Copyright © 2003, American Society for Microbiology. All Rights Reserved.
Molecular Typing of
Salmonella enterica
Serovar Typhi Isolates
from Various Countries in Asia by a Multiplex PCR Assay on
Variable-Number Tandem Repeats
Yichun Liu,
1May-Ann Lee,
1Eng-Eong Ooi,
2Yeo Mavis,
3Ai-Ling Tan,
3and Hung-Hiang Quek
1*
Biomedical Science Laboratory, Defence Medical Research Institute, Defence Science and Technology Agency,1
Environmental Health Institute, National Environment Agency,2and Department of Pathology,
Singapore General Hospital,3Singapore
Received 8 June 2003/Returned for modification 23 April 2003/Accepted 9 June 2003
A multiplex PCR method incorporating primers flanking three variable-number tandem repeat (VNTR) loci
(arbitrarily labeled TR1, TR2, and TR3) in the CT18 strain of Salmonella enterica serovar Typhi has been
developed for molecular typing of S. entericaserovar Typhi clinical isolates from several Asian countries,
including Singapore, Indonesia, India, Bangladesh, Malaysia, and Nepal. We have demonstrated that the multiplex PCR could be performed on crude cell lysates and that the VNTR banding profiles produced could be easily analyzed by visual inspection after conventional agarose gel electrophoresis. The assay was highly discriminative in identifying 49 distinct VNTR profiles among 59 individual isolates. A high level of VNTR profile heterogeneity was observed in isolates from within the same country and among countries. These VNTR profiles remained stable after the strains were passaged extensively under routine laboratory culture
condi-tions. In contrast to theS. entericaserovar Typhi isolates, an absence of TR3 amplicons and a lack of length
polymorphisms in TR1 and TR2 amplicons were observed for otherS. entericaserovars, such asSalmonella
entericaserovar Typhimurium,Salmonella entericaserovar Enteritidis, andSalmonella entericaserovar Para-typhi A, B, and C. DNA sequencing of the amplified VNTR regions substantiated these results, suggesting the high stability of the multiplex PCR assay. The multiplex-PCR-based VNTR profiling developed in this study provides a simple, rapid, reproducible, and high-resolution molecular tool for the epidemiological analysis of
S. entericaserovar Typhi strains.
Salmonella entericaserovar Typhi is the etiological agent of typhoid fever, of which there are an estimated 16 million cases with 600,000 related deaths worldwide (15). The gram-negative rod-shaped bacterium is pathogenic only in humans, where it can be cultured from blood and stools. Infection occurs when water or food contaminated with S. entericaserovar Typhi is consumed. Most patients who recover from the infection are able to eliminate the bacterium completely from their bodies. However, some of them may remain as healthy carriers, con-tinuously sheddingS. entericaserovar Typhi in their stools (5). Epidemiological studies of pathogens are of great impor-tance in controlling their dissemination. The capability to strain type pathogens is a critical tool in epidemiological in-vestigations. A number of strain-typing methods have been developed forS. entericaserovar Typhi. The classical methods of phage typing and isoenzyme analysis have been increasingly complemented by molecular techniques, such as pulsed-field gel electrophoresis (PFGE) (37, 38), ribotyping (2, 25), ran-dom amplification of polymorphic DNA (RAPD) (32), and DNA fingerprinting using the mobile genetic element IS200 (39), as well as the recent development of amplified fragment length polymorphism (AFLP) for discriminating S. enterica serovar Typhi strains (22).
Despite the availability of the above-mentioned methods,
there is still a lack of methods with the right combination of rapidity, ease of use, reproducibility, and discriminatory power for typing S. enterica serovar Typhi. PFGE, ribotyping, and AFLP procedures are technically complicated and time-con-suming. RAPD is rapid but is hampered by its poor reproduc-ibility. IS200fingerprinting has been reported to lack discrim-inative power (40).
Variable-number tandem repeats (VNTR) have been in-creasingly used as molecular markers for strain typing of var-ious bacteria. VNTR, or short sequence repeats, consist of unique DNA elements that are repeated in tandem (41). In-dividual strains within a bacterial species often maintain the same sequence element but with different copy numbers. Such variation is often caused by slipped-strand mispairing during DNA replication (6, 43). Since sequence homology exists be-tween strains in the flanking region of the VNTR locus, PCR amplification with flanking-sequence-specific primers can be used to determine the variations in copy numbers of repeat units that reflect the intraspecies genetic diversity. Therefore, individual strains could be easily identified by the amplicon sizes. This forms the basis for using VNTR for strain typing.
The availability of complete microbial genomic sequences has greatly facilitated the identification of VNTR for strain typing. Using software programs that rapidly scan the genomes for them, VNTR can be quickly located, and primers for PCR analysis can made based on the flanking sequences. We adopted this method of VNTR detection in this study, which resulted in the development of a multiplex PCR assay using three different VNTR loci as molecular markers to differenti-* Corresponding author. Mailing address: Biomedical Science
Lab-oratory, Defence Medical Research Institute, 10 Medical Dr. No. 02-14, Singapore 117597, Singapore. Phone: (65) 97380437. Fax: (65) 67791677. E-mail: [email protected].
4388
on May 15, 2020 by guest
http://jcm.asm.org/
ate clinicalS. entericaserovar Typhi isolates from several Asian countries. To our knowledge, this is the first report describing the use of multiplex-PCR-based VNTR profiling for the strain typing ofS. entericaserovar Typhi isolates.
MATERIALS AND METHODS
Bacterial isolates and culture conditions.TheS. entericaserovar Typhi isolates used in this study are described in Table 1. A total of 61S. entericaserovar Typhi isolates collected during the years 2000 and 2001 were obtained from the Sin-gapore General Hospital. Four pairs of the 61 isolates were collected from four different patients (i.e., one pair per patient). Three of the four pairs were collected at different times over the course of infection, while one pair was collected from fresh whole blood versus a blood clot on the same day. Therefore, the total number of individual clinical isolates was 57 (Table 1). Of these 57 isolates, 13 were indigenous isolates obtained from sporadic cases in Singapore, while the rest were isolated from imported cases from several Asian countries, including Indonesia, India, Bangladesh, Malaysia, and Nepal. In addition,S. entericaserovar Typhi strains CT18 and TY2-b were obtained from the Salmo-nellaGenetic Stock Centre (SGSC), University of Calgary, Calgary, Canada, to be used as reference strains in the study.
Phage typing of the clinical isolates was carried out by the hospital, but some isolates did not produce conclusive results (Table 2).
Strains of otherSalmonella entericaserovars were used as controls in the study. They includedS. entericaserovar Typhimurium (strains TR7095 and SL1344),S. entericaserovar Enteritidis (strains LK5 and PT4) obtained from the SGSC, and
S. entericaserovar Paratyphi A, B, and C from the Singapore General Hospital.
Preparation of crude bacterial DNA.About one to five bacterial colonies from Luria-Bertani agar medium were suspended in 200l of distilled H2O. The cell suspension was boiled at 95°C for 7 min and then stored at 4°C before being used directly for PCR.
Selection of VNTR loci and PCR primers.Five VNTR loci, designated TR1 to TR5, were selected by the Tandem Repeat Finder program (http://tandem .biomath.mssm.edu/trf/trf.html) (4) from the entire genomic sequence ofS. en-tericaserovar Typhi CT18, a multiple-drug-resistant strain (26), under GenBank accession no. NC_003198. PCR primers were designed from sequences flanking the respective TR1 to TR5 loci (Table 3). These five loci, each containing a unique direct repeat, are located within intergenic regions, increasing the prob-ability of copy number variation.
Screening for length polymorphism via PCR of individual VNTR loci.Prior to performing the multiplex PCR assay, the five VNTR loci were evaluated for the presence of allelic variations using PCR with the respective primer pairs on 20 differentS. entericaserovar Typhi isolates. All PCR assays were performed with the Platinum PCR Supermix kit (Invitrogen Life Technologies). Each 25-l reaction mixture contained a mixture of 1l of the suspension of the boiled bacterial cell lysate, 5 pmol of each primer, and 22.5l of the Supermix supplied
with the kit. After an initial denaturation at 94°C for 2 min, the reaction was performed for 35 cycles at 94°C for 30 s, 55°C for 30 s, and 72°C for 1 min, followed by an extension step of 72°C for 7 min. In these 20 isolates, allelic variations were eventually detected in the TR1, TR2, and TR3 loci but not the TR4 and TR5 loci (see Results). Therefore, the TR1, TR2, and TR3 loci were further characterized in this study.
Multiplex PCR assay.The primers flanking the TR1, TR2, and TR3 loci were selected for developing the multiplex PCR assay. Each 25-l reaction mixture contained 1l of the bacterial lysate suspension and 10 pmol each of the forward and reverse primers for TR1 and TR3, as well as 12.5 pmol each of the corre-TABLE 1. 59S. entericaserovar Typhi isolates used in this study
Origin Reference no. of isolatesa Yr of collection
Singapore 349, 578, 581 2001
156, 320, 326, 489, 493, 517, 541,
557, 587, 612 2000
Indonesia 12, 31, 77, 117, 158, 182, 208, 219, 226, 256, 293, 303, 311, 343, 377, 425, 426, 447
2001
76, 191, 288, 308, 463, 609 2000
India 10, 120, 138 2001
33, 72, 234, 261, 306, 354, 410 2000
Bangladesh 283, 315, 446, 508 2001
136, 549 2000
Malaysia 396 2001
470 2000
Nepal 365, 367 2001
SGSC CT18, TY2-b 2001
[image:2.603.43.283.80.246.2]aPaired isolates were obtained for isolates 493, 256, and 136. Each pair was collected from the corresponding patients 17, 35, and 38 days apart, respectively. A paired isolate was also obtained for 306 from fresh whole blood versus a blood clot of a patient on the same day.
TABLE 2. VNTR profiles and phage types of 59S. entericaserovar Typhi isolates
VNTR profile
(TR1/TR2/TR3)a reference no.Isolate
VNTR profile designation
Phage typeb
5x/36.9x/2.3x 587 S1 E1
8x/35.9x/72.3x 156 S2 NC
10x/16.9x/2.3x 557, 612 S3 D2
10x/23.9x/2.3x 320, 326 S4 D2
12x/16.9x/2.3x 578 S5 NC
13x/6.9x/2.3x 581 S6 NC
13x/32.9x/3.3x 541 S7 A1
15x/20.9x/2.3x 349 S8 D1
16x/12.9x/2.3x 517 S9 NC
19x/10.9x/§2.3x 493 S10 NC
19x/15.9x/2.3x 489 S11 NC
6x/27.9x/2.3x 77 IDN1 NC
9x/11.9x/2.3x 303 IDN2 K1
10x/11.9x/§2.3x 226, 377 IDN3 A
10x/30.9x/␦2.3x 256 IDN4 NC
11x/15.9x/2.3x 12 IDN5 D2
12x/6.9x/2.3x 31, 426, 470 IDN6/MS2 D2/NC
12x/11.9x/2.3x 343 IDN7 D2
12x/17.9x/2.3x 208, 219, 293 IDN8 D2
12x/19.9x/2.3x 609 IDN9 D2
12x/20.9x/2.3x 182 IDN10 NC
12x/26.9x/2.3x 117 IDN11 NC
12x/32.9x/3.3x 447 IDN12 M1
13x/4.9x/2.3x 158 IDN13 NC
13x/12.9x/2.3x 288 IDN14 D2
14x/14.9x/§2.3x 191 IDN15 NC
14x/21.9x/2.3x 311 IDN16 NC
14x/22.9x/§2.3x 308 IDN17 UVS1
14x/24.9x/2.3x 425 IDN18 D2
15x/5.9x/2.3x 463 IDN19 NC
16x/20.9x/2.3x 76 IDN20 NC
5x/29.9x/2.3x 234, 261, 354 IN1 E1
5x/31.9x/2.3x 10 IN2 E1
5x/32.9x/2.3x 120 IN3 E1
6x/22.9x/1.3x 72 IN4 J1
10x/15.9x/2.3x 33 IN5 E1
12x/27.9x/2.3x 138 IN6 NC
14x/9.9x/4.3x 410 IN7 A1
14x/14.9x/2.3x 306 IN8 NC
5x/33.9x/2.3x 136 B1 NC
8x/8.9x/2.3x 549 B2 NC
8x/24.9x/2.3x 283 B3 NC
9x/18.9x/2.3x 315 B4 NC
9x/27.9x/2.3x 446 B5 NC
18x/13.9x/2.3x 508 B6 NC
11x/8.9x/3.3x 396 MS1 A
10x/10.9x/3.3x 365, 367 N1 M1
11x/9.9x/2.3x TY2-b TY2-b type 12x/27.9x/§3.3x CT18 CT18 type
aThe VNTR profiles were defined based on the copy numbers of the respec-tive repeats at the three different loci in each isolate.xrepresents the copy number of each VNTR;␦represents a 122-bp deletion; § represents a 61-bp deletion in the 5⬘upstream sequences of the TR3 alleles.
bNC, not confirmed.
on May 15, 2020 by guest
http://jcm.asm.org/
[image:2.603.306.540.89.570.2]sponding primers for TR2, in addition to 22.5l of the Supermix. The same PCR cycling conditions described above were used. A 5-l aliquot of the PCR prod-ucts was resolved on a 1.5% (wt/vol) agarose gel containing 0.5g of ethidium bromide ml⫺1in 0.5⫻TBE buffer (45 mM Tris-borate, 1 mM EDTA, pH 8.0). A 100-bp DNA marker (Promega Corp., Madison, Wis.) was used as a molecular size standard. The multiplex PCR was repeated more than five times using different thermocyclers and different preparations of the cell lysates to ascertain the reproducibility of the results. Finally, the reproducibility, discriminatory power, and typeability of this multiplex-PCR-based VNTR typing method, in comparison with other PCR-based typing methods, including RAPD, enterobac-terial repetitive intergenic consensus PCR, and AFLP, were evaluated on 20 randomly selectedS. entericaserovar Typhi isolates.
DNA sequencing and data analysis.To confirm the identities of the amplicons and that any length polymorphism was due to variations in the VNTR copy number, DNA sequencing was performed on the nonmultiplexed PCR products from all 61 clinical isolates and the 2 reference strains following a purification process primarily by an enzyme digest method. An alternative method using the QIAquick PCR purification kit (Qiagen, Chatsworth, Calif.) was used only for PCRs with lower yields. The enzyme digest method involved treating 8l of PCR products with 0.3 U of exonuclease I and 1.5 U of shrimp alkaline phosphatase (Amersham) in a 10-l reaction mixture at 37°C for 5 min to remove the excess primers and deoxynucleoside triphosphates, followed by the inactivation of the enzymes at 72°C for 15 min. A 2-l aliquot of the enzyme-treated PCR products was used directly in the sequencing reaction. Each PCR fragment was sequenced from both directions using the same PCR primers with a DYEnamic ET dye terminator kit (Amersham Pharmacia Biotech) according to the manufacturer’s instructions. The sequencing reactions were resolved on a MegaBACE auto-mated capillary sequencer. DNA sequences were analyzed with the LASER-GENE software package (DNASTAR, Madison, Wis.) and Tandem Repeat Finder (4).
RESULTS
Screening for allelic variations at the TR1, TR2, and TR3
VNTR loci.PCR was performed on 20 isolates using individual
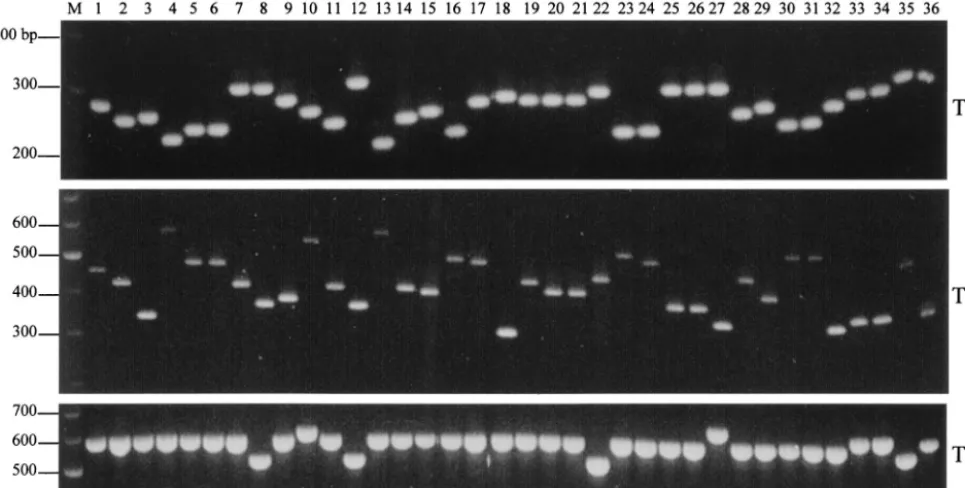
primer pairs to evaluate the potential use of the TR1 to TR5 VNTR loci as molecular markers for the strain typing of S. entericaserovar Typhi.Length polymorphisms of the amplified fragments among the 20 isolates were observed for the TR1, TR2, and TR3 loci but not for the TR4 and TR5 loci, for which amplicons of ⬃420 and 190 bp, respectively, were observed (results not shown). The specificity of the primers could be seen from the single band obtained in each reaction. Length polymorphisms were especially pronounced for TR1 and TR2. Allelic variations at these three loci were also observed when the remaining isolates were screened. The results of the am-plification of representative isolates can be seen in Fig. 1.
It was also observed that there were no changes in the sizes of the TR1, TR2, and TR3 amplicons from four isolates that had been passaged for 40 rounds (results not shown), suggest-ing that the three loci have sufficient genetic stability for use as molecular strain-typing markers. In addition, no changes in the
sizes and sequences of TR1, TR2, and TR3 were observed in the paired S. enterica serovar Typhi isolates, especially the three pairs that were collected from the same patients at dif-ferent times (Fig. 1, lanes 8 and 12 and 30 and 31) over the course of infection. This implied that the VNTR markers in the same strain did not change over the course of an infection. This is a strong indication that these markers could cluster epide-miologically related strains instead of erroneously segregating them.
Sequence analysis of individual VNTR alleles.The
specific-ity of the PCR was confirmed by the sequencing data. The TR1, TR2, and TR3 sequences of the CT18 strain were iden-tical to those from GenBank. Sequence analysis of all ampli-cons from hospital and reference strains revealed that the length polymorphisms observed among the TR1 and TR2 am-plicons were indeed the result of different copy numbers of the respective repeats. As expected, paired isolates taken from the same patient had identical repetitive sequences despite the different times of collection or types of clinical specimens. Among the 59 individual isolates, including two reference strains, a total of 14 distinct alleles ranging between 2 and 19 copies of the 7-bp repeat unit were observed at the TR1 locus, while a total of 29 distinct alleles ranging between 4.9 and 36.9 copies of the 8-bp repeat were detected at the TR2 locus. In contrast to TR1 and TR2, there were only four alleles of 1.3, 2.3, 3.3, and 4.3 copies of the 26-bp repeat at the TR3 locus among the 59 isolates (Table 2).
Sequencing analysis of the TR3 amplicons also revealed deletions of 61- and 122-bp nucleotides in the 5⬘ upstream region of the TR3 repeats among seven different isolates (re-sults not shown). The 61-bp deletion was found in six isolates (493, 226, 377, 191, 308, and CT18), while the 122-bp deletion was found only in the Indonesian isolate 256 (Table 2). There-fore, unlike the TR1 and TR2 amplicons, the length polymor-phisms for the TR3 amplicons among these seven isolates is a result of variations in the copy number of the TR3 VNTR, as well as the presence of the nucleotide deletions in the 5⬘
upstream regions.
Multiplex-PCR-based VNTR analysis.Amplicons of
[image:3.603.44.540.80.193.2]differ-ent sizes were observed for isolates when they were subjected to multiplex PCR with primers for TR1, TR2, and TR3 (Fig. 2). Based on visual observation, each of these bands was sim-ilar in size to its counterpart obtained during amplification with individual primer pairs (Fig. 1). A total of 49 arbitrarily as-signed strain types based on the banding profiles could be identified among the 59 isolates (Table 2). The results showed TABLE 3. PCR primers flanking the five VNTR loci selected from theS. entericaserovar Typhi CT18 genome
Primer Nucleotide positions Sequence (5⬘-3⬘) VNTR sequence
TR1F1 2017115–2017136 AGA ACC AGC AAT GCG CCA ACG A (AGAAGAA)12
TR1R1 2017354–2017375 CAA GAA GTG CGC ATA CTA CAC C
TR2F1 2556810–2556831 CCC TGT TTT TCG TGC TGA TAC G (CCAGTTCC)27
TR2R2 2557299–2557320 CAG AGG ATA TCG CAA CAA TCG G
TR3F1 2926145–2926166 CGA AGG CGG AAA AAA CGT CCT G (CGCGGGGATCGGTTTATCCCCGCTGG)3.3
TR3R1 2926668–2926689 TGC GAT TGG TGT CGT TTC TAC C
TR4F2 4396728–4396749 AAA AGC CCG TCT AGT CTT GCA G (GAAATAAAAAATG)2.1
TR4R1 4397127–4397148 ATC CTT CGG TAT CGG GGT ATC C
TR5F1 4624169–4624190 TGA AAA CCG GCT CGT AGC AGT G (CGTCACG)4.7
TR5R1 4624342–4624363 CAT ACG GTT ACT GCG GGA TTG G
on May 15, 2020 by guest
http://jcm.asm.org/
that we were able to segregate 13 Singapore isolates into 11 VNTR types (S1 to S11), 24 Indonesian isolates into 20 VNTR types (IND1 to IND20), 10 Indian isolates into 8 VNTR types (IN1 to IN8), 6 Bangladeshi isolates into 6 VNTR types (B1 to B6), and 2 Malaysian isolates into 2 VNTR types (MS1 and MS2). Two Nepalese isolates were found to have identical VNTR types (N1). With the excep-tion of the Malaysian isolate 470 (VNTR profile MS2), which is identical in profile to the Indonesian isolates 31 and 426 (VNTR profile IDN6), most of the isolates from each country were distinctive and different from the isolates from all other countries. Intracountry variation in the VNTR profiles was generally observed, although there were some
[image:4.603.54.537.75.319.2]isolates that had the same profile. The reference strains CT18 and TY2-b exhibited distinct banding profiles com-pared with one another and with the rest of the isolates (Table 2 and Fig. 2, lanes 35 and 36). Repetition of the multiplex PCR confirmed that the results were reproducible. Comparison of the VNTR profiles with the phage-typing data revealed that isolates with the same VNTR profile share the same phage type. However, isolates with the same phage types could exhibit different VNTR profiles (Table 2). Finally, a comparison between the multiplex PCR assay and other PCR-based strain-typing methods, such as RAPD, en-terobacterial repetitive intergenic consensus PCR, and AFLP, demonstrated that VNTR profiling had the best mix of dis-FIG. 1. Gel electrophoresis of the TR1, TR2, and TR3 VNTR loci of 36 representative isolates amplified using the TR1, TR2, and TR3 primer pairs. Lanes: 1 to 14, Singapore isolates (lanes 8 and 12 are paired isolates collected 17 days apart); 15 to 22, Indonesian isolates; 23 to 27, Indian isolates; 28 to 32, Bangladeshi isolates (lanes 30 and 31 are paired isolates collected 38 days apart); 33 and 34, Nepalese isolates; 35 and 36,S. entericaserovar Typhi CT18 and TY2-b; M, 100-bp DNA marker.
FIG. 2. Agarose gel analysis of the VNTR banding profiles amplified from the 36 representativeS. entericaserovar Typhi isolates by multiplex PCR containing TR1, TR2, and TR3 primer pairs. Lanes: 1 to 14, Singapore isolates (lanes 8 and 12 are paired isolates collected 17 days apart); 15 to 22, Indonesian isolates; 23 to 27, Indian isolates; 28 to 32, Bangladeshi isolates (lanes 30 and 31 are paired isolates collected 38 days apart); 33 and 34, Nepalese isolates; 35 and 36,S. entericaserovar Typhi CT18 and TY2-b; M, 100-bp DNA marker.
on May 15, 2020 by guest
http://jcm.asm.org/
[image:4.603.48.541.557.686.2]criminatory power, reproducibility, typeability, speed, ease of usage, and cost (data not shown).
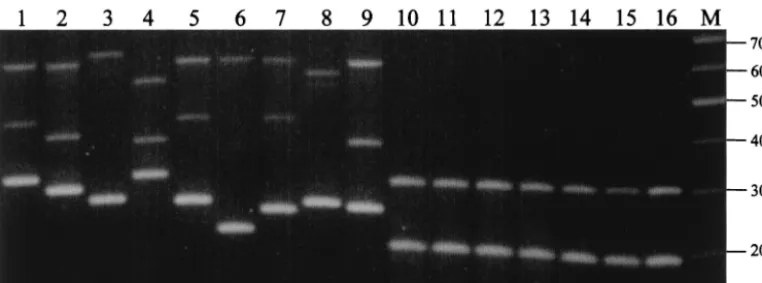
Specificity of the multiplex PCR assay.When the multiplex
PCR was performed on S. entericaserovar Typhimurium, S. entericaserovar Enteritidis, andS. entericaserovar Paratyphi A, B, and C, two amplicons of⬃200 and 300 bp were observed for all reactions (Fig. 3). This is in sharp contrast to the highly polymorphic VNTR profiles in theS. enterica serovar Typhi isolates. A PCR assay using individual primer pairs confirmed that the 200-bp fragment was amplified by the TR1 primers while the 300-bp fragment was a product of the TR2 primers. However, no amplification was observed using the TR3 prim-ers (data not shown), suggesting that sequences of the TR3 primer sites are conserved inS. entericaserovar Typhi.
DISCUSSION
In this study, we characterized three VNTR loci from S. entericaserovar Typhi strain CT18 and used them as molecular markers to discriminate among isolates from Singapore, Indo-nesia, India, Bangladesh, Malaysia, and Nepal. A multiplex PCR assay containing primers specific to the flanking se-quences of these VNTR loci was developed. Our results have shown that this method is rapid, reproducible, and highly dis-criminative for the strain typing of S. entericaserovar Typhi isolates. The banding profiles could be easily interpreted by visual inspection after electrophoresis on conventional agarose gels.
Our study showed that substantial genetic heterogeneity at the VNTR loci exists amongS. entericaserovar Typhi isolates within and among geographical areas. Previous studies using phenotypic markers, such as multilocus enzyme electrophore-sis, had suggested that S. entericaserovar Typhi strains from different geographical regions were from two different clones (31). However, more recent studies using genotypic methods uncovered more variability than was previously observed (24, 35). These studies suggested that multipleS. entericaserovar Typhi clones exist within and among countries. One of the studies, of 75 multidrug-resistantS. entericaserovar Typhi iso-lates from four independent outbreaks of typhoid fever in southern Vietnam between 1993 and 1997, reported that each
of these outbreaks was caused by a single bacterial strain (8). However, different outbreaks did not derive from the clonal expansion of a singleS. entericaserovar Typhi strain, leading to the conclusion that different clones were in circulation. Franco et al. (13) also found genetic heterogeneity in 69 Indonesian and Peruvian isolates by using envelope protein profiles and chromosomal restriction endonuclease digestion patterns. A study of Chilean strains isolated between 1977 and 1986 re-vealed multipleS. entericaserovar Typhi ribotypes, suggesting that there were multiple sources of infection due to different strains being in circulation (12). Work on 73 Vietnamese and 217 Hong Kong strains using plasmid profile analysis, plasmid fingerprinting, ribotyping, and total-DNA fingerprinting re-vealed a high level of genetic heterogeneity within each coun-try, as well as among the countries (21). A Malaysian study using PFGE on 158S. entericaserovar Typhi isolates revealed that individual outbreaks were associated with closely related PFGE strains, whereas isolates from sporadic cases were very diverse (37). A more comprehensive PFGE analysis of 120S. entericaserovar Typhi isolates from Southeast Asia (60 from Malaysia, 50 from Indonesia, and 10 from Thailand) also found considerable genetic diversity among them, although some PFGE profiles were shared between countries (38). Nair et al. (23) reported that many PFGE types are circulating in Asia. Our findings are therefore in concordance with these reports. Given the high volume of human traffic among Singapore, Indonesia, and Malaysia, we would have expected more ge-netic homogeneity among isolates. This was not observed. Nonetheless, the probability of observing identical VNTR pro-files in different countries is also dependent on the locations where the isolates were collected. It is not unreasonable to postulate that locations with a higher frequency of cross-bor-der interactions (e.g., towns and cities where there are more foreign workers and visitors) are more likely to have isolates with identical strain profiles. Moreover, the possibility that increased sample size would reveal identical VNTR profiles among isolates of different countries could not be ruled out.
[image:5.603.102.483.71.212.2]The globalization of international trade and travel has changed the epidemiological profiles of many diseases by mak-ing it easier for pathogens to cross geographical borders. Based FIG. 3. Specificity of the multiplex PCR assay for strain typingS. entericaserovar Typhi isolates. Lanes: 1 to 9, variousS. entericaserovar Typhi isolates; 10,S. entericaserovar Typhimurium TR7095; 11,S. entericaserovar Typhimurium SL1344; 12,S. entericaserovar Enteritidis LK5; 13,S. entericaserovar Enteritidis PT4; 14,S. entericaserovar Paratyphi A; 15,S. entericaserovar Paratyphi B; 16,S. entericaserovar Paratyphi C; M, 100-bp DNA marker.
on May 15, 2020 by guest
http://jcm.asm.org/
upon a surveillance report by the Ministry of the Environment (http://www.env.gov.sg), 15 to 20% of the typhoid cases in Singapore from the years 1996 to 2000 were indigenous cases. These indigenousS. entericaserovar Typhi isolates had VNTR profiles that were distinctive and different from those of the other countries studied. This suggests that there is a cluster of strains unique to and circulating in Singapore. Whether the strains responsible for the sporadic cases were identical to those strains harbored by healthy S. enterica serovar Typhi carriers remains to be investigated.
In our study, we sequenced the TR1, TR2, and TR3 ampli-cons to verify PCR specificity and to confirm that any length polymorphism observed was due solely to variations in VNTR copy numbers. This was necessary, since these loci were being analyzed for the first time. Our results confirmed that the sequencing of the TR1 and TR2 amplicons would not be nec-essary for subsequent strain-typing attempts. Sequencing of TR3 might be needed, since the length polymorphism of the TR3 amplicons could be the result of deletions at the 5⬘ flank-ing regions. However, given the high copy number variability among isolates at the TR1 and TR2 loci, the sequencing of TR3 would be quite an infrequent event, since length polymor-phisms at TR1 and TR2 are varied enough for strain separa-tion.
We have yet to ascertain if the VNTR polymorphisms seen in the TR1, TR2, and TR3 loci, as well as the deletions in the TR3 upstream region, have any bearing on microbial physiol-ogy and virulence. All three VNTR loci are found in noncoding regions⬃200 to 700 bp upstream of hypothetical open reading frames. TR1 is adjacent to an open reading frame that codes for a hypothetical protein with hydrophobic membrane-span-ning regions that is highly homologous to theEscherichia coli YedE putative permease protein. VNTR copy number varia-tions in the promoter region or within the coding regions of virulence genes have been documented to have an effect on a pathogen’s ability to adapt to a hostile host environment (14, 28, 29, 30, 41).
To our knowledge, PCR-based VNTR profiling has not been reported for studying the relationship between theS. enterica serovar Typhi isolates, although it has been used for the strain typing of a number of bacterial species, e.g.,Mycoplasma pneu-moniae(7),Staphylococcus aureus(10),Bacillus anthracis(3), Mycobacterium tuberculosis (16), Neisseria meningitidis (27), group B streptococci (19), andYersinia pestis(1). This highly versatile method can be further improved by using primers tagged with fluorescent dyes, allowing accurate sizing of am-plicons after electrophoresis on an automated DNA se-quencer. New primers can also be added to increase the dis-criminative power. More recently, multiple-locus VNTR analysis, a scaled up version of VNTR amplification, has been used to increase discriminative power for differentiating among isolates from genetically highly homogenous bacterial species, such asY. pestis(18),B. anthracis(17) andFrancisella tularensis(11).
The high degree of reproducibility and discriminative power of our multiplex PCR method suggests that it could be used for both comparative typing and library typing. Comparative typ-ing methods are used for outbreak investigations with the aim of segregating strains (34). These methods must therefore be highly typeable and discriminative. Library typing methods are
used for epidemiological surveillance and hence must be highly reproducible within and between laboratories for an extended period. These techniques should also have some standard type nomenclature for easy comparison of results. Our VNTR-based multiplex PCR fulfils these requirements by giving fast, discriminative, and reproducible results with minimal reagent and manpower costs.
ACKNOWLEDGMENT
This work was supported by a grant from the Defense Research Committee, Ministry of Defence, Singapore.
REFERENCES
1. Adair, D. M., P. L. Worsham, K. K. Hill, A. M. Klevytska, P. J. Jackson, A. M. Friedlander, and P. Keim.2000. Diversity in a variable-number tan-dem repeat fromYersinia pestis.J. Clin. Microbiol.38:1516–1519. 2. Altwegg, M., F. W. Hickman-Brenner, and J. J. Farmer.1989. Ribosomal
RNA gene restriction patterns provide increased sensitivity for typing Sal-monella typhistrains. J. Infect. Dis.160:145–149.
3. Andersen, G. L., J. M. Simchock, and K. H. Wilson.1996. Identification of a region of genetic variability amongBacillus anthracisstrains and related species. J. Bacteriol.178:377–384.
4. Benson, G.1999. Tandem repeats finder: a program to analyze DNA se-quences. Nucleic Acids Res.27:573–580.
5. Buchwald, D. S., and M. J. Blaser.1984. A review of human salmonellosis. II. Duration of excretion following infection with non-typhiSalmonella.Rev. Infect. Dis.6:345–356.
6. Bzymek, M., and S. T. Lovett.2001. Instability of repetitive DNA sequences: the role of replication in multiple mechanisms. Proc. Natl. Acad. Sci. USA
98:8319–8325.
7. Colman, S. D., P. C. Hu, and K. F. Bott.1990. Prevalence of novel repeat sequences in and around the P1 operon in the genome ofMycoplasma pneumoniae.Gene87:91–96.
8. Connerton, P., J. Wain, T. T. Hien, T. Ali, C. Parry, N. T. Chinh, H. Vinh, V. A. Ho, T. S. Diep, N. P. Day, N. J. White, G. Dougan, and J. J. Farrar.
2000. Epidemic typhoid in Vietnam: molecular typing of multiple-antibiotic-resistantSalmonella entericaserotype typhi from four outbreaks. J. Clin. Microbiol.38:895–897.
9. Dalla-Costa, L. M., K. Irino, J. Rodrigues, I. N. Rivera, and L. R. Trabulsi.
1998. Characterisation of diarrhoeagenicEscherichia coliclones by ribotyp-ing and ERIC-PCR. J. Med. Microbiol.47:227–234.
10. Del Vecchio, V. G., J. M. Petroziello, M. J. Gress, F. K. McCleskey, G. P. Melcher, H. K. Crouch, and J. R. Lupski.1995. Molecular genotyping of methicillin-resistantStaphylococcus aureusvia fluorophore-enhanced repet-itive-sequence PCR. J. Clin. Microbiol.33:2141–2144.
11. Farlow, J., K. L. Smith, J. Wong, M. Abrams, M. Lytle, and P. Keim.2001.
Francisella tularensisstrain typing using multiple-locus, variable-number tan-dem repeat analysis. J. Clin. Microbiol.39:3186–3192.
12. Fica, A. E., S. Prat Miranda, A. Fernandez Ricci, K. D’Ottone, and F. C. Cabello.1996. Epidemic typhoid in Chile: analysis by molecular and con-ventional methods ofSalmonella typhistrain diversity in epidemic (1977 and 1981) and nonepidemic (1990) years. J. Clin. Microbiol.34:1701–1707. 13. Franco, A., C. Gonzalez, O. S. Levine, R. Lagos, R. H. Hall, S. L. Hoffman,
M. A. Moechtar, E. Gotuzzo, M. M. Levine, D. M. Hone, et al.1992. Further consideration of the clonal nature ofSalmonella typhi: evaluation of molec-ular and clinical characteristics of strains from Indonesia and Peru. J. Clin. Microbiol.30:2187–2190.
14. Hood, D. W., M. E. Deadman, M. P. Jennings, M. Bisercic, R. D. Fleisch-mann, J. C. Venter, and E. R. Moxon.1996. DNA repeats identify novel virulence genes inHaemophilus influenzae.Proc. Natl. Acad. Sci. USA93:
11121–11125.
15. Ivanhoff, B.1995. Typhoid fever, global situation and W. H. O. recommen-dations. Southeast Asian J. Trop. Med. Public Health26(Suppl. 2):1–6. 16. Kamerbeek, J., L. Schouls, A. Kolk, M. van Agterveld, D. van Soolingen, S.
Kuijper, A. Bunschoten, H. Molhuizen, R. Shaw, M. Goyal, and J. van Embden.1997. Simultaneous detection and strain differentiation of Myco-bacterium tuberculosisfor diagnosis and epidemiology. J. Clin. Microbiol.
35:907–914.
17. Keim, P., L. B. Price, A. M. Klevytska, K. L. Smith, J. M. Schupp, R. Okinaka, P. J. Jackson, and M. E. Hugh-Jones.2000. Multiple-locus vari-able-number tandem repeat analysis reveals genetic relationships within
Bacillus anthracis.J. Bacteriol.182:2928–2936.
18. Klevytska, A. M., L. B. Price, J. M. Schupp, P. J. Worsham, J. Wong, and P. Keim.2001. Identification and characterization of variable-number tandem repeats in theYersinia pestisgenome. J. Clin. Microbiol.39:3179–3185. 19. Lachenauer, C. S., R. Creti, J. L. Michel, and L. C. Madoff.2000. Mosaicism
in the alpha-like protein genes of group Bstreptococci.Proc. Natl. Acad. Sci. USA97:9630–9635.
on May 15, 2020 by guest
http://jcm.asm.org/
20. Levinson, G., and G. A. Gutman.1987. Slipped-strand mispairing: a major mechanism for DNA sequence evolution. Mol. Biol. Evol.4:203–221. 21. Ling, J. M., N. W. Lo, Y. M. Ho, K. M. Kam, N. T. Hoa, L. T. Phi, and A. F.
Cheng.2000. Molecular methods for the epidemiological typing of Salmo-nella entericaserotype Typhi from Hong Kong and Vietnam. J. Clin. Micro-biol.38:292–300.
22. Nair, S., E. Schreiber, K. L. Thong, T. Pang, and M. Altwegg.2000. Geno-typic characterization ofSalmonella typhiby amplified fragment length poly-morphism fingerprinting provides increased discrimination as compared to pulsed-field gel electrophoresis and ribotyping. J. Microbiol. Methods41:
35–43.
23. Nair, S., C. L. Poh, Y. S. Lim, L. Tay, and K. T. Goh.1994. Genome fingerprinting ofSalmonella typhi by pulsed-field gel electrophoresis for subtyping common phage types. Epidemiol. Infect.113:391–402.
24. Pang, T.1998. Genetic dynamics ofSalmonella typhi-diversity in clonality. Trends Microbiol.6:339–342.
25. Pang, T., M. Altwegg, G. Martinetti, C. L. Koh, and S. Puthucheary.1992. Genetic variation among Malaysian isolates ofSalmonella typhias detected by ribosomal RNA gene restriction patterns. Microbiol. Immunol.36:539– 543.
26. Parkhill, J., G. Dougan, K. D. James, N. R. Thomson, D. Pickard, J. Wain, C. Churcher, K. L. Mungall, S. D. Bentley, M. T. Holden, M. Sebaihia, S. Baker, D. Basham, K. Brooks, T. Chillingworth, P. Connerton, A. Cronin, P. Davis, R. M. Davies, L. Dowd, N. White, J. Farrar, T. Feltwell, N. Hamlin, A. Haque, T. T. Hien, S. Holroyd, K. Jagels, A. Krogh, T. S. Larsen, S. Leather, S. Moule, P. O’Gaora, C. Parry, M. Quail, K. Rutherford, M. Simmonds, J. Skelton, K. Stevens, S. Whitehead, and B. G. Barrell.2001. Complete genome sequence of a multiple drug resistantSalmonella enterica
serovar Typhi CT18. Nature413:848–852.
27. Peak, I. R., M. P. Jennings, D. W. Hood, and E. R. Moxon.1999. Tetranucle-otide repeats identify novel virulence determinant homologues inNeisseria meningitidis.Microb. Pathog.26:13–23.
28. Peak, I. R., M. P. Jennings, D. W. Hood, M. Bisercic, and E. R. Moxon.1996. Tetrameric repeat units associated with virulence factor phase variation in
Haemophilusalso occur inNeisseriaspp.andMoraxella catarrhalis.FEMS Microbiol. Lett.137:109–114.
29. Renders, N., L. Licciardello, C. Ijsseldijk, M. Sijmons, L. van Alphen, H. Verbrugh, and A. van Belkum.1999. Variable numbers of tandem repeat loci in genetically homogeneousHaemophilus influenzaestrains alter during per-sistent colonisation of cystic fibrosis patients. FEMS Microbiol. Lett.173:
95–102.
30. Seib, K. L., I. R. Peak, and M. P. Jennings.2002. Phase variable restriction-modification systems inMoraxella catarrhalis.FEMS Immunol. Med. Micro-biol.32:159–165.
31. Selander, R. K., P. Beltran, N. H. Smith, R. Helmuth, F. A. Rubin, D. J. Kopecko, K. Ferris, B. D. Tall, A. Cravioto, and J. M. Musser.1990. Evo-lutionary genetic relationships of clones ofSalmonellaserovars that cause human typhoid and other enteric fevers. Infect. Immun.58:2262–2275. 32. Shangkuan, Y. H., and H. C. Lin.1998. Application of random amplified
polymorphic DNA analysis to differentiate different strains ofSalmonella typhiand otherSalmonellaspecies. J. Appl. Microbiol.85:693–702. 33. Strand, M., T. A. Prolla, R. M. Liskay, and T. D. Petes.1993. Destabilization
of tracts of simple repetitive DNA in yeast by mutations affecting DNA mismatch repair. Nature365:274–276.
34. Struelens, M. J., Y. De Gheldre, and A. Deplano.1998. Comparative and library epidemiological typing systems: outbreak investigations versus sur-veillance systems. Infect. Control Hosp. Epidemiol.19:565–569.
35. Thong, K. L., Y. L. Goh, R. M. Yasin, M. G. Lau, M. Passey, G. Winston, M. Yoannes, T. Pang, and J. C. Reeder.2002. Increasing genetic diversity of
Salmonella entericaserovar typhi isolates from Papua New Guinea over the period from 1992 to 1999. J. Clin. Microbiol.40:4156–4160.
36. Thong, K. L., A. M. Cordano, R. M. Yassin, and T. Pang.1996. Molecular analysis of environmental and human isolates ofSalmonella typhi.Appl. Environ. Microbiol.62:271–274.
37. Thong, K. L., Y. M. Cheong, S. Puthucheary, C. L. Koh, and T. Pang.1994. Epidemiologic analysis of sporadicSalmonella typhiisolates and those from outbreaks by pulsed-field gel electrophoresis. J. Clin. Microbiol.32:1135– 1141.
38. Thong, K. L., S. Puthucheary, R. M. Yassin, P. Sudarmono, M. Padmidewi, E. Soewandojo, I. Handojo, S. Sarasombath, and T. Pang.1995. Analysis of
Salmonella typhiisolates from Southeast Asia by pulsed-field gel electro-phoresis. J. Clin. Microbiol.33:1938–1941.
39. Threlfall, E. J., E. Torre, L. R. Ward, B. Rowe, and I. Gibert.1993. Insertion sequence IS200can differentiate drug-resistant and drug-sensitive Salmo-nella typhiof Vi-phage types E1 and M1. J. Med. Microbiol.39:454–458. 40. Threlfall, E. J., E. Torre, L. R. Ward, A. Davalos-Perez, B. Rowe, and I.
Gibert.1994. Insertion sequence IS200fingerprinting ofSalmonella typhi: an assessment of epidemiological applicability. Epidemiol. Infect.112:253–261. 41. van Belkum, A.1999. The role of short sequence repeats in epidemiologic
typing. Curr. Opin. Microbiol.2:306–311.
42. van Belkum, A., S. Scherer, W. van Leeuwen, D. Willemse, L. van Alphen, and H. Verbrugh.1997. Variable number of tandem repeats in clinical strains ofHaemophilus influenzae.Infect. Immun.65:5017–5027.
43. van Belkum, A., S. Scherer, L. van Alphen, and H. Verbrugh.1998. Short-sequence DNA repeats in prokaryotic genomes. Microbiol. Mol. Biol. Rev.
62:275–293.