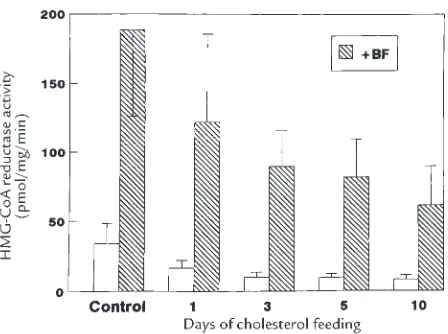
Increasing dietary cholesterol induces different regulation of classic and alternative bile acid synthesis
Full text
Figure
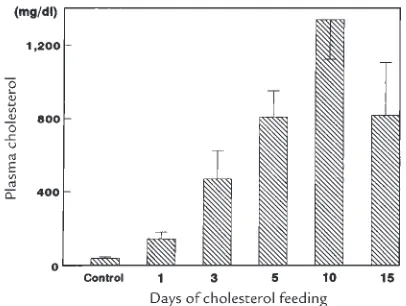
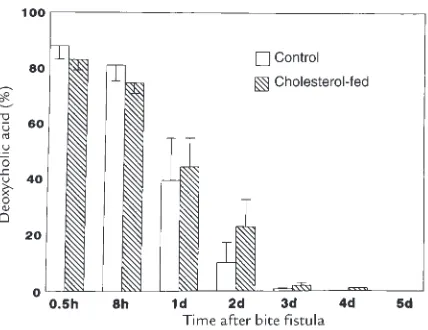
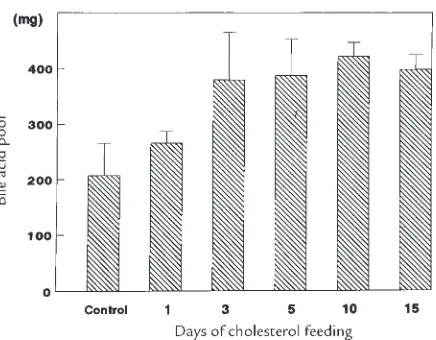
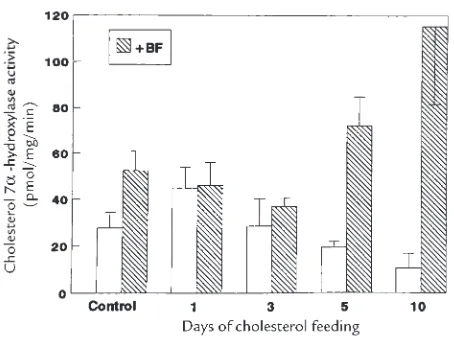
Related documents
The Coulomb stress τ C (α, q ) at the asperities on the sliding surface for arbitrary α (angle between the plumb line and the radius pointing from the axis of rotation to the cen-
This project is useful for the deaf and dumb, it can also be used for the (speechless) patients with half of their bodies paralyzed and who are not able to speak but
According to the framework, the independent variables were categorised into three main groups; starting from most distal factors: (i) community factors (place of residence,
Despite the FIELD-COMPLEXITY of Dahmen method was lower than that for classical method, Dahmen method is not suitable for computing odd points using on-the-fly
An analysis encompassing two case studies conducted in forward and backward regions of Maharashtra (India) has shown deterioration in the financial health of central level
The Ad-hoc On-Demand Distance Vector (AODV) routing protocol builds on the DSDV algorithm, it is an on demand routing algorithm, but in comparison to DSR it is a net
