r---1
f,dian.!. Plant Pltysiol., Vol. 4, No. I, (N.S.) pp. 15- I 'J (Jan.-March, 1999)
CHLOROPHYLL AS AN INDICATOR OF LIGHT INTENSITY IN
ANDROGRAPHIS PANICULATA
PRATIMAKAPUR
Department oC Botany, Mahila Mahavidyalaya, Banaras Hindu University Varanasi- 221005
Received on 2 Jan, 1998, Revised on 10 Nov., 1998
SUMMARY
Impact 'f light intesity on Andrographis paniculata showed that increasing light reduces the chlorophyll content on unit leaf area, land area as well as per gram dry weight basis. Light and age had significant effect on the chlorophyll content. Both, chlorophyll a and b were significantly affected by light. A negative correlation was observed between light and chlorophyll alb ratio.
Key words: Andrographis, chlorophyll, light intensity.
INTRODUCTION
Light, \\hich predominantly originates as a radiant flux, is unique among the environmental factors as a driving variable and individual organisms may be affected by any one of its aspects of intensity, colour, duration and direction. Among these, intensity of light nevertheless is most critical variable influencing photosynthesis through its absorption by chlorophyll (Treshow, 1970).
As a characteristic of the family Acanthaceac, leaves of Andrographis paniculata also change colour from green to red and vice versa from time to time. The reddening begins with the onset of the flowering and is delayed by a vYeek in shaded plants. Keeping this in view, the experiments were conducted to understand the behaviour of the chlorophyll under different light conditions. A slight fluctuation in the chlorophyll is enough to trigger changes in physiological processes of the plants including photosynthesis. Thus, in dry matter production, chlorophyll plays one of the most basic role.
MATERIALS AND METHODS
given four light intensity treatment. One set of plants was kept in the open, without attenuating light and was considered as plants under 100% relative light intensity (RLI), while the other three sets were kept under 15. 30 and 45% of the total light by attenuating light by thatching. Chlorophyll content of leaves, stem and inflorescence was detennined in plants of different age following Amon ( 1949), and further estimation on unit dry weight basis was done according to Wood and Bachelord (1969) Quantitative estimations were done for chlorophyll on unit leaf area basis, per unit land area as well as its distribution pattern. Chlorophyll index ( C I) was calculated according to Singh ( 1974).
CI = Total chlorophyll Ground area
RESULTS AND DISCUSSION
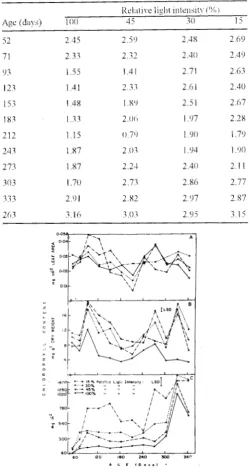
The detrimental effect of light intensity on the chlorophll content per unit leaf area (mg em-=) is evident from Fig. lA, and a negative correlation (r = -0.94) was also observed between the two With age, a rise was Andrographis paniculata (Burm. F.) Wall ex Nees., recorded up to 93 days in each of the four light regimes, a member of family Acantahaceae and a bitter shrub was followed by a fall up to 212 days. beyond ,,hich the trend
is not very clear. A negative correlation was obtained Table I
between chlorophyll (mg cm-2
) and NAR (r = -0.91) and
Variation in the chlorophyll a/b ratio of Andrographis panicu!ata with light and age.
SLW (r = -0.83).
Similar to chlorophyll per unit leaf area, the chlorophyll per unit dry weight (mg g·1
) also bore a
negative correlation (r = -0. 99) \\ith light. The value in this case too increased up to 93 days. where the first peak was recorded at all the light levels. A decline however. followed differently under different light condition. A second peak was recorded at 273 days in 15,30 and 100% RLI plants, while it was attained a month earlier at 243 days in the 45% exposure plants (Fig. 1 B). the values then declined and rose sharply again under shaded condition to yic:ld a third peak at 333 days coinciding with the peak of the second \Cgetative stage. The highest value of 19.68 mg g·1
\\as recorded by the 15'1';, RLI plants at 93 days.
Chlorophyll content (mg m2
) was signifncantly
;.lffected both by light as well as age of the plants. With ltght, chlorophyll followed a decreasing trend at almost all the ages and a significant relationship (r = -0. 90) was obtained between the tvvo. \Vith age. an increasing trend was obtained up to 123 days in the upper three light regimes \\bile at the 15% level it continued to increase up to 153 days. After a brief slack period, the values started rising again at 183 days in the 30 and 45°/r, and at the 243 and 7""'3 days in 15 and I 00% plants respectively. (Fig 1 C). The peak \vas obtained at 3 3 3 days, in each of the four light intensities.
Significant variation in the chlorophyll content was brought forth by its components a and b. Chlorophyll a/ b ratio showed a significant negative correlation (r =
-0. 95) with light. With age, the ratio declined up to 212 cl:1ys and then increased steadily (Table l)
Relative distribution of chlorophyll(% of total) in the component parts presented in Fig. 2. does not reveal any consistent trend with light. With advancing age, leaf showed a decline up to the second flowering stage. but reco\ ered thereupon (Fig. 2). Contrary to the leaves, stem showed an increasing trend of chlorophyll up to 153 days
16
RelatiYe light intem;it\ ('% 1
Age (days) !()() 45 30 15
52 245 2.5lJ 24R 2.69
71 2.33 2.32 240 2.49
93 1.55 141 2.71 2.G3
123 1.41 I ' ' ... ,'!,) 261 240
!53 1.48 I.R'! 2.51 2.67
1R3 133 2.W> 1.')7 2.2R
212 115 07') l 90 1.7')
243 JR7 2()3 1.94 1.90
273 l.R7 2.2c.l 240 2.11
303 1.70 2.73 2.86 2.77
333 2.91 2.R2 2.97 2.87
2(13 3.16 .un 2.95 3.1 ~
0·0'
-"
A~ 0·04 I ~,....·,. \ ...
__ ... -··;.~---
.•
~-···· .. ..._.r ., .'
I
.,
· . . --~---. I ' .-·-· ilr'"":..:_-:_· I ""'"\
~
-.... :1 --1 E" 0·01 -"-<\~-·-·-i/
\.J
60 t...6<L:o--::C:,2o:---,-:-,oo=--~240::---="""=----~,.:-::-'o
6 (, E: f 0 .. " . l •
Fig. I : Chlorphhyll content A · mg em' lear area, B . mg g-1 dry
\\eight, C : mg m·'land area in tour light regimes during gnmth period of A. paniculata.
in the p !ants exposed to 3 0'% RLI and up to 183 days in 15 and 100'/'o RLI plants while up to 212 days in 45% RLI plants. A decline after this persisted up to the end, except at 363 days where the values rose again with the luxuriant grovvth of rejuvenated plants. In case of inflorescence, an
CHLOROPHYLL AND LIGHT INTENSITY
RLI- 15% 30% 45% 100%
~8~y~f ~
I
~ ~ ~I
71 ~
J
g
E3 §93~ .~ ~ El
I
123~
I
§ ~ m ~•
153 t==:l
I
§g ..I
~•
~•
183
•
~•
~•
S
::::1
212~··· • • • ~
•
h=-1•
§Loof . •243 ~
I
~§I
§I
h=j ~~~~~~-1273 ~
I
8
s
I
~I
303 [ ~ [3 ] ~
333 ~ j
e
~. I
363 § . E3
I
El
EgjI
100°/o CHLOROPHYLL DISTRIBUTION PATTERN ("to OF TOTAL)
Fig. 2: Distribution of chlorophyll ('Yo of total) in photos\nthetic parts /1. paniculata under four l;ght kvcls
increasing trend was observed only up to I X3 days in the
mitial three intensities, after which the values dropped. Tn I 00%) RLT plants, the increase was seen up to !53 days
followed by a sharp fall and values picked up again from 183 days except for a sharp fall at 273 days (Fig. 2)
An inverse relation of CI \Vith light intensity \\as
observed at almost all the ages (Table II) a linear increase
''as obsen ed \\ith advancing age up to 93 days in all the
light regimes. The values remained steady up to 123 days
in the initial intensities and up to 183 days under full
exposure, followed by a fall and rise differently under
different light intensities. Mostly the values rose at lo\\er
lights The peak of CI \\as obtained at different ages by the different light plants.
Chlorophyll is the most imp011ant basis of dry matter
production on the earth. A h1gh level of dry matter
production is possible when chlorophyll is present in a significantly high amnunt. Reddening of leaves \\ith
increasing light coupled With a decline in total chlorophyll is evident from the negative correlation obtained bet\\een
chlorophyll and light intensity. Anthocyanian synthesis.
lnd1tmJ P/,1/il Phy.,1ol .. VoL 4. :\o.l. (1\.S.) pp. 15-19 (Jan.-1\larch. 199'))
which imparts colour to the leaves under full exposure ts
promoted by high light intensities moderatly lo\\
temperature and adequate supply of sugars (Salisbury
and Ross 1974 ). The lowering of temperature under
shaded condition, coupled \\ith a reduction in the
assimilation rate, hampering the supply may be a contributory to the reduced reddening of leaves and
consequently delayed senescence of the plants under lo\\ light intensities. Higher light intensities have a damaging
effect on the chlorophyll, both on weight as well as area
basis. This is substantiated by a sequential reduct10n in
chlorophyll content as the intensity of light increased, which was also reported by Naidu and Swamy ( 1993) and
Laing et ol. _ ( 1995). Pemmal and Rao ( 1991) also obsend
that a real reduction to 30% light, increases the chlorophyll
content in cotton plants. Decrease in the chloroph: II
content of the plants may be related to the fact that the
leaves mature rapidly as \veil as age and fall earlier under sunny conditions. Higher concentration of chloroph) II in
the herb layer under trees and shrubs were also shown by
Ovington and Lav\fence (196 7). who reported that the average chlorophyll content of the living material of the
Table II Effect of light and age on chlorophyll index
(CI-mg cm-2)
Relative lig_ht mtcnsitv ('Yr,)
Age (days) ]0() 45 311 15
52 0 006 0.006 0.0!3 () 014
71 () 014 0 017 ().(140 0 112X
93 ()()] 5 () ln'J () 1142 II (17}\ 123 () 015 0.0:\'J (I 042 11.117S
153 () 015 01125 OIUX () IIX5
IX3 I)()] :5 0 02:' 11.1140 0 115(,
212 0 Ill X 0 0 IX () 032 () 1159
243 0 ()]9 0 031 11113X () 1142
273 lUI I(, I) 025 11.053 () 125
303 (). 034 0 040 0.1155 1).(194
333 0. 10 I 0 1 c,x 0.234 II 1 XX
263 II()(,(, 0 072 ll21S 020X
herb layer in the open prairie was always lower than that under Oak canopies.
Light affects not only the total chlorophyll but also its
~onents as is evident from a negative correlation
between light and chlorophyll alb ratio, which was also observed by Argyroudi-Akoyunoglou and Akoyunoglou (1970) in young bean plants. This was also observed by Clayton (1966) when the hours of exposure were raised which he has attributed to the fom1ation of the chlorophyll containing only photosystem I. In addition to this the light and dark cycle seems to be an ideal for testing that chlorophyll is a precursor to chlorophyll b (Akoyunoglou
eta!., 1967) and for examining further, the possiblity that the conversion of chlorophyll a and b is photochemical rather than biochemical. This however needs further
investigation in A. paniculata. Contrary to this
Masarovicova and Eilas (1981) and Naidu and Swamy (1993) observed that shading decreases a/b ratio. Detrimental effect of light on the chlorophyll per unit leaf area is evident from the negative correlation between the two in A. paniculata. Contrary to this Kirita and Hozumu ( 1973) presented a hyperbolic curve between chlorophyll per unit leaf area and mean relative illumination, while a linear relationship was obtained by Okubo eta!., (1975 b). Sharma and Sen (1971) also observed maximum chlorophyll per unit leaf area in Solanum nigrum Linn. when grown under continuous illumination. Cooper and Qua lis (1969) observed that shading increases chlorophyll on unit dry weight basis but decreases on unit area basis. Negative association of SLW with chlorophyll per unit leaf area shows that light favours the accumulation of photosynthates, while it reduces the assimilatory unit. In
contrast to this Okubo eta!. (1975a) obtained a significant positive correlation between chlorophyll mg cm-2 and SL W. The relationship in the present investigation indicates that even though the light intensity inside the leaf must be reduced by the chloroplast pigments and even though the spectnun is alsC' changed in the direction of the wavelength of decreasing effectiveness, light was not enough to saturate chlorophyll accumulation at the sites available because under conditions in which leaf
111
thickness increased, the site of chlorophyll formation decreased (Friend 1961). The negative correlation between chlorophyll per unit leaf area and NAR of Androgmphis paniculata may be due to the fact that the leaves with high chlorophyll content do not photosynthesize more rapidly because they probably lack the enzymes or coenzymes which are the product of light reaction for reducing the available C0
2 (Salisbury and Ross 1974) Reverse results vvere obtained on the other hand by Okubo et of.. ( 1975 a, b)
ACKNOWLEDGEMENT
The author is greatly indebted to Dr. K. C. Misra, Rtd. Reader of Botany, Banaras Hindu University, Varanasi, for guidance throughout the study period. Financial assistance from Post-Graduate Institute of Medical Sciences is gratefully acknowledged.
REFERENCES
Akoyunoglou, G., Argvroudi-Akoyunoglou, J.I-L Michel-Wolwertz, M.R. and Sironval, C. (19G7). Chlorophyll a as a precursor for chlorophyll h synthesis in harelv leaves. Chimika Chronilw. 32A • 5-8.
Arg\'roudi-Akoyunoglou, .J .H. and Akoyunoglou, G. ( J 970). Photoinduced changes in the cl1lorophyll a to chlorophyll h ratio in young bean plants. Phmt Physiol., 46 • 247-249. Amon, D.!. ( 1949). Copper enzymes in isolated chloroplasts. Plant
PhysioL, 24 • 1-15.
Clayton. R.K. (1966). Physical process involving chlorophyll:-; in vivo
In thechlorophylls ( ed.) L.P. Vernon and G.R. Seelv,Academic Press, New York and London. pp. 609-641.
Copper, C.S. and Qualis, M. (1967) Morphology and chlorophyll content of shade and sun leaves oftwo legumes, Crop. Sci., 7• 672-673
Friend, D.J.C. ( 1961 ). The control of chlorophyll accumulation in leaves of Marquis wheat bv temperature and light intensity. II chlorophyll content relative to leaf area and thickness. I'hysiol. l'lanl., 14 • 28-39.
CHLOROPHYLL AND LIGHT INTENSITY
Kirita, H. and Hozumi, K. ( 1 973 ). Estimation of the total chlorophyll amount and its seasonal changes in a Warm-Temperate evergreen Oak forest at Minamata, Japan.!. Ecol., 23: 195-200.
Liang, N., Nagayama, M., Nakata, M. and Maruyama, K. (1995). Growth, photosynthesis and nitrogen content inj apanese beech
(Fagus crenata B.) Seedlings grown under live irradiance.
Photosynthetica, 31 : 257-268.
Masarovicova, E. and Stefanick, L. (1990). Some ecophysiological features in sun and shade leaves oftall beech trees. Biol. Plant., 32 : 374-387.
Naidu, C.V. and Swamy, P.M. (1993). Effect of shade on growth, biomass production and associated physiological parameters in
Pongamia pinnata (Linn.). Pierre. Indian.!. Plant Physiol.,
36 :212-214.
Okubo, T., Kawanabe, S and Hoshino, M. (1975 a). Chlorophyll amount for analysis of matter production in forage crops. II Sessional variation in maximum crop growth rate and leaf photosynthesis, and their correlation with chlorophyll content inalfalfaandladinoclover..J. Japan. Grassl. Sci., 21 : 124 135.
Okubo. T., Kawanabe, S. and Hoshino, M. (1975 b). Chlorophyll amount for analysis of matter production in gorage crops. ill
lndianJ. Plant Physiol., Vol. 4, No.!, (N,S.) pp. 15-19 (Jan.-March, 1999)
Leaf photosynthesis under dim light, maximum crop growth rate under dillcrent density and their dependence on chlorophyll content in alfalf~t..!. Japan. Grassl. Sci., 21 : 136-145.
Ovington, J.D. and Lawrence, D.B. ( J %7). Comparative chlorophyll and energy studies of prairie. savana, Oakwood and maize ecosystem. Ecology, 48 : 515-524.
Perumal. N.K. and R<w, M.R.K. (1991 ). Etrcct of low light on the growth and yield in cotton. Indian.!. Plant Physiol., 34 : 288-290.
Salisbury, F.B. and Ross, C (1974 ). Plant Physiology. Prentice/Hall of India Pvt. Ltd, New Delhi.
Sharma, K.D. and Sen, D.N. (1971 ). Photofraction in relation to grmvth behaviour in Solanum nignan Li1m. Oecol. Plan!.;
6 : 35-42.
Singh, M.K. ( 1974 ). Ecophysiology ofsixrice varieties under dry land !arming. Ph. D. thesis. Banaras Hindu University.
Treshow. M. ( 1970). Envirorunent and Plant Response. McGraw-Hill Book Company, N.Y.
Wood, J.P. and Bachelord, E.P. (1969). Variation in chlorophyll concentration in the foliage of Radiantpine. Aust. Forest1y,
33: 9-14.