0095-1137/96/$04.0010
Copyrightq1996, American Society for Microbiology
Rapid Characterization of Periodontal Bacterial Isolates by
Using Fluorogenic Substrate Tests
MARK F. J. MAIDEN,* ANNE TANNER,ANDPATRICK J. MACUCH
Department of Periodontal Microbiology, Forsyth Dental Center, Boston, Massachusetts 02115
Received 5 May 1995/Returned for modification 15 June 1995/Accepted 22 November 1995
Eighty-nine species of subgingival bacteria, represented by 121 reference strains and 892 patient isolates, including gram-negative, gram-positive, aerobic, facultatively anaerobic, microaerophilic, and anaerobic spe-cies, were characterized with a panel of fluorogenic, 4-methylumbelliferyl-linked substrate tests. Identifications of all patient isolates were confirmed by sodium dodecyl sulfate-polyacrylamide gel electrophoresis (SDS-PAGE) of whole-cell proteins relative to reference strains. Characteristic profiles of positive fluorogenic reactions differentiated most of the species, including five Porphyromonas species, six pigmenting and five nonpigmenting Prevotella species, Bacteroides forsythus, three Capnocytophaga species, six Actinomyces species, four Propionibacterium species, and eight Streptococcus species. Two mannoside isomers differentiated
Actino-myces israelii and ActinoActino-myces gerencseriae. In addition to Porphyromonas gingivalis, B. forsythus, and Capnocy-tophaga species, Fusobacterium alocis, Actinomyces odontolyticus, Actinomyces meyeri, and Bifidobacterium dentium
were all positive for so-called trypsin-like activity. Fusobacterium nucleatum, Eikenella corrodens, Actinobacillus
actinomycetemcomitans, and Campylobacter species were nonreactive with the carbohydrate-based substrates
tested. Fluorogenic substrate tests provided a sensitive and simple method for biochemical characterization that could presumptively identify to species level most subgingival isolates within 4 h. The method was ideal for rapidly obtaining presumptive identifications of isolates prior to confirming identifications by definitive methods, such as SDS-PAGE.
The characterization and identification of bacterial isolates have traditionally been based on phenotypic traits, which are derived from biochemical test reactions. One disadvantage of these tests has been the requirement of strains to grow in order to produce a detectable reaction. For fastidious or slowly grow-ing organisms such as many anaerobes, this requirement for growth has made biochemical testing very slow or, in some cases, not possible.
To overcome the limitations of traditional biochemical tests, rapid tests that do not require the growth of the test strains have been developed. These methods are based on the use of artificial substrates to detect preformed enzymes. Several com-mercial bacterial characterization and identification systems utilizing chromogenic substrates have been available for some years. These systems have been used to examine many of the species of oral bacteria (6, 14, 36, 40, 43, 44). However, these systems are designed with a fixed, preselected panel of sub-strates that are optimized for particular groups of species (mostly clinical) and are therefore not necessarily optimal for other groups, such as oral species.
An alternative method that uses fluorogenic substrates is now possible. A wide range of synthetic enzyme substrates based on conjugates of sugars, amino acids, and peptides with the fluorogen 4-methylumbelliferone (4MU) are commercially available. Enzymatic cleavage of these substrates yields a bright fluorescence that is readily and simply observed under long-wave UV illumination; no developing reagents are re-quired. Many fluorogenic tests with these compounds have been described as aids for the rapid identification of bacteria (22–24), including certain periodontal species (1, 18, 26–28, 37). More recently, a panel of fluorogenic tests has been used
for the differentiation of streptococci (3, 45). A commercial system based on fluorogenic substrates is also now available (9); however, no results for oral bacteria have been published. This study assessed the utility of a panel of fluorogenic substrates in the rapid characterization of a wide range of periodontal bacterial isolates. The fluorogenic substrates were chosen because they were simpler to use and were potentially more sensitive and rapid than chromogenic substrates.
MATERIALS AND METHODS
Type and reference strains of the major genera of oral bacteria were examined, including both gram-negative and gram-positive, aerobic, facultatively anaerobic, microaerophilic, and anaerobic species. The gram-negative species included
Por-phyromonas species, black-pigmented and nonpigmented Prevotella and Bacte-roides species, and Fusobacterium, Capnocytophaga, Campylobacter (Wolinella),
and Selenomonas species. The gram-positive species included Streptococcus,
Pep-tostreptococcus, Actinomyces, Propionibacterium, Bifidobacterium, and Eubacte-rium species. Fresh isolates of these species were obtained from subgingival
plaque samples cultured at Forsyth Dental Center from studies of the periodon-tal microflora of initial periodonperiodon-tal lesions, gingivitis, and healthy sites and of dental implants. Each strain was characterized by Gram stain reaction, cell morphology, and ability to grow in air. Most strains were maintained on TSBY blood agar (BBL Trypticase soy agar [20 g/liter], BBL brain heart infusion agar [26 g/liter], yeast extract [10 g/liter], hemin [5 mg/liter], defibrinated sheep blood [50 ml/liter]). Porphyromonas species were maintained on HK medium (BBL Trypticase soy agar [40 g/liter], hemin [5 mg/liter], menadione [0.5 mg/liter]), and
Bacteroides forsythus strains were maintained on NAM medium (BBL Trypticase
soy agar [40 g/liter], hemin [5 mg/liter], N-acetylmuramic acid [10 mg/liter], defibrinated sheep blood [50 ml/liter]). Campylobacter species were maintained on formate-fumarate medium (brain heart infusion agar [52 g/liter], supple-mented with yeast extract [10 g/liter], sodium formate [2 g/liter], disodium fu-marate [4.1 g/liter], hemin [5 mg/liter], and defibrinated sheep blood [50 ml/ liter]).
Fluorogenic substrate tests.The method for fluorogenic substrate tests was based on that of Whiley et al. (45). All of the substrates tested were from Sigma. The substrates included 4MU-a-L-arabinopyranoside (ARA), 4MU-b-D -cello-biopyranoside (CEL), 4MU-a-L-fucoside (FUC), 4MU-b-D-galactoside (bGA), 4MU-a-D-glucoside (aGL), 4MU-b-D-glucoside (bGL), 4MU-N-acetyl-b-D -glu-cosamide (NGL), 4MU-b-xyloside (XYL), benzoylarginine-7-amido,4-methyl-coumarin (TRP), 4MU-sulfate (SO4), proline-7-amido,4-methylcoumarin (PRO), 4MU-b-D-fucoside (bFU), 4MU-a-D-galactoside (aGA), 4MU-b-D -glu-curonide (bGN), 4MU-N-acetyl-b-D-galactosamide (NGA), 4MU-a-D
-man-* Corresponding author. Mailing address: Department of Periodon-tal Microbiology, Forsyth DenPeriodon-tal Center, 140 Fenway, Boston MA 02115. Phone: (617) 262-5200. Fax: (617) 262-4021. Electronic mail address: [email protected].
376
on May 15, 2020 by guest
http://jcm.asm.org/
nopyranoside (aMA), and 4MU-b-D-mannopyranoside (bMA). Each 4-MU-linked substrate was first prepared as a 1003stock solution in dimethyl sulfoxide at a concentration of 10 mg/ml. To prepare the tests, the substrate stock solutions were diluted to 0.1 mg/ml with TES buffer (Sigma; 50 mM, pH 7.5), and a 50-ml volume of each substrate was added to each well in one column of a 96-well microtiter tray. The first column of wells received TES buffer alone as a negative (no substrate) control. The stock solutions and predispensed trays were stored at
2208C and were stable in these conditions for at least 6 months.
Dense suspensions (A600of approximately 1.0, about 109cells per ml) of the bacterial strains to be tested were prepared in 0.6 ml of TES buffer with cells harvested from 3- to 5-day anaerobic cultures. For each strain, 50ml of suspen-sion was added to each well in one row of a microtiter tray, thus inoculating one well of each substrate plus the no substrate control. One row of substrates was inoculated with TES buffer to serve as a negative (uninoculated) control. After
incubation at 378C for 4 h, reactions were read under long-wave UV light (366 nm). Positive reactions were shown by a bright blue-white fluorescence. Positive test control organisms were B. forsythus ATCC 43037 and Prevotella oris ATCC 33573, and the negative test organism was Veillonella parvula ATCC 10790. Type and reference strains were tested on at least three separate occasions. Multiple fresh isolates of the species were each tested once before confirmation of their identifications by sodium dodecyl sulfate-polyacrylamide gel electrophoresis (SDS-PAGE).
[image:2.612.58.557.91.282.2]SDS-PAGE.The identifications of all fresh isolates were confirmed by SDS-PAGE of whole-cell proteins. This technique is well established as a definitive phenotypic method for strain identification (7, 13, 16, 20, 21, 25, 31, 34, 35, 39, 41, 42, 47). Briefly, a dense suspension of each strain in distilled water was sonicated, mixed with an equal volume of treatment buffer containing SDS and mercaptoethanol, and boiled for 5 min. A drop of Phenol Red tracking dye was
TABLE 1. Fluorogenic substrate reactions of reference strains and fresh isolates of Porphyromonas species and Bacteroides species and new
Bacteroides-like species
Organism Reference strain
(no. of isolates tested)a
Reaction with substrateb
ARA CEL FUC bGA aGL bGL NGL XYL TRP
Porphyromonas spp.
P. gingivalis 33277T(23) 2(13) 2(0) 2(0) 1(100) 2(0) 2(0) 1(100) 2(0) 1(100)
Oribaculum catoniaec 51270T(8) W (0) 2(0) 1(100) 1(88) 1(100) 2(0) 1(100) 2(0) 1(88)
P. asaccharolytica 25260T(1) 2(0) 2(0) 1(100) 2(0) 2(0) 2(0) 1(100) 2(0) 2(0)
P. endodontalis 25806T 2 2 2 2 2 2 1 2 2
P. circumdentariad 12469T 2 2 2 2 2 2 1 2 1
P. salivosad 11632T 2 2 2 1 1 2 1 2 1
B. forsythus 43037T(51) 1(67) 1(88) 1(100) 1(100) 1(98) 1(100) 1(100) 2(55) 1(100)
Bacteroides spp.
B. heparinolyticus 35895T 1 1 1 1 1 1 1 1 2
B. zoogleoformans 33285T 1 1 1 1 1 1 1 1 2
Bacteroides-like spp.e
J. ignava 51276T 2 2 2 2 2 2 2 2 2
C. morbi 51271T W 2 1 1 1 1 2 2 2
H. seregens 51272T W 2 2 1 1 2 1 2 2
a
All American Type Culture Collection strains except when noted. T, type strain.
b
W, weakly positive. Numbers in parentheses are the percentages of isolates giving a positive reaction.
c
Described by Moore and Moore (29) and reclassified as Porphyromonas catoniae by Willems and Collins (48).
d
Species isolated from cats by Love et al. (19).
e
Species described by Moore and Moore (29).
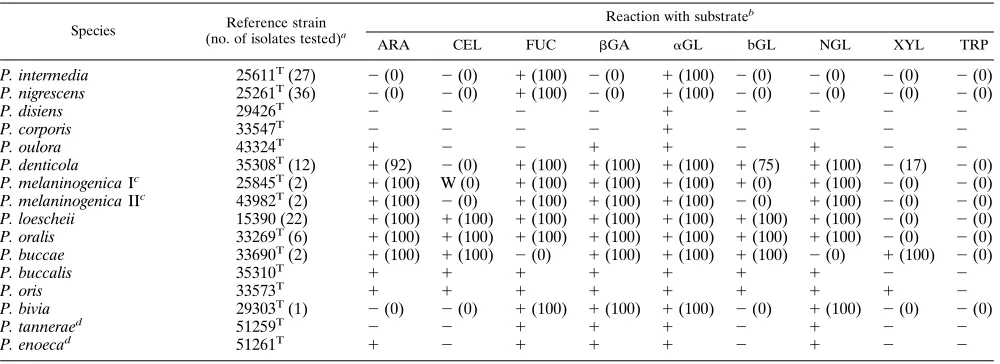
TABLE 2. Fluorogenic substrate reactions of reference strains and fresh isolates of Prevotella species
Species Reference strain
(no. of isolates tested)a
Reaction with substrateb
ARA CEL FUC bGA aGL bGL NGL XYL TRP
P. intermedia 25611T(27) 2(0) 2(0) 1(100) 2(0) 1(100) 2(0) 2(0) 2(0) 2(0)
P. nigrescens 25261T(36) 2(0) 2(0) 1(100) 2(0) 1(100) 2(0) 2(0) 2(0) 2(0)
P. disiens 29426T 2 2 2 2 1 2 2 2 2
P. corporis 33547T 2 2 2 2 1 2 2 2 2
P. oulora 43324T 1 2 2 1 1 2 1 2 2
P. denticola 35308T(12) 1(92) 2(0) 1(100) 1(100) 1(100) 1(75) 1(100) 2(17) 2(0)
P. melaninogenica Ic 25845T(2) 1(100) W (0) 1(100) 1(100) 1(100) 1(0) 1(100) 2(0) 2(0)
P. melaninogenica IIc 43982T(2) 1(100) 2(0) 1(100) 1(100) 1(100) 2(0) 1(100) 2(0) 2(0)
P. loescheii 15390 (22) 1(100) 1(100) 1(100) 1(100) 1(100) 1(100) 1(100) 2(0) 2(0)
P. oralis 33269T(6) 1(100) 1(100) 1(100) 1(100) 1(100) 1(100) 1(100) 2(0) 2(0)
P. buccae 33690T(2) 1(100) 1(100) 2(0) 1(100) 1(100) 1(100) 2(0) 1(100) 2(0)
P. buccalis 35310T 1 1 1 1 1 1 1 2 2
P. oris 33573T 1 1 1 1 1 1 1 1 2
P. bivia 29303T(1) 2(0) 2(0) 1(100) 1(100) 1(100) 2(0) 1(100) 2(0) 2(0)
P. tanneraed 51259T 2 2 1 1 1 2 1 2 2
P. enoecad 51261T 1 2 1 1 1 2 1 2 2
a
All American Type Culture Collection strains except when noted. T, type strain.
b
W, weakly positive. Numbers in parentheses are percentages of isolates giving a positive reaction.
c
DNA homology groups of Holdeman and Johnson (10).
d
Species described by Moore et al. (30).
on May 15, 2020 by guest
http://jcm.asm.org/
[image:2.612.58.558.495.677.2]then added to each preparation. A sample (5 to 15ml) of each preparation was loaded into a well of a 4% (wt/vol) acrylamide stacking gel overlying a 12% acrylamide separating gel (31). On each gel, type or reference strains appropriate for the presumptive identification of the unknown strains were run along with the unknowns. The electrophoresis was run for 2 h at a constant current of 30 mA per gel, and the gels are then stained by a modified Coomassie blue method (8). The identity of each isolate was determined by visual comparison with the reference strains on the same gel.
RESULTS
The reactions of each group of oral species with the sub-strates tested are listed in Tables 1 to 11. No fluorescence was detected in uninoculated substrate controls or in inoculated no substrate controls. Positive reactions were usually uniformly strong, but some individual isolates gave weakly positive reac-tions with one or more substrates (approximately 20% of all positive reactions recorded). These weakly positive reactions were clearly distinguishable from negative reactions and in most cases matched positive reactions in reference strains and other isolates.
Characteristic profiles of positive fluorogenic reactions, in combination with Gram staining and aerotolerance, differen-tiated most of the species examined. The reaction profiles obtained for the reference strains were consistent between separate test occasions, with only minor variations in the strengths of some reactions. For the patient isolates, multiple isolates of some species gave profiles identical or very similar to each other and to those of the reference strains. A few
species showed more variation among the profiles obtained from fresh strains; however, the profiles were still characteris-tic. Table 12 shows the degrees to which the profiles of patient isolates differed from those of the corresponding reference strains. Several species were unreactive with the carbohydrate-based substrates, and some did not react with any of the sub-strates. For some of the species examined, no representatives were identified among the fresh patient isolates. For these species, the profiles of the type or reference strains are given. The Porphyromonas species and the closely related B. for-sythus (Table 1) were all positive with the NGL substrate. Porphyromonas gingivalis, B. forsythus, and two animal oral species, Porphyromonas circumdentaria and Porphyromonas salivosa, were all characterized by a positive reaction with the TRP (trypsin-like) substrate. P. gingivalis and Porphyromonas asaccharolytica were further differentiated by their reactions with the FUC and bGA substrates, and Porphyromonas end-odontalis reacted only with the NGL substrate. P. circumden-taria and P. salivosa were differentiated by their reactions with thebGA andaGL substrates, and B. forsythus strains reacted with all of the carbohydrate-based substrates. The type strains of Bacteroides heparinolyticus and Bacteroides zoogleoformans also reacted with all the carbohydrate-based substrates and gave identical reaction profiles. However, these two species were easily distinguished by SDS-PAGE protein profiles.
The type strains of four newly proposed species (29) were examined (Table 1). Johnsonella ignava, which has been de-TABLE 3. Fluorogenic substrate reactions of reference strains and fresh isolates of Selenomonas species and Campylobacter species
Organism Reference strain
(no. of isolates tested)a
Reaction with substrateb
ARA CEL FUC bGA aGL bGL NGL XYL TRP SO4
Selenomonas artemidis 43528T(1) 2(0) 2(0) 2(0) 2(0) 1(100) 2(0) 2(0) 2(0) 2(0) NT (NT)
Selenomonas dianae 43527T(2) W (0) 2(0) 2(0) 1(100)2(0) 2(0) 2(0) 2(0) 2(0) NT (0)
Selenomonas flueggei 43531T(10) 1(100) 2(0) 2(0) 1(100) W (100) 2(0) 2(0) 2(0) 2(0) NT (10)
Selenomonas infelix 43532T(5) 1(100) 2(0) 2(0) 1(100) W (100) 2(20) 2(0) 2(20) 2(0) NT (NT)
Selenomonas noxia 43541T(7) 2(0) 2(0) 2(0) 2(0) 2(0) 2(0) 2(0) 2(0) 2(0) NT (0)
Selenomonas sputigena 35185T(9) 1(100) 2(0) 2(0) 1(100) W (100) 2(11) 2(67) W (22) 2(0) NT (0)
Campylobacter rectus, C. showae, C. concisus, and C. curvus
—c 2 2 2 2 2 2 2 2 2 2
C. gracilis 33236T(25) 2(0) 2(0) 2(0) 2(0) 2(0) 2(0) 2(0) 2(0) 2(0)2(33)
aAll American Type Culture Collection strains except when noted. T, type strain.
bW, weakly positive; NT, not tested. Numbers in parentheses are percentages of isolates giving a positive reaction.
[image:3.612.59.556.83.212.2]cC. rectus ATCC 33238T, C. showae ATCC 51146T, C. concisus ATCC 33237T, C. curvus ATCC 35224T.
TABLE 4. Fluorogenic substrate reactions of reference strains and fresh isolates of Fusobacterium, Leptotrichia, and Capnocytophaga species
Organism Reference strain
(no. of isolates tested)a
Reaction with substrateb
ARA CEL FUC bGA aGL bGL NGL XYL TRP SO4
F. nucleatum subsp. polymorphumc 10953T(14) 2(0) 2(0) 2(0) 2(0) 2(0) 2(0) 2(0) 2(0) 2(0) 1(86)
F. periodonticum 33693T 2 2 2 2 2 2 2 2 W 1
F. alocis 35896T(3) 2(0) 2(0) 2(0) 2(0) 2(0) 2(0) 2(0) 2(0) 1(100) 2(0)
L. buccalis 23471T 1 2 2 1 1 2 2 2 2 2
C. ochracea 27872T(2) 1(0) 1(100) 2(0) 1(100)1(100)1(100) 1(100)2(0) 1(100) NT (0)
C. sputigena 33612T(6) 1(83) 1(100) 2(0) 1(100)1(100)1(100) 1(67) 1(100)1(100) NT (0)
C. gingivalis 33624T(20) 2(15) 2(0) 2(0) 1(85) 1(95) 1(85) W (0) 2(45) 1(100) NT (0)
aAll American Type Culture Collection strains except when noted. T, type strain.
bW, weakly positive; NT, not tested. Numbers in parentheses are percentages of isolates giving a positive reaction.
cF. nucleatum subsp. nucleatum ATCC 25586Tand 15 isolates, F. nucleatum subsp. vincentii ATCC 49256Tand 15 isolates, F. nucleatum subsp. fusiforme NCTC
11326Tand 3 isolates, F. sulci ATCC 35585T, and F. nechrophorum ATCC 25286Twere negative with all of the substrates.
on May 15, 2020 by guest
http://jcm.asm.org/
[image:3.612.59.555.569.685.2]scribed as nonfermentative, was correspondingly unreactive with any of the substrates. Oribaculum catoniae, Catonella morbi, and Hallella seregens each produced distinctive fluoro-genic profiles. O. catoniae strains reacted with the trypsin-like substrate and most closely resembled B. forsythus strains in their fluorogenic reaction profiles.
Prevotella intermedia and Prevotella nigrescens (formerly two DNA homology groups of Prevotella intermedia [11]) gave identical profiles, as did two homology groups of Prevotella melaninogenica (Table 2). Prevotella corporis and Prevotella dis-iens reacted only with theaGL substrate. Prevotella tannerae, a newly described species (30), gave the same pattern of reac-tions as Prevotella bivia, and Prevotella enoeca (30) gave a similar pattern.
The nonpigmenting Prevotella species P. buccae and P. oris, which are difficult to distinguish biochemically, were separated by their reactions with the NGL and FUC (Table 2), as previ-ously reported for a commercial, chromogenic test (6). In ad-dition, reaction with the fluorogenic XYL substrate differen-tiated P. oris from Prevotella buccalis and Prevotella loescheii. The fluorogenic substrates did not differentiate P. loescheii, P. buccalis, and P. oralis, but P. loescheii can usually be distin-guished by longer cells and the formation of brown pigment in most (but not all) strains.
The Selenomonas species (Table 3) reacted with few of the
fluorogenic substrates. Selenomonas artemidis reacted only with theaGL substrate, and Selenomonas noxia was unreactive with all of the substrates. The other species all gave character-istic patterns of ARA, bGA, and a-glucoside reactions. Al-though these reactions did not differentiate the Selenomonas species, they did distinguish them as a group from the mor-phologically similar Campylobacter species which were unreac-tive with all of the substrates. One third of Campylobacter (Bacteroides) gracilis strains reacted weakly with the sulfate substrate.
[image:4.612.58.555.95.165.2]Positive reactions with SO4differentiated Fusobacterium nu-cleatum subspecies polymorphum strains and the Fusobacte-rium periodonticum type strain from F. nucleatum subspecies nucleatum, F. nucleatum subspecies vincentii, and F. nucleatum subspecies fusiforme, Fusobacterium nechrophorum, and Fuso-bacterium sulci, which were negative for all tests (Table 4). Fusobacterium alocis strains were positive with the trypsin-like substrate but were negative with all other tests. The Capnocy-tophaga species all reacted with the trypsin-like reagent (Table 4), as reported previously for a chromogenic version of this substrate (18). Capnocytophaga sputigena and Capnocytophaga ochracea differed in their reactions with the ARA and XYL substrates. Capnocytophaga gingivalis and related strains were all negative with the CEL substrate, whereas C. sputigena and C. ochracea strains were all positive.
TABLE 5. Fluorogenic substrate reactions of reference strains and fresh isolates of Eikenella, Actinobacillus, Haemophilus, and
Cardiobacterium species
Organism Reference strain
(no. of isolates tested)a
Reaction with substrateb
ARA CEL FUC bGA aGL bGL NGL XYL TRP SO4 PRO
E. corrodens 23834T 2 2 2 2 2 2 2 2 2 2 1
A. actinomycetemcomitans 29523 2 2 2 2 2 2 2 2 2 NT NT
H. aprophilus 33389T(9) 1(56) 2(0) 2(0) 1(100) 1(0) 2(0) 2(0) 2(0) 2(0) NT (0) NT (0)
C. hominis 15826T(5) 2(0) 2(0) 2(0) 2(0) 2(0) 2(0) 2(0) 2(0) 1(100) 2(0) 2(0)
a
All American Type Culture Collection strains except when noted. T, type strain.
b
W, weakly positive; NT, not tested. Numbers in parentheses are percentages of isolates giving a positive reaction.
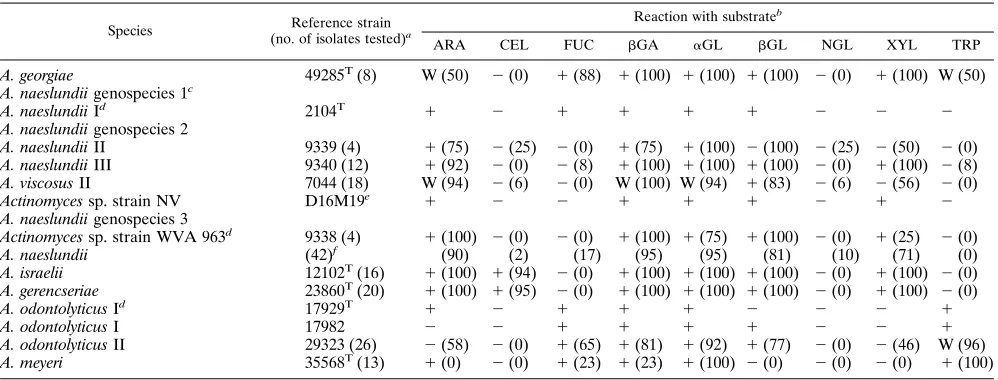
TABLE 6. Fluorogenic substrate reactions of reference strains and fresh isolates of Actinomyces species
Species Reference strain
(no. of isolates tested)a
Reaction with substrateb
ARA CEL FUC bGA aGL bGL NGL XYL TRP
A. georgiae 49285T(8) W (50) 2(0) 1(88) 1(100) 1(100) 1(100) 2(0) 1(100) W (50)
A. naeslundii genospecies 1c
A. naeslundii Id 2104T 1 2 1 1 1 1 2 2 2
A. naeslundii genospecies 2
A. naeslundii II 9339 (4) 1(75) 2(25) 2(0) 1(75) 1(100) 2(100) 2(25) 2(50) 2(0)
A. naeslundii III 9340 (12) 1(92) 2(0) 2(8) 1(100) 1(100) 1(100) 2(0) 1(100) 2(8)
A. viscosus II 7044 (18) W (94) 2(6) 2(0) W (100) W (94) 1(83) 2(6) 2(56) 2(0)
Actinomyces sp. strain NV D16M19e 1 2 2 1 1 1 2 1 2
A. naeslundii genospecies 3
Actinomyces sp. strain WVA 963d 9338 (4) 1(100) 2(0) 2(0) 1(100) 1(75) 1(100) 2(0) 1(25) 2(0)
A. naeslundii (42)f (90) (2) (17) (95) (95) (81) (10) (71) (0)
A. israelii 12102T(16) 1(100) 1(94) 2(0) 1(100) 1(100) 1(100) 2(0) 1(100) 2(0)
A. gerencseriae 23860T(20) 1(100) 1(95) 2(0) 1(100) 1(100) 1(100) 2(0) 1(100) 2(0)
A. odontolyticus Id 17929T 1 2 1 1 1 2 2 2 1
A. odontolyticus I 17982 2 2 1 1 1 1 2 2 1
A. odontolyticus II 29323 (26) 2(58) 2(0) 1(65) 1(81) 1(92) 1(77) 2(0) 2(46) W (96)
A. meyeri 35568T(13) 1(0) 2(0) 1(23) 1(23) 1(100) 2(0) 2(0) 2(0) 1(100)
a
All American Type Culture Collection strains except when noted. T, type strain.
b
W, weakly positive. Numbers in parentheses are percentages of isolates giving a positive reaction.
c
DNA homology groups probably representing separate species (Johnson et al. [12]).
d
I, II, III, and WVA 963, serotype designations.
e
Virginia State Polytechnic Institute Anaerobe Laboratory culture collection.
f
Fresh isolates not identifiable to genospecies level.
on May 15, 2020 by guest
http://jcm.asm.org/
[image:4.612.58.557.488.678.2]Strains of Eikenella corrodens, Actinobacillus actinomycetem-comitans, and Cardiobacterium hominis were negative with the carbohydrate-based substrates (Table 5). E. corrodens gave a positive reaction with the PRO substrate, as reported previ-ously with a commercial chromogenic system (40). C. hominis strains reacted with the trypsin-like substrate, in agreement with results from chromogenic tests (4). A positive reaction with thebGA substrate differentiated Haemophilus aphrophi-lus from A. actinomycetemcomitans.
The Actinomyces species tested (Table 6) included several recently described taxa: Actinomyces georgiae, Actinomyces ger-encseriae, and three DNA homology groups (genospecies) of Actinomyces naeslundii (12). A. georgiae gave a characteristic profile that permitted strain identification. A. naeslundii strains overall gave similar profiles, including isolates identified as one of the serotypes, or genospecies, and a large number of isolates identifiable within none of these taxa. There were no clear differences between the profiles of the serotypes or the pro-posed genospecies.
Actinomyces israelii and A. gerencseriae differed from strains of A. naeslundii and other Actinomyces species by reacting with the cellobioside substrate. A. gerencseriae (previously A. israelii serotype II) gave a profile identical to that of A. israelii, and, therefore, these two species could not be differentiated by these substrates. When additional substrates were tested, two isomers of mannopyranoside distinguished A. israelii strains, which degraded theb-mannoside substrate, from A. gerencse-riae strains, which degraded thea-mannoside substrate (Table 7). Strains of both Actinomyces odontolyticus and Actinomyces meyeri gave positive reactions with the trypsin-like substrate, which differentiated them from the other Actinomyces species. A. meyeri strains differed from A. odontolyticus in being
nega-tive with thebGL and XYL substrates. Many fresh isolates of A. meyeri were positive with only the aGL and trypsin-like substrates.
Strains of Propionibacterium (previously Arachnia) propioni-cus produced profiles very similar to that of A. odontolytipropioni-cus (Table 8). Propionibacterium acnes strains gave a characteristic profile of bGL, aGL, and NGL reactions, although several fresh isolates reacted only with the NGL substrate. Lactoba-cillus uli strains reacted only with thebGL substrate. The type strains of Propionibacterium avidum, Propionibacterium granu-losum, Lactobacillus oris, Atopobium parvulum, Atopobium rimae, and Corynebacterium matruchotii all gave distinctive re-action profiles. No fresh strains of any of these species were isolated. The type strain of C. matruchotii was a further exam-ple of a gram-positive organism that reacted with the trypsin-like substrate. Rothia dentocariosa also produced a character-istic reaction pattern; however, fresh isolates gave a profile simpler than that of the type strain, reacting only with theaGL substrate.
Bifidobacterium dentium (Table 9) produced a profile that included a positive reaction with the trypsin-like substrate. The reaction profile of Eubacterium saburreum differentiated this species from the morphologically similar F. nucleatum strains, which were unreactive with the carbohydrate-based substrates (Table 4). The type strain of Eubacterium limosum reacted only weakly with thebGL substrate. Eubacterium brachy, Eubacte-rium lentum, EubacteEubacte-rium nodatum, and EubacteEubacte-rium timidum strains were all unreactive with the test panel.
[image:5.612.57.557.83.139.2]The majority of Streptococcus species examined were readily differentiated by the test panel (Table 10). Streptococcus gor-donii was positive with the CEL and FUC substrates, which distinguished it from Streptococcus sanguis. The type strain of TABLE 7. Additional fluorogenic substrate reactions of reference strains and fresh isolates of A. israelii and A. gerencseriae
Species Reference strain
(no. of isolates tested)a
Reaction with substrateb
bFU aGA bGN NGA aMA bMA
A. israelii 12102T(6) 1(100) 1(100) W (67) 2(0) 2(0) 1(100)
A. gerencseriae 23860T(17) 1(100) 1(100) 2(0) 2(0) 1(100) 2(0)
aAll American Type Culture Collection strains except when noted. T, type strain.
bW, weakly positive. Numbers in parentheses are percentages of isolates giving a positive reaction.
TABLE 8. Fluorogenic substrate reactions of reference strains and fresh isolates of Propionibacterium, Lactobacillus, and Atopobium species,
R. dentocariosa, and C. matruchotii
Organism Reference strain (no. of isolates tested)a
Reaction with substrateb
ARA CEL FUC bGA aGL bGL NGL XYL TRP
P. propionicus I 14157T 1 2 1 1 1 1 2 W NT
29324 W 2 1 1 1 2 2 2 2
P. propionicus II 29326 2 2 2 1 1 1 2 2 2
P. propionicus (4)c (25) (0) (75) (75) (100) (75) (0) (75) (100)
P. acnes 11828T(45) 2(0) 2(0) 2(0) W (87) 1(58) 2(0) 1(100) 2(0) 2(0)
P. avidum 25577T 2 2 2 1 1 1 1 1 2
P. granulosum 25564T 2 2 2 2 1 2 2 2 2
L. oris 49062T 1 2 2 1 1 1 2 1 2
L. uli 49627T(5) 2(0) 2(0) 2(0) 2(0) 2(0) 1(100) 2(0) 2(0) 2(0)
A. parvulum 33793T 2 W 2 1 2 W W 2 2
A. rimae 49626T 2 2 2 2 2 2 W 2 2
R. dentocariosa 17931T(6) 2(0) 2(0) 2(0) 1(0) 1(100) 2(0) 1(0) 2(0) 2(0)
C. matruchotii 14266T 2 2 2 2 W 2 2 2 1
a
All American Type Culture Collection strains except when noted. T, type strain.
b
W, weakly positive; NT, not tested. Numbers in parentheses are percentages of isolates giving a positive reaction.
c
Fresh isolates not identified as P. propionicus I or II.
on May 15, 2020 by guest
http://jcm.asm.org/
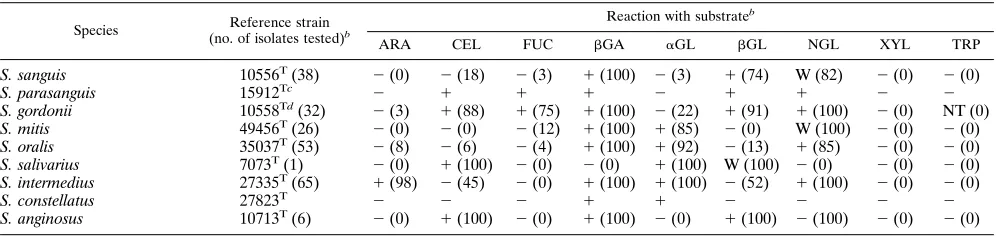
[image:5.612.57.558.549.703.2]Streptococcus parasanguis (46) gave the same profile as S. gor-donii but was different by SDS-PAGE protein profile. Strepto-coccus oralis and StreptoStrepto-coccus mitis strains gave very similar reaction profiles and could not be differentiated by this panel of tests. Streptococcus pneumoniae gave reactions very similar to those of S. oralis. Streptococcus salivarius gave a character-istic profile, and each of the three ‘‘Streptococcus milleri’’ spe-cies (Streptococcus intermedius, Streptococcus constellatus, and Streptococcus anginosus) were differentiated by the tests. S. intermedius was the only Streptococcus species to be consis-tently positive with the ARA substrate.
Peptostreptococcus micros and Peptostreptococcus magnus (Table 11) were negative with all of the carbohydrate-based substrates, and Peptostreptococcus anaerobius reacted only with the aGL substrate. The type strain of P. micros and about a third of fresh isolates gave weak or positive reactions with the trypsin-like substrate. All P. micros strains and the P. anaero-bius type strain were positive with the PRO substrate. Strains of Veillonella species were all nonreactive with all of the tests, except for a small proportion of Veillonella atypica isolates that reacted with the sulfate substrate. The type strain of Neisseria mucosa was unreactive in all the tests, and the type strain of Neisseria sicca reacted with only the PRO substrate. However, the one patient isolate of N. sicca that was identified also reacted with the arabinoside andbGA substrates.
DISCUSSION
Most of the subgingival bacterial species examined in this study produced characteristic profiles of positive reactions with the panel of fluorogenic substrates that were tested. The reac-tion profiles obtained, in combinareac-tion with a Gram stain and aerotolerance test, enabled the majority of fresh isolates to be presumptively identified to the corresponding species. The
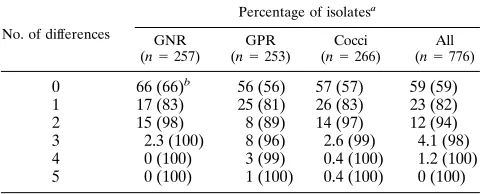
numbers of isolates tested for many of the species were too low to generate a stand-alone identification scheme (for which many more strains would be required), but the results did show that the profiles were very consistent. The majority (59%) of fresh isolates gave profiles identical to those of the correspond-ing reference strains, and 94% differed by no more than two reactions (Table 12). The most varied profiles were seen in the gram-positive rods, in particular among A. naeslundii and A. odontolyticus strains. In the case of A. naeslundii, this may be related to the known heterogeneity of the species (12). The present results did not, however, show any consistent associa-tion between fluorogenic test profiles and the proposed geno-species groupings of A. naeslundii.
[image:6.612.57.554.86.149.2]The individual reactions of the species tested with the flu-orogenic substrates agreed in general with the results reported previously with commercial systems based on p-nitrophenol-andb-naphthylamide-linked chromogenic substrates (6, 14, 36, 40, 43, 44). The panel of fluorogenic substrates tested worked equally well with aerobic, facultatively anaerobic, microaero-philic, and anaerobic species. For all of these organisms, the test trays could be incubated in a single atmosphere, air, which is an advantage over growth-based biochemical tests. The flu-orogenic substrate tests were simpler than the corresponding chromogenic substrate tests, since no color development re-agents were required. This also meant that the reactions could be reincubated after reading when a stronger reaction was sought. In many cases, it was possible to read some of the reactions within minutes of inoculation, but some species and substrate combinations required longer incubations (2 to 4 h) to produce a strong positive reaction. In practice, a 4-h incu-bation was chosen to obtain the strongest reactions while still permitting the tests to be completed within the laboratory working day. The microwell plate format was more convenient for performing these longer incubations than spot tests on TABLE 9. Fluorogenic substrate reactions of reference strains and fresh isolates of Bifidobacterium and Eubacteriumaspecies
Organism Reference strain
(no. of isolates tested)b
Reaction with substratec
ARA CEL FUC bGA aGL bGL NGL XYL TRP
B. dentium 27534 (8) 1(100) W (25) 2(13) 1(63) 1(88) 1(75) 2(0) 1(100) 1(75)
E. limosum 8486T 2 2 2 2 2 W 2 2 2
E. saburreum 33271T(5) 1(60) 2(0) 1(100) 1(100) 1(100) 1(80) 2(20) 2(0) 2(0)
a
E. brachy ATCC 33089T
and 8 isolates, E. timidum ATCC 33092T
and 8 isolates, E. lentum ATCC 25559T
, and E. nodatum ATCC 33099T
were all negative for all of the substrates.
b
All American Type Culture Collection strains except when noted. T, type strain.
c
W, weakly positive. Numbers in parentheses are percentages of isolates giving a positive reaction.
TABLE 10. Fluorogenic substrate reactions of reference strains and fresh isolates of Streptococcus species
Species Reference strain
(no. of isolates tested)b
Reaction with substrateb
ARA CEL FUC bGA aGL bGL NGL XYL TRP
S. sanguis 10556T(38) 2(0) 2(18) 2(3) 1(100) 2(3) 1(74) W (82) 2(0) 2(0)
S. parasanguis 15912Tc 2 1 1 1 2 1 1 2 2
S. gordonii 10558Td(32) 2(3) 1(88) 1(75) 1(100) 2(22) 1(91) 1(100) 2(0) NT (0)
S. mitis 49456T(26) 2(0) 2(0) 2(12) 1(100) 1(85) 2(0) W (100) 2(0) 2(0)
S. oralis 35037T(53) 2(8) 2(6) 2(4) 1(100) 1(92) 2(13) 1(85) 2(0) 2(0)
S. salivarius 7073T(1) 2(0) 1(100) 2(0) 2(0) 1(100) W (100) 2(0) 2(0) 2(0)
S. intermedius 27335T(65) 1(98) 2(45) 2(0) 1(100) 1(100) 2(52) 1(100) 2(0) 2(0)
S. constellatus 27823T 2 2 2 1 1 2 2 2 2
S. anginosus 10713T(6) 2(0) 1(100) 2(0) 1(100) 2(0) 1(100) 2(100) 2(0) 2(0)
a
All American Type Culture Collection strains except when noted. T, type strain.
b
W, weakly positive; NT, not tested. Numbers in parentheses are percentages of isolates giving a positive reaction.
c
National Collection of Type Cultures strain number.
d
Biovar 2 described by Kilian et al. (15). Strains ATCC 12396 (biovar 1) and ATCC 33399 (biovar 3) gave the same profile.
on May 15, 2020 by guest
http://jcm.asm.org/
[image:6.612.58.556.576.694.2]filter paper (28) or direct application of substrate solution to colonies on agar plates (1, 37). Incubation for more than 4 to 6 h gave somewhat stronger reactions with a few weakly react-ing strains but in most cases did not have any advantage over 4-h incubation.
The panel of substrates described was selected to best char-acterize the widest range of oral species based on published data with other systems and substrates. The tests were there-fore able to include a combination of substrates not included in any single commercial system. One example was the inclusion of the XYL substrate which aided the differentiation of P. oris from P. buccalis and P. loescheii, which was not possible with the RapID-ANA system (6). Another very useful differentia-tion was provided by the two mannoside isomers, which sepa-rated A. israelii and A. gerencseriae. If used as a double-spot test, these two substrates could provide a very quick and easy method for distinguishing these two species. It should be noted that more fluorogenic substrates are available than were in-cluded in the panel tested, including many for aminopepti-dases. Some of these may be useful for characterizing certain groups of oral isolates, such as the asaccharolytic species.
Apart from some of the limitations discussed above, the several available commercial systems have been shown to be useful for differentiating various groups of oral bacteria (6, 14, 36, 40, 43, 44). One additional advantage of the fluorogenic substrate method was its relative cheapness; the estimated total cost of the panel of tests described is less than 50 cents per strain, versus at least 2 dollars for the commercial systems. There were a few differences noted from published data for certain species and substrates. On the basis of chromogenic substrate tests, negative reactions were reported forbGA in P. gingivalis and P. salivosa, for NGL in P. asaccharolytica, P. endodontalis, and P. circumdentaria, and for trypsin-like activity in P. circumdentaria (5, 19). The corresponding fluoro-genic substrates all produced positive reactions with these spe-cies (Table 1). These differences could be due to differences in the sensitivities of the fluorogenic and chromogenic substrates, differences in the accessibility of the substrates to the enzymes (either by cell permeability or by enzyme active-site topology), or nutritional differences due to a different culture medium. With some of the Streptococcus species (Table 10), there were some differences noted from published results with the same fluorogenic substrates (3, 45). In this case, the different reac-tions were most probably due to differences in the basal media on which the strains were cultured in the different studies.
Possession of an enzyme activity that degrades artificial tryp-sin substrates such as benzoylarginine-b-naphthylamide (BANA), so-called trypsin-like activity, has been proposed as a possible virulence factor in the suspected periodontal pathogens P. gingivalis, B. forsythus, and Treponema denticola (2, 17, 33). Capnocytophaga species also have this activity. One interesting finding from the present study was that several other oral species also gave positive reactions with the fluorogenic version of the trypsin-like substrate. These included the gram-positive species A. odontolyticus, A. meyeri, C. matruchotii, and B. den-tium and the gram-negative species F. alocis and C. hominis. The possible relevance of this observation to the roles of these other species in periodontal infections is unclear.
The positive reactions of P. gingivalis with thebGA substrate and of P. asaccharolytica with the FUC substrate were inter-esting considering the asaccharolytic nature of these organ-isms. These observations were consistent with previous reports (43, 44). It is possible that these enzymes are used by the organisms to alter components of host cells or salivary com-ponents; there is increasing interest in the hexosaminidase activities of P. gingivalis as potential virulence factors.
[image:7.612.62.556.83.227.2]Several species did not react with any of the substrates; however, this unreactivity was a differential characteristic that could distinguish these species from morphologically similar, reactive species. For example, the Campylobacter species were distinguished from the reactive Selenomonas species, and F. nucleatum strains were distinguished from Capnocytophaga species and Eubacterium saburreum (a gram-positive species TABLE 11. Fluorogenic substrate reactions of reference strains and fresh isolates of Peptostreptococcus, Veillonella, and Neisseria species
Organism Reference strain (no. of isolates tested)a
Reaction with substrateb
ARA CEL FUC bGA aGL bGL NGL XYL TRP SO4 PRO
P. micros 33270T(33) 2(0) 2(0) 2(0) 2(0) 2(0) 2(0) 2(0) 2(0) W (27) 2(0) 1(100)
P. magnus 15794T 2 2 2 2 2 2 2 2 2 NT 2
P. anaerobius 27337T 2 2 2 2 1 2 2 2 2 NT 1
V. atypica 17744T(9) 2(0) 2(0) 2(0) 2(0) 2(0) 2(0) 2(0) 2(0) 2(0) NT (11) 2(0)
V. parvula 10790T(6) 2(0) 2(0) 2(0) 2(0) 2(0) 2(0) 2(0) 2(0) 2(0) NT (0) 2(0)
V. dispar 17748T 2 2 2 2 2 2 2 2 2 NT 2
D. pneumosintesc 33048T 2 2 2 2 2 2 2 2 2 2 2
N. mucosa 19696T 2 2 2 2 2 2 2 2 2 2 NT
N. sicca 29256T(1) 2(100) 2(0) 2(0) 2(100) 2(0) 2(0) 2(0) 2(0) 2(0) 2(0) 1(100)
aAll American Type Culture Collection strains except when noted. T, type strain.
bW, weakly positive; NT, not tested. Numbers in parentheses are percentages of isolates giving a positive reaction. c
Previously Bacteroides pneumosintes (Willems and Collins [49]).
TABLE 12. Degrees of differences between the fluorogenic reaction profiles of patient isolates and reference strains
No. of differences
Percentage of isolatesa
GNR (n5257)
GPR (n5253)
Cocci (n5266)
All (n5776)
0 66 (66)b 56 (56) 57 (57) 59 (59)
1 17 (83) 25 (81) 26 (83) 23 (82)
2 15 (98) 8 (89) 14 (97) 12 (94)
3 2.3 (100) 8 (96) 2.6 (99) 4.1 (98) 4 0 (100) 3 (99) 0.4 (100) 1.2 (100) 5 0 (100) 1 (100) 0.4 (100) 0 (100)
a
GNR, gram-negative rods (Tables 1 to 5); GPR, gram-positive rods (Tables 6 to 9). Cocci appear in Tables 10 and 11. Numbers in parentheses are cumu-lative percentages of isolates.
on May 15, 2020 by guest
http://jcm.asm.org/
[image:7.612.315.555.605.702.2]that often gives a gram-negative stain). A. actinomycetemcomi-tans strains also failed to react with any of the carbohydrate-based substrates tested, differentiating them from H. aphrophi-lus strains. A. actinomycetemcomitans strains do ferment a range of carbohydrates, and their failure to react with the fluorogenic substrates may have been due to poor permeability of the synthetic substrates.
Some groups of related species showed similar patterns of fluorogenic reactions, and for these species the results of the fluorogenic tests were compared with published phylogenies based on 16S rRNA sequence data (32, 38). P. intermedia, P. nigrescens, P. disiens, P. corporis, and Prevotella oulora consti-tute a subcluster within the Prevotella cluster of the Bacteroides subgroup, and these were all characterized by a small number of fluorogenic reactions. The P. melaninogenica and P. oralis groups, which constitute the two other Prevotella subclusters, all reacted with many more of the substrates. The Porphyromo-nas cluster includes the only members of the Bacteroides sub-division, three Porphyromonas species, ‘‘Bacteroides’’ macacae, and B. forsythus, that demonstrate the so-called trypsin-like activity against benzoylarginine based substrates. O. catoniae, one of the species proposed by Moore and Moore (29) (Table 1), was found to react with the TRP substrate, suggesting that it also belongs to the Porphyromonas cluster. This possibility has recently been confirmed by phylogenetic analysis based on 16S rRNA sequencing (48), which clearly places O. catoniae in the genus Porphyromonas. Dialister (previously Bacteroides) pneumosintes has been reported to belong to the Sporomusa branch of the Clostridium subphylum, with Veillonella and Me-gasphera species as its closest relatives (49). In the fluorogenic tests, the type strain of D. pneumosintes was unreactive with any of the fluorogenic substrates, as were the Veillonella spe-cies. It has been recommended that genetically defined phy-logenies should be corroborated by other characterization and taxonomic methods. The present study indicated good agree-ment between enzymatic patterns assayed by fluorogenic sub-strate tests and phylogenetic groupings for several groups of species. These tests may thus be useful as a rapid and conve-nient method of biochemical characterization in taxonomy studies.
Fluorogenic substrate tests were found to provide a sensitive and simple method for biochemical characterization that could presumptively identify to species level many subgingival iso-lates within 4 h. The method was ideal for rapidly obtaining presumptive identifications of isolates before confirmation of identifications by definitive methods, such as whole-cell pro-tein profiles by SDS-PAGE.
ACKNOWLEDGMENTS
We thank Mary Sheehan and Charles DeMartino for assistance with strain identifications.
This study was supported by grants DE 09513 and DE 10160 from the National Institutes for Health.
REFERENCES
1. Alcoforado, G. A., T. L. McKay, and J. Slots. 1987. Rapid method for detection of lactose fermenting oral microorganisms. Oral Microbiol. Im-munol. 2:35–38.
2. Bedi, G. S., and T. Williams. 1994. Purification and characterization of a collagen-degrading protease from Porphyromonas gingivalis. J. Biol. Chem.
269:599–606.
3. Beighton, D., J. M. Hardie, and R. A. Whiley. 1991. A scheme for the identification of viridans streptococci. J. Med. Microbiol. 35:367–372. 4. Brunn, B., Y. Ying, E. Kirkegaard, and W. Frederiksen. 1984. Phenotypic
differentiation of Cardiobacterium hominis, Kingella indologenes and CDC group EF-4. Eur. J. Clin. Microbiol. 3:230–235.
5. Collins, M. D., D. N. Love, J. Karjalainen, A. Kanervo, B. Forsblom, A.
Willems, S. Stubbs, E. Sarkiala, G. D. Bailey, D. I. Wigney, and H.
Jousim-ies-Somer.1994. Phylogenetic analysis of members of the genus
nas and description of Porphyromonas cangingivalis sp. nov. and Porphyromo-nas cansulci sp. nov. Int. J. Syst. Bacteriol. 44:674–679.
6. Dellinger, C. A., and L. V. Moore. 1986. Use of the RapID-ANA system to screen for enzyme activities that differ among species of bile-inhibited
Bac-teroides. J. Clin. Microbiol. 23:289–293.
7. Elliott, J. A., and R. R. Facklam. 1993. Identification of Leuconostoc spp. by analysis of soluble whole-cell protein patterns. J. Clin. Microbiol. 31:1030– 1033.
8. Heukeshoven, J., and R. Dernick. 1988. Increased sensitivity for Coomassie staining of sodium dodecyl sulfate-polyacrylamide gels using PhastSystem Development Unit. Electrophoresis 9:60–61.
9. Hinnebusch, C. J., D. M. Nikolai, and D. A. Bruckner. 1991. Comparison of API Rapid Strep, Baxter MicroScan Rapid Pos ID Panel, BBL Minitek Differential Identification system, IDS RapID STR system, and Vitek GPI to conventional biochemical tests for identification of viridans streptococci. Am. J. Clin. Pathol. 96:459–463.
10. Holdeman, L. V., and J. L. Johnson. 1982. Description of Bacteroides
loes-cheii sp. nov. and emendation of the descriptions of Bacteroides melanino-genicus (Oliver and Wherry) Roy and Kelly 1939 and Bacteroides denticola
Shah and Collins 1981. Int. J. Syst. Bacteriol. 32:399–409.
11. Johnson, J. L., and L. V. Holdeman. 1983. Bacteroides intermedius comb. nov., and descriptions of B. corporis sp. nov. and B. levii sp. nov. Int. J. Syst. Bacteriol. 33:15–25.
12. Johnson, J. L., L. V. Moore, B. Kaneko, and W. E. Moore. 1990. Actinomyces
georgiae sp. nov., Actinomyces gerencseriae sp. nov., designation of two
geno-species of Actinomyces naeslundii, and inclusion of A. naeslundii serotypes II and III and Actinomyces viscosus serotype II in A. naeslundii genospecies 2. Int. J. Syst. Bacteriol. 40:273–286.
13. Khwaja, K. J., P. Parish, M. J. Aldred, and W. G. Wade. 1990. Protein profiles of Capnocytophaga species. J. Appl. Bacteriol. 68:385–390. 14. Kilian, M. 1978. Rapid identification of Actinomycetaceae and related
bac-teria. J. Clin. Microbiol. 8:127–133.
15. Kilian, M., L. Mikkelsen, and J. Henrichsen. 1989. Taxonomic study of viridens streptococci: description of Streptococcus gordonii sp. nov. and emended description of Streptococcus sanguis (White and Niven 1946),
Strep-tococcus oralis (Bridge and Sneath 1982), and StrepStrep-tococcus mitis (Andrews
and Horder 1906). Int. J. Syst. Bacteriol. 39:471–484.
16. Kinder, S. A., K. S. Kornman, and S. C. Holt. 1989. Characterization of selected gram-negative oral microorganisms by SDS-PAGE. Oral Microbiol. Immunol. 4:52–56.
17. Lala, A., A. Amano, H. T. Sojar, S. J. Radel, and E. De Nardin. 1994.
Porphyromonas gingivalis trypsin-like protease: a possible natural ligand for
the neutrophil formyl peptide receptor. Biochem. Biophys. Res. Commun.
199:1489–1496.
18. Laughon, B. E., S. A. Syed, and W. J. Loesche. 1982. API ZYM system for identification of Bacteroides spp., Capnocytophaga spp., and spirochetes of oral origin. J. Clin. Microbiol. 15:97–102.
19. Love, D. N., G. D. Bailey, S. Collings, and D. A. Briscoe. 1992. Description of Porphyromonas circumdentaria sp. nov. and reassignment of Bacteroides
salivosus (Love, Johnson, Jones, and Calverley 1987) as Porphyromonas
(Shah and Collins 1988) salivosa comb. nov. Int. J. Syst. Bacteriol. 42:434– 438. (Errata, 42:660, 1992 and 43:630, 1993.)
20. Maiden, M. F., and A. Tanner. 1991. Identification of oral yeasts by poly-acrylamide gel electrophoresis. Oral Microbiol. Immunol. 6:187–190. 21. Maiden, M. F., A. Tanner, and W. E. Moore. 1992. Identification of
Seleno-monas species by whole-genomic DNA probes, sodium dodecyl
sulfate-polyacrylamide gel electrophoresis, biochemical tests and cellular fatty acid analysis. Oral Microbiol. Immunol. 7:7–13.
22. Manafi, M., and W. Kneifel. 1990. Rapid methods for differentiating gram-positive from gram-negative aerobic and facultative anaerobic bacteria. J. Appl. Bacteriol. 69:822–827.
23. Manafi, M., W. Kneifel, and S. Bascomb. 1991. Fluorogenic and chromoge-nic substrates used in bacterial diagnostics. Microbiol. Rev. 55:335–348. 24. Manafi, M., and M. L. Rotter. 1991. A new plate medium for rapid
pre-sumptive identification and differentiation of Enterobacteriaceae. Int. J. Food Microbiol. 14:127–134.
25. Millership, S. E., and S. V. Want. 1992. Whole-cell protein electrophoresis for typing Mycobacterium tuberculosis. J. Clin. Microbiol. 30:2784–2787. 26. Moncla, B. J., and P. Braham. 1989. Detection of sialidase (neuraminidase)
activity in Actinomyces species by using 29-(4-methylumbelliferyl)a-D -N-acetylneuraminic acid in a filter paper spot test. J. Clin. Microbiol. 27:182– 184.
27. Moncla, B. J., P. Braham, and S. L. Hillier. 1990. Sialidase (neuraminidase) activity among gram-negative anaerobic and capnophilic bacteria. J. Clin. Microbiol. 28:422–425.
28. Moncla, B. J., P. Braham, L. K. Rabe, and S. L. Hillier. 1991. Rapid presumptive identification of black-pigmented gram-negative anaerobic bac-teria by using 4-methylumbelliferone derivatives. J. Clin. Microbiol. 29:1955– 1958.
29. Moore, L. V., and W. E. C. Moore. 1994. Oribaculum catoniae gen. nov., sp. nov.; Catonella morbi gen. nov., sp. nov.; Hallella seregens gen. nov., sp. nov.;
on May 15, 2020 by guest
http://jcm.asm.org/
Johnsonella ignava gen. nov., sp. nov.; and Dialister pneumosintes gen. nov.,
comb. nov., nom. rev., anaerobic gram-negative bacilli from the human gingival crevice. Int. J. Syst. Bacteriol. 44:187–192.
30. Moore, L. V. H., J. L. Johnson, and W. E. C. Moore. 1994. Descriptions of
Prevotella tannerae sp. nov. and Prevotella enoeca sp. nov. from the human
gingival crevice and emendation of the description of Prevotella
zoogleofor-mans. Int. J. Syst. Bacteriol. 44:599–602.
31. Moore, W. E. C., D. E. Hash, L. V. Holdeman, and E. P. Cato. 1980. Polyacrylamide slab gel electrophoresis of soluble proteins for studies of bacterial floras. Appl. Environ. Microbiol. 39:900–907.
32. Paster, B. J., F. E. Dewhirst, I. Olsen, and G. J. Fraser. 1994. Phylogeny of
Bacteroides, Prevotella, and Porphyromonas spp. and related bacteria. J.
Bac-teriol. 176:725–732.
33. Pederson, E. D., J. W. Miller, S. Matheson, L. G. Simonson, D. E. Chadwick,
P. J. Covill, D. W. Turner, B. L. Lamberts, and H. E. Morton.1994. Trypsin-like activity levels of Treponema denticola and Porphyromonas gingivalis in adults with periodontitis. J. Clin. Periodontol. 21:519–525.
34. Pot, B., C. Hertel, W. Ludwig, P. Descheemaeker, K. Kersters, and K. H.
Schleifer.1993. Identification and classification of Lactobacillus acidophilus,
L. gasseri and L. johnsonii strains by SDS-PAGE and rRNA-targeted
oligo-nucleotide probe hybridization. J. Gen. Microbiol. 139:513–517.
35. Slayne, M. A., M. J. Aldred, and W. G. Wade. 1990. A rapid, semi-automated SDS-PAGE identification system for oral anaerobic bacteria. J. Appl. Bac-teriol. 68:391–395.
36. Slots, J. 1981. Enzymatic characterization of some oral and nonoral gram-negative bacteria with the API ZYM system. J. Clin. Microbiol. 14:288–294. 37. Slots, J. 1987. Detection of colonies of Bacteroides gingivalis by a rapid fluorescence assay for trypsin-like activity. Oral Microbiol. Immunol. 2:139– 141.
38. Tanner, A., M. F. J. Maiden, B. J. Paster, and F. E. Dewhirst. 1994. The impact of 16S ribosomal RNA phylogeny on the taxonomy of oral bacteria. Periodontology 2000 5:26–51.
39. Tanner, A. C. 1986. Characterization of Wolinella spp., Campylobacter
con-cisus, Bacteroides gracilis, and Eikenella corrodens by polyacrylamide gel
electrophoresis. J. Clin. Microbiol. 24:562–565.
40. Tanner, A. C., M. N. Strzempko, C. A. Belsky, and G. A. McKinley. 1985. API ZYM and API An-Ident reactions of fastidious oral gram-negative species. J. Clin. Microbiol. 22:333–335.
41. Tanner, A. C. R., M. A. Listgarten, J. L. Ebersole, and M. N. Strzempko. 1986. Bacteroides forsythus sp. nov., a slow growing fusiform Bacteroides sp. from the human oral cavity. Int. J. Syst. Bacteriol. 36:213–221.
42. Vandamme, P., E. Falsen, B. Pot, B. Hoste, K. Kersters, and J. De Ley. 1989. Identification of EF group 22 campylobacters from gastroenteritis cases as
Campylobacter concisus. J. Clin. Microbiol. 27:1775–1781.
43. Van Winkelhoff, A. J., M. Clement, and J. de Graaff. 1988. Rapid charac-terization of oral and nonoral pigmented Bacteroides species with the ATB Anaerobes ID system. J. Clin. Microbiol. 26:1063–1065.
44. Van Winkelhoff, A. J., T. J. van Steenbergen, N. Kippuw, and J. de Graaff. 1986. Enzymatic characterization of oral and non-oral black-pigmented
Bac-teroides species. Antonie Leeuwenhoek J. Microbiol. 52:163–171.
45. Whiley, R. A., H. Fraser, J. M. Hardie, and D. Beighton. 1990. Phenotypic differentiation of Streptococcus intermedius, Streptococcus constellatus, and
Streptococcus anginosus strains within the ‘‘Streptococcus milleri group.’’ J.
Clin. Microbiol. 28:1497–1501.
46. Whiley, R. A., H. Y. Fraser, C. W. Douglas, J. M. Hardie, A. M. Williams, and
M. D. Collins.1990. Streptococcus parasanguis sp. nov., an atypical viridans
Streptococcus from human clinical specimens. FEMS Microbiol. Lett. 56:
115–121.
47. Whiley, R. A., and J. M. Hardie. 1989. DNA-DNA hybridization studies and phenotypic characteristics of strains within the ‘‘Streptococcus milleri group.’’ J. Gen. Microbiol. 135:2623–2633.
48. Willems, A., and M. D. Collins. 1995. Reclassification of Oribaculum catoniae (Moore and Moore 1994) as Porphyromonas catoniae comb. nov. and emen-dation of the genus Porphyromonas. Int. J. Syst. Bacteriol. 45:578–581. 49. Willems, A., and M. D. Collins. 1995. Phylogenetic placement of Dialister
pneumosintes (formerly Bacteroides pneumosintes) within the Sporomusa
sub-branch of the Clostridium subphylum of the gram-positive bacteria. Int. J. Syst. Bacteriol. 45:403–405.