA NA
mnd2⌬apc11-13 ⫹⫹⫹ ⫹⫹⫹b NA NA NA
swm1⌬ubc4⌬ ⫹⫹⫹ ⫹⫹⫹ ⫹⫹⫹ ⫺ NA
swm1⌬CDC28-VF SLa NA NA NA NA mnd2⌬cdh1⌬ ⫹⫹⫹ ⫹⫹⫹ ⫹⫹⫹ ⫹⫹⫹ ND
mnd2⌬cdc20-1 ⫹⫹⫹b NA NA NA NA
swm1⌬cdh1⌬ ⫹⫹⫹ ⫹⫹⫹ ⫹⫹⫹ ⫹⫹⫹ NA
swm1⌬sic1⌬ ⫹⫹⫹ ⫹⫹⫹ ⫹⫹⫹ ⫹/⫺ NA mnd2⌬ubc4⌬ ⫹⫹⫹ ⫹⫹⫹ ⫹⫹⫹ ⫹⫹⫹ ND mnd2⌬CDC28-VF ⫹⫹⫹ ⫹⫹⫹ ⫹⫹⫹ ⫹⫹⫹ ⫹⫹⫹ swm1⌬mad2⌬ ⫹⫹⫹ ⫹⫹⫹ ⫹⫹⫹ ⫹⫹⫹ ⫹⫹⫹
swm1⌬bub2⌬ ⫹⫹⫹ ⫹⫹⫹ ⫹⫹⫹ ⫹⫹⫹ ⫺ mnd2⌬mad2⌬ ⫹⫹⫹ ⫹⫹⫹ ⫹⫹⫹ ⫹⫹⫹ ⫹⫹⫹ mnd2⌬bub2⌬ ⫹⫹⫹ ⫹⫹⫹ ⫹⫹⫹ ⫹⫹⫹ ⫹⫹⫹ swm1⌬clb2⌬ ⫹⫹⫹ ⫹⫹⫹ ⫹⫹⫹ ⫺ NA
swm1⌬clb5⌬ ⫹⫹⫹ ⫹⫹⫹ ⫹⫹⫹ ⫹⫹⫹ NA
SL, synthetic lethality; NA, not applicable. ND, not deter-mined.
SL, synthetic lethality; NA, not applicable.
aSynthetic lethality for both spore germination and vegeta-aSynthetic lethality for both spore germination and
vegeta-tive growth. tive growth.
bDeletion ofMND2does not significantly lower the
nonper-missive temperature of the single mutant.
sive temperatures for budding yeast growth, Swm1p is required for full APC/C function both during the
mi-nificantly defective in Pds1p ubiquitination and binding totic cell cycle and in G1arrest. This result is consistent
of recombinant Cdh1p to purified APC/C complexes with previous reports that swm1⌬ mutants exhibit
de-in vitro. If APC/CCdh1p interactions are significantly
dis-fects in APC/C substrate proteolysis at nonpermissive
rupted byapc9⌬in vivo, we would have expected to see temperatures (Schwickart et al. 2004; Ufano et al.
a much more dramatic stabilization of Pds1p in␣ -factor-2004). In arrest-release experiments conducted at 25⬚,
arrested cells. Our conclusions are supported by the
swm1⌬exhibited partial stabilization of Pds1p-13XMYC,
fact thatapc9⌬mutants arrest normally in␣-factor, in compared to an almost complete failure to degrade
contrast tocdh1⌬mutants, which fail to arrest normally Clb2p-3XHA. Onset of Clb5p-3XHA degradation was
due to high cyclin B/CDK levels (Schwabet al.1997). delayed byⵑ15 min but Clb5p-3XHA levels eventually
One interesting possibility is that the role of Apc9p may decreased to those similar to wild type. The delay in
be primarily restricted to APC/CCdc20p-mediated
proteol-onset of Clb5p-3XHA degradation was not simply due
ysis of the pool of Clb2p whose degradation occurs at to cell cycle delay resulting from failure to degrade
the metaphase-to-anaphase transition, but not the pool Pds1p, asswm1⌬pds1⌬was still delayed in Clb5p-3XHA
that is degraded by APC/CCdh1pupon exit from mitosis.
degradation. Our results indicate that loss of Swm1p
In their study of yeast completely lacking APC/C func-does not affect the degradation of all APC/C substrates.
tion,ThorntonandToczyski(2003) reported that of
apc9⌬ mutants exhibited less severe proteolysis
de-all the essential APC/C subunits,cdc27⌬alone did not fects than didswm1⌬mutants. In␣-factor arrested cells,
require deletion of PDS1for viability, only abrogation where APC/CCdh1pis active, only a small fraction of the
of B-type cyclin activity bySIC1overexpression and dele-initial pool of ectopically expressed Pds1p was stabilized
tion of CLB5. In the context of this result, and given in apc9⌬. In arrest-release experiments, the timing of
our failure to observe significant defects in Pds1p prote-Clb2p-3XHA was delayed 15 min and the kinetics of
olysis inapc9⌬strains, our observation that overexpres-Pds1p-13XMYC and Clb5p-3XHA degradation were
sim-sion of CDC27(but notCDC16 orCDC23) rescues the ilar to those of wild type. Our results suggest that Apc9p
synthetic lethality of apc9⌬ swm1⌬ is intriguing, sug-is required in vivo either for only the full activity of
gesting that, in some APC/C complexes, Apc9p and a small fraction of APC/CCdh1p complexes or against a
Cdc27p may collaborate in the degradation of Clb2p but limited number of APC/C substrates. Alternatively,
dele-not Pds1p. The loss of Cdc27p from APC/C complexes tion of APC9 simply may not have substantial
conse-isolated fromapc9⌬strains supports this notion (Zacha-quences for the physical integrity of the APC/Cin vivo,
riaeet al.1998;Passmoreet al.2003). Deletion ofPDS1, leading to delay in the complete degradation of some
CLB2, or CLB5 alone failed to suppress the synthetic substrates (Clb2p), but not others (Pds1p and Clb5p).
lethality ofapc9⌬swm1⌬(data not shown), and it will be Our data do not support the conclusions ofPassmore
TABLE 4
apc9⌬genetics
Genotype 25⬚ 30⬚ 33⬚ 35⬚ 37⬚ apc9⌬ ⫹⫹⫹ ⫹⫹⫹ ⫹⫹⫹ ⫹⫹⫹ ⫹⫹⫹ apc9⌬swm1⌬ SLa NA NA NA NA
apc9⌬mnd2⌬ ⫹⫹⫹ ⫹⫹⫹ ⫹⫹⫹ ⫹⫹⫹ ND apc9⌬ubc4⌬ ⫹⫹⫹ ⫹⫹⫹ ⫹⫹⫹ ⫹⫹⫹ ND apc9⌬doc1⌬ ⫹⫹⫹ ⫹⫹⫹ ⫺ ⫺ NA
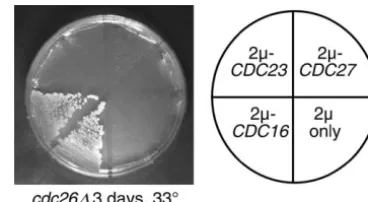
apc9⌬cdc26⌬ ⫹⫹⫹ ⫹⫹⫹ ⫺ ⫺ NA Figure8.—Suppression of APC/C double mutants by over-apc9⌬cdh1⌬ ⫹⫹⫹ ⫹⫹⫹ ⫹⫹⫹ ⫹⫹⫹ ND expression of specific TPR proteins. Overexpression ofCDC16 apc9⌬cdc20-1 SLa,b NA NA NA NA suppresses the temperature sensitivity ofcdc26⌬. cdc26⌬::KanMX
(YVA435) was transformed with 2plasmids expressing a sin-apc9⌬sic1⌬ ⫹⫹⫹ ⫹⫹⫹ ⫹⫹⫹ ⫹⫹⫹ ND
gle APC/C component or regulator or with empty vector only. SL, synthetic lethality; NA, not applicable; ND, not deter- Individual transformants were outgrown on selective media mined. at 25⬚and then applied to selective media at the nonpermissive
aSynthetic lethality for both spore germination and
vegeta-temperature. tive growth.
bSome reversion/papillation was observed.
Cdc27p (Zachariaeet al.1998;Passmoreet al.2003). Schwickartet al.(2004) further report that Swm1p is substrates must be deleted to restore the viability ofapc
required for the efficient binding of Cdc16p, Cdc27p, double-mutant lethalities.
and Apc9p and that incdc23mutants a subcomplex of An emerging model of APC/C subunit function is that
Swm1p, Cdc16p, Cdc27p, and Apc9p fails to bind to the TPR proteins serve as linkers that couple the specificity
other APC/C subunits. They conclude that Swm1p, factors, bound to their substrates, to the APC/C’s catalytic
Cdc26p, Cdc16p, Cdc27p, and Apc9p form a subcom-machinery.Vodermaieret al.(2003) were recently able
plex whose association with the rest of the APC/C is to fractionate human APC/C into subcomplexes, one
Cdc23p dependent. It is important to note that in bud-comprising APC1, APC2, APC4, APC5, and APC11, but
ding yeast all three TPR proteins are encoded by essen-lacking the four TPR proteins and APC10. This
subcom-tial genes. If the function of Swm1p, Apc9p, and Cdc26p plex was competent for the assembly of polyubiquitin
really is to enhance the binding of TPR proteins, then, chains, but not for thein vitroubiquitination of securin.
given the fact thatSWM1, CDC26, andAPC9are nones-Two TPR proteins, APC7 and APC3 (CDC27), strongly
sential genes, it follows that thein vivobinding of TPR/ bound the carboxy termini of CDH1 and CDC20, and
WD-40/substrate complexes to the APC/C core catalytic APC3 was previously shown to bind the carboxy
termi-complex is only partially disrupted in any of the single nus of APC10. CDC20, CDH1, and DOC1 all share a
mutants or thecdc26⌬apc9⌬double mutant. conserved isoleucine arginine motif (Vodermaieret al.
In contrast to our results for Swm1p and Apc9p, we 2003) on their carboxy termini, suggesting that this
did not observe a significant requirement for Mnd2p motif is recognized directly by APC3/CDC27 (Wendt
in APC/C-dependent proteolysis against any of the
sub-et al. 2001; Vodermaier et al. 2003). Consistent with
strates that we examined. In cell cycle experiments, the these data, the phosphorylation of TPR proteins by Clb/
timing of degradation of Pds1p-13XMYC, Clb2p-3XHA, CDK activity has been shown to enhance the binding
and Clb5p-3XHA inmnd2⌬was not substantially differ-of Cdc20p to the complex (RudnerandMurray2000).
ent from that of wild-type strains, and both Pds1p and In the context of this model, the available biochemical
Ase1p-3XHA were rapidly degraded when expressed ec-and genetic data increasingly suggest that Swm1p,
topically from theGAL1 promoter in␣-factor-arrested Apc9p, and Cdc26p collaborate to stabilize the
associa-cells. Our results are consistent with the lack of ubiquiti-tion of TPR/WD-40/substrate complexes to the APC/
nation defects in mnd2⌬ reported by Passmore et al.
C core catalytic machinery.SWM1is a dosage suppressor
(2003). Given the lack of APC/C proteolysis defects in of cdc23-54, and the conditional synthetic lethality of
mnd2⌬, it is difficult to explain the genetic interactions
swm1⌬ ubc4⌬ is suppressed by the overexpression of
ofmnd2⌬mutants that we observe withapc1-21,cdc26⌬,
CDC23and, to a lesser extent,CDC27.swm1⌬is
syntheti-and doc1⌬ and the multicopy suppression of apc-21
cally lethal with bothapc9⌬andcdc26⌬, and these
syn-by 2-MND2 overexpression. Mnd2p clearly associ-thetic lethalities can be rescued by overexpression of
ates with the APC/C in both vegetatively growing cells specific TPR proteins. TAP-APC/C complexes purified
and cells arrested with␣-factor, hydroxyurea, or noco-from strains lacking Cdc26p contain reduced levels of
dozole (Yoon et al. 2002; Hall et al. 2003), but it is Cdc27p, Cdc16p, Cdc23p, and Apc9p (Zachariae et
unclear whether all APC/C complexes contain Mnd2p
al.1998;Schwickart et al.2004), whereas complexes
Immu-TABLE 5 comes deleterious when APC/C catalytic function is compromised by other mutants. Finally, it is possible
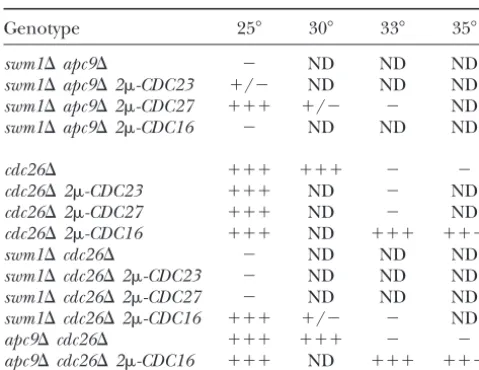
Synthetic lethality ofapc⌬double mutants can be rescued by
that both Mnd2p and Apc1p may have
as-of-yet-undis-overexpression of specific TPR proteins
covered functions unrelated to APC/C ubiquitination activity.
Genotype 25⬚ 30⬚ 33⬚ 35⬚
We thank Doug Koshland, Dave Pellman, Ray Deshaies, Mike Tyers,
swm1⌬apc9⌬ ⫺ ND ND ND
Scott Givan, and Erica Johnson for providing reagents. Thanks go to
swm1⌬apc9⌬2-CDC23 ⫹/⫺ ND ND ND
Isabelle Pot and Daniel Kornitzer for critical reading of the manuscript
swm1⌬apc9⌬2-CDC27 ⫹⫹⫹ ⫹/⫺ ⫺ ND
and to members of the Hieter Lab, Topher Carroll, and Dave Morgan
swm1⌬apc9⌬2-CDC16 ⫺ ND ND ND
for helpful discussions. This work was funded by National Institutes of Health grant CA16569 to P.H.
cdc26⌬ ⫹⫹⫹ ⫹⫹⫹ ⫺ ⫺
cdc26⌬2-CDC23 ⫹⫹⫹ ND ⫺ ND cdc26⌬2-CDC27 ⫹⫹⫹ ND ⫺ ND
cdc26⌬2-CDC16 ⫹⫹⫹ ND ⫹⫹⫹ ⫹⫹⫹ LITERATURE CITED
swm1⌬cdc26⌬ ⫺ ND ND ND
Adames, N. R., J. R. OberleandJ. A. Cooper, 2001 The surveillance
swm1⌬cdc26⌬2-CDC23 ⫺ ND ND ND
mechanism of the spindle position checkpoint in yeast. J. Cell
swm1⌬cdc26⌬2-CDC27 ⫺ ND ND ND Biol.153:159–168.
swm1⌬cdc26⌬2-CDC16 ⫹⫹⫹ ⫹/⫺ ⫺ ND Amon, A., 2001 Together until separin do us part. Nat. Cell Biol.
apc9⌬cdc26⌬ ⫹⫹⫹ ⫹⫹⫹ ⫺ ⫺ 3:E12–E14.
Amon, A., S. IrnigerandK. Nasmyth, 1994 Closing the cell cycle
apc9⌬cdc26⌬2-CDC16 ⫹⫹⫹ ND ⫹⫹⫹ ⫹⫹⫹
circle in yeast: G2 cyclin proteolysis initiated at mitosis persists until the activation of G1 cyclins in the next cycle. Cell77:1037–
ND, not determined.
1050.
Bachmair, A., D. FinleyandA. Varshavsky, 1986 In vivo half-life of a protein is a function of its amino-terminal residue. Science
234:179–186.
Bloecher, A., G. M. VenturiandK. Tatchell, 2000 Anaphase
nodepletion of Mnd2p-13XMYC from glass-bead lysate
spindle position is monitored by the BUB2 checkpoint. Nat. Cell
depleted the majority of Cdc27p from the supernatant
Biol.2:556–558.
but not all of Cdc23p, Cdc16p, or Apc2p, suggesting Burton, J. L., andM. J. Solomon, 2001 D box and KEN box motifs in budding yeast Hsl1p are required for APC-mediated degradation
the possibility that APC/C complexes lack Mnd2p. One
and direct binding to Cdc20p and Cdh1p. Genes Dev.15:2381–
formal possibility is that, in otherwise wild-type mitotic
2395.
cells, the functional contribution of Mnd2p to overall Charles, J. F., S. L. Jaspersen, R. L. Tinker-Kulberg, L. Hwang, A. Szidonet al., 1998 The Polo-related kinase Cdc5 activates
APC/C activity is so minimal as to be undetectable
and is destroyed by the mitotic cyclin destruction machinery in
within the limitations of our assays, but lack of Mnd2p
S. cerevisiae. Curr. Biol.8:497–507.
exacerbates APC/C defects when the complex is further Ciechanover, A., 1994 The ubiquitin-proteasome proteolytic
path-way. Cell79:13–21.
perturbed. This hypothesis is supported by the limited
Cohen-Fix, O., 2001 The making and breaking of sister chromatid
amount of existing data about Mnd2p and its
Schizosac-cohesion. Cell106:137–140.
charomyces pombe homolog Apc15p. Growth curve and Cohen-Fix, O., J. M. Peters, M. W. KirschnerandD. Koshland,
1996 Anaphase initiation in Saccharomyces cerevisiae is
con-flow cytometry experiments revealed thatmnd2⌬stains
trolled by the APC-dependent degradation of the anaphase
inhib-grow slightly slower than wild type, but even when inhib-grown
itor Pds1p. Genes Dev.10:3081–3093.
at 37⬚there is little increase in the 2N DNA content in Cross, F. R., 2003 Two redundant oscillatory mechanisms in the
yeast cell cycle. Dev. Cell4:741–752.
mnd2⌬ (Hall et al. 2003; this work). Deletion of S.
Deshaies, R. J., V. ChauandM. Kirschner, 1995 Ubiquitination
pombeApc15p is viable, but unlike budding yeastmnd2⌬,
of the G1 cyclin Cln2p by a Cdc34p-dependent pathway. EMBO
apc15⌬/apc15⌬mutants did not exhibit meiotic defects J.14:303–312.
Donovan, J. D., J. H. Toyn, A. L. JohnsonandL. H. Johnston, 1994
(Yoonet al.2002). Consistent with our findings,apc15⌬
P40SDB25, a putative CDK inhibitor, has a role in the M/G1
is synthetically lethal withcut4-553, a
temperature-sensi-transition in Saccharomyces cerevisiae. Genes Dev.8:1640–1653.
tive mutant in theS. pombeApc1p homolog (Yoonet al. Fang, G., H. Yu and M. W. Kirschner, 1998 Direct binding of CDC20 protein family members activates the
anaphase-promot-2002). Another hypothesis is that Mnd2p may play a
ing complex in mitosis and G1. Mol. Cell2:163–171.
limited role in promoting the initial assembly of APC/
Gietz, R. D., R. H. Schiestl, A. R. WillemsandR. A. Woods, 1995
C complexes or in maintaining the stability or proper Studies on the transformation of intact yeast cells by the LiAc/
SS-DNA/PEG procedure. Yeast11:355–360.
cellular localization of one or more APC/C subunits, a
Gmachl, M., C. Gieffers, A. V. Podtelejnikov, M. MannandJ. M. function that might be dispensable once the APC/C is
Peters, 2000 The RING-H2 finger protein APC11 and the E2
fully assembled and activated. However, levels of Apc1p enzyme UBC4 are sufficient to ubiquitinate substrates of the
anaphase-promoting complex. Proc. Natl. Acad. Sci. USA97:
inmnd2⌬are not substantially different from those in
8973–8978.
wild-type cells, indicating that Mnd2p does not regulate
Goh, P. Y., H. H. LimandU. Surana, 2000 Cdc20 protein contains
overall Apc1p levels (V.Aneliunas, personal communi- a destruction-box but, unlike Clb2, its proteolysis is not acutely
dependent on the activity of anaphase-promoting complex. Eur.
cation). In addition, Mnd2p may be required for the
J. Biochem.267:434–449.
turnover of a limited subset of APC/C substrates whose
Gorbsky, G. J., 2001 The mitotic spindle checkpoint. Curr. Biol.
proteolysis is not required for cell cycle progression 11:R1001–R1004.
Gordon, D. M., andD. M. Roof, 2001 Degradation of the kinesin