Copyright 0 1982 by the Genetics Society of America
ASYMMETRIC EFFECTS O F DELETIONS AND SUBSTITUTIONS
ON HIGH NEGATIVE INTERFERENCE IN COLIPHAGE LAMBDA
G. J. VANCE MAKIN' A N D W. SZYBALSKI'
McArdIe Laboratory for Cancer Research, University of Wisconsin, Modison, Wisconsin 53706 F. R. BLATTNER
Deportment of Genetics, University of Wisconsin, Madison, Wisconsin 53706 Manuscript received June 24, 1980
Revised copy accepted August 5, 1982
ABSTRACT
Experiments have been performed to help clarify the role of nonhomologies in phage h recombination. Three-factor crosses were carried out, and the frequencies of single and double recombinants in the two adjoining intervals were compared when the central marker was either a double point mutation (vlv3) or deletion (rex-cl deletion) or nonhomologous substitution (imm434). In all cases the lefthand marker was a bio substitution (Fec- phenotype, which does not permit plating on recA-), and the righthand marker was a n amber mutation in gene 0. Experiments were performed in all four possible arrange- ments of the central and rightward markers, while selecting for the Fec' phenotype on the recA- host. As anticipated, high negative interference (HNI) was observed with point mutations, but when the central marker was a substi- tution nonhomology, HNI was reduced about tenfold. Surprisingly, when the central marker was a simple deletion, a dramatic asymmetry in results was observed, with HNI being exhibited only when the central deletion marker was acquired by the double recombinant. These results indicate that under normal conditions (red', gam', ret+) and with noninhibited DNA replication, recombi- nation in coliphage h entails a highly asymmetric step that could be at the level of strand transfer or mismatch repair.
HE
mechanisms of general recombination have proved to he a particularly
T
intractable problem in molecular biology, although numerous possibilities
have been proposed
(MOSIG
1970;
RADDING
1973,
1978;
MESELSON
and
RADDING
1975;
BROKER
and
DOERMANN
1975;
HOTCHKISS
1974).
Most
of the proposed
mechanisms are of the break-and-rejoin type and may he subdivided into two
mechanistic types: those in which the exchange of genetic information occurs
through the transfer of a single-stranded portion of one parental chromosome
into the other, and those in which the genetic information,
as
duplex
D N A ,is
reassembled from the parental chromosomes into a recombinant chromosome.
Strand transfer has been postulated by several workers [see, for example,
Present address: Department of Clinical Neurological Sciences, University of Western Ontario, London. To whom reprint requests should be addressed.
Ontario, Canada N6G 2B2.
300
V. MAKIN, W. SZYBALSKI A N D F.R.
BLATTNERWHITEHOUSE
1963; HOLLIDAY
1964; Fox 1966; MESELSON
and RADDING
1975), and
has been shown to be a physical possibility by BROKER
and
LEHMAN
(1971),
SIGAL
and ALBERTS
(1972) and THOMPSON,
CAMIEN
and WARNER
(1976)l.
The two formal break-and-rejoin mechanisms (with and without strand
transfer) can be refined by considering the possible roles of isomerization of
crossed-strand exchanges (EMERSON
1969; SIGAL
and ALBERTS
1972; SOBELL
1973)
and of repair of mismatched base pairs by cellular repair functions. Thus, the
break-and-rejoin model could involve
(1)
isomerization of a crossed-strand
exchange, followed by cleavage of the crossed strands or by cleavage of the
uncrossed strands,
(2)
cleavage of the uncrossed strands in the absence of a n
isomerization (see e.g., BLATTNER et al. 1974a), or (3) by the less likely route of
reassembly of double strand fragments of parental DNA (see SZYBALSKI
1964;
THOMAS
1966). In the case of strand transfer models, observed patterns of
recombination would depend upon the length of the strand transfer, the pres-
ence of associated isomerization of crossed-strand intermediates and mismatch
repair of heterozygosities.
HIGH NEGATIVE INTERFERENCE I N LAMBDA
301
The second possible mode of strand transfer resulting in HNI is one producing
a heteroduplex region covering only the central marker of the classical three-
factor cross. Such a heteroduplex would segregate double recombinants upon
replication without requiring any associated mismatch repair (see, for example,
FOX
1966; BLATTNER
et
al. 1974a). An important prediction of such a model is
that HNI should operate mainly in the case of double recombinants, with the
frequency of triple and higher order recombinants not significantly higher than
the product of double and single recombination frequencies, unless there is
additional mismatch repair. This appears to be the case for recombination in
phage
h
carried out under wild-type (Red',
Reef,
Gam+, normal replication)
conditions
(J. D. BOREL, T. M. SHINNICK,
V.
MAKIN,
W. SZYBALSKI
and
F.
R.
BLATTNER,
unpublished data). This mechanism is not in conflict with the long
strand transfers observed by WHITE
and
Fox
(1974) under replication-deficient
conditions since, as recognized by them, short insertion heteroduplexes may
simply not mature into phage particles under their conditions (see also STAHL
and STAHL
1974).
It might be predicted that a strand transfer process, whether symmetric
(HOLLIDAY
1964) or asymmetric (MESELSON
and RADDING
1975; see also
DISCUS- SION),would be hindered by major genetic nonhomologies (BROKER
and LEHMAN
1971;
Fox,
DUDNEY
and SODERCREN
1979). To test this prediction, three-factor
crosses have been performed in which the central marker could be either a
point mutation, simple deletion
or
substitution nonhomology. This was done
with all possible orientations of markers in order to detect any possible coop-
erative effects (positive
or
negative) of particular marker combinations upon
recombination
or
subsequent recombinant recovery. As hypothesized, the con-
siderable abundance of double recombinants observed with the point mutation
crosses was reduced by an average of tenfold when the substitution nonhomol-
ogy served as the central marker. In the case of the simple deletion, a dramatic
asymmetry in results was observed, HNI being exhibited by only one subset of
marker orientations.
All of the experiments reported here have been carried out under conditions
in which the Red system of
h
is operating and the Rec system of
E. coli is
partially inhibited by the Gam function of
h
(KARU et al. 1974). In addition,
replication is normal so that there is no major constraint upon emerging phages
to be in some way recombinant in order to be maturable (STAHL
and STAHL
1974; SKALKA
1974), nor is there any reason to suppose that the error-prone
recombination enzymes (SOS system) have been activated (WITKIN
1976).
MATERIALS AND METHODS
Media a n d buffers: NZB/Mg medium consists of 10 g/liter N-Z-amine A (Humko-Sheffield), 5 g/
liter NaCl and 0.01 M MgS04. Tris-MgS04 (TMS) buffer is 0.1 M Tris HCl, 0.1 M NaCl and 0.01 M
Plastic Petri plates, containing 40 ml of a mixture of 10 g/liter N-Z-amine A, 11 g/liter Bacto-Agar (Difco Laboratories) and 5 g/liter NaCl, were well dried before use. Top agar was a mixture of 10 g/ liter N-Z-amine A, 6.5 g/liter Bacto-agar and 5 g/liter NaCl.
302
V . MAKIN, W . SZYBALSKI A N D F. R . BLATTNERE. coli Ymel, grown in NZB/Mg. Phages with turbid phenotype were treated identically except that three to five plaques were suspendcd in 1 ml chloroform-saturated TMS and allowed to stand a few minutes at room temperature; the chloroform was driven off in a stream of filtered air and 0.1 ml of the resulting suspension was used to infect 0.1 ml of cells. In each case, 15-min incubation at 37' w a s followed by the addition of 10 ml NZB/Mg, and the culture w a s shaken at 37" until lysis was complete. Chloroform was then added and bacterial debris removed by centrifugation. In the case of the XcZ'nin5 used here, which grows poorly, a secondary lysate w a s prepared by infecting 10 ml of stationary phase Ymel cells in NZB/Mg with the primary lysate, at multiplicity of infection (m.0.i.) of IO-", incubating 15 min at 37", and diluting into 1 liter of NZB/Mg for further growth at 37" with shaking until lysis was complete. After removal of debris, the phage were sedimented by 3 h r centrifugation at 18,000 rpm in a Beckman rotor 19 and resuspended in 5 ml TMS.
Plaque assays: Phage diluted in TMS were adsorbed in 0.1 ml of stationary phage indicator cells and incubated for 15 min at 37". Top agar, 2.5 ml, at 50" was added and the suspension was gently vortexed and poured over bottom agar in a plastic Petri plate.
Construction of phage strains employed in crosses: The origin of bacteria and of phage strains employed in these crosses is specified in Table 1. Construction crosses were done as plate crosses (BLATTNER et al. 1974b) or in the same manner a s the crosses for quantitative analysis. Although only phages in which the N gene was deleted required the nin5 deletion for their growth (COURT and SATO 1969; FIANDT et al. 1971), we incorporated this deletion into all parental phages to ensure that both single and double recombinants from any one cross were uniform with respect to this marker. Crosses used in construction of the nin5 phages with all required arrangements of the three markers are given in Table 2. Structure of all strains was verified as follows (see also MAKIN 1976). Efficiency of plating for all Xbio phages was less than on recA- bacteria, for all hOam29 less
than on
homoimmune lysogens. The measured buoyant densities of the phages (Table 3) were in good agreement with predicted densities. Several of the phages were also checked by electron microscopy of appropriate DNA heteroduplexes (BLATTNER et al. 1974a; M. FIANDT, unpublished data).
Physical properties of the phages and positions of markers: guoyant densities of the phages, which served for the verification of their physical structure, are listed in Table 3. Positions of markers used in this study, expressed in %A units 1485 base pairs (bp)] measured from the left terminus, along with references, arc shown in Table 4.
Crosses for quantitative analysis: Phage particles, 5 X lo", of each parent, as calculated from the lowest of three independent titerings to ensure a minimum m.0.i. of 5, were adsorbed to 10' cells of late log phase Ymel cells growing in NZB/Mg, for 15 min at 37" in a total volume of 0.3-0.8 ml. NZB/Mg, 10 ml, was then added, and cultures were shaken at 37' in 125-1111 Erlenmeyer flasks for 75 min more, followed by addition of chloroform and storage at 4'. Detailed analysis of the crosses was completed within 5 wk.
Quantitative analysis of crosses 1-12: Total output was determined for each cross by plating o n Ymel cells (Table 5, column B). Parental (Table 5, columns C and D) and recombinant (Table 5, columns E and F) outputs were also determined by plating on appropriate bacteria wherever possible. Crosses 1-8 were plated on N100(+80am2 su3) to near-confluent lysis and incubated at least 18 hr at 37O to allow maximum visibility of turbid plaques. Clear plaques from crosses 1 and 5 were transferred by toothpick to lawns of N100(+80am2) and N100(@80am2 S U ~ ) , respectively. Appearance of a clear or sectored area of lysis on N100(+80am2) implies the cI- phages were O+. The validity of this assumption has been confirmed in reconstruction experiments. Turbid plaques from crosses 3 and 7 were similarly analyzed. Clear plaques from crosses 2 and 4, and turbid plaques from crosses 6 and 8, were analyzed similarly except for a n initial purification procedure designed to separate them from the confluent background of Fec+am+ phages. This was accom- plished by transferring the plaque by toothpick onto a lawn of N100(+80amZ su3) and then streaking out with a strip of paper; lawns were poured 24 hr previously and stored at 4" (this results in smaller plaques). Approximately six plaques per plate can be purified in this way. Crosses 9 and 11 were plated on N100(+80am2 su3)(XOam29) and N100(@8Dum2 su3)(himm434 Oam29), respectively, at a concentration sufficient to give about 25 plaques per plate, and these were then transferred by toothpick to N100(+80am2)(hOam29) followed by N100(@80amZ su3)(XOam29) in the case of cross 9 or N100(@80amZ)(Ximm434 Oam29) followed by N100(+80am2 su3)(Ximm434 Oam29) in the case of cross 11 Crosses 10 and 12 were plated on the appropriate indicators as for crosses 9 and 11, hut
HIGH NEGATIVE INTERFERENCE I N LAMBDA TABLE 1
Sources of bacterial, lysogenic and phage strains"
- Relevant phenotype
Bacteria and lysogens Ymel
W3350
su3, suppresses Oam29, not
su O
Pam3
NlOO
N100(+8Oam2) N100(+80am2 su3) N100(+3Oam2)(hOom29) N100(+80am2)(himm4340 N100(+80am2 su3)(hOam29) N100(+3Oam2 su3)(Ximm4340 Ymel(himm21 clts4 Pam3) CA266(Aimm434 Pam8O) W3350(h)
Bacteriophages" Ymel(h)(PZ) Ymel(himm434)(P2) C600(himm434)(P2) Ymel(P2)
NlOO(Ximm434) W3350(P2) AclAKH54 Pam80
am29)
am29)
hcZAKH54 nin5
Xb515b519bioKH341 c1857 Ximm21 clts4 nin5 himm434 cl- Oam29Pam3 hbioN2-1 imm434 nin5
hvl v3 Oam29 hbio30-7 Pam3 nin5 Xbio30-7 nin5
nin5'
suo, recA
sul, suppresses Oom29, not Pam80
su2, suppresses Oam29
rex-cl deletion
Sourceb and/or reference
303
J. S. SALSTROM J. S. SALSTROM;
J. S. SALSTROM; GOTTESMAN W. F. DOVE
W. F. DOVE This study This study
This study This study
CAMPBELL (1961) and YARMOLINSKY (1968)
J. S. SALSTROM This study
1.
S. SALSTROMThis study J. S. SALSTROM J. S. SALSTROM J. S. SALSTROM J. S. SALSTROM K. HASS; BLATTNER et al. K. HASS
K. HASS
(1974b)
J. S. SALSTROM
1.
S. SALSTROM J. S. SALSTROM J. BORELJ. S. SALSTROM J. S. SALSTROM
" All donors from McArdle Laboratory, University of Wisconsin, Madison, WI.
For phages constructed in this study see Tables 3 and 4.
A new Abio isolated a s hSpi- after induction of 594 (Ab515b519 xis6 cI857 nin5) lysogen. The bio cndpoint lies between Nam7 and Nom53 a s determined by marker rescue study, at 72.5 %A (see Table 4).
plaques were first purified on N100(+8Oam2 su3) by the paper strip technique and then toothpicked to N100(+80am2) followed by N100(+80om2 su3).
a
-
i i c .m
--
0 0
.- 0
5
c o c oE a
5 5
-
h h - hU %
a a e.a ""
g
s z
G Z
g
gggg
.U E E - . ? , ? N E N E E E E o,LN h
&
.? %.E
.E
E,s
.- U
s<S'zz
s 6 s ~ s g 4 L ~ g o
.
; a ~ ? 2 ~ ~ 0 T T ~ 0 0 0 0 a ; a ; a ; ~
a
~ ~ 3 u u z * * * u u u u ~ * * 3
i
&
a a a aI r0 C \ I W E E - V - w v w w -U c
.E
.E
4 4 r<-
VYVVD r W Q '
N r-a
&
g e E m < < s E E E $ $ $ $ E E E m
m
a m m m
C C C C m m m m c u m "
m 0 0 0 0
.e
.e
.E .EE E E E .5 0 0 0 0
C 1 1 1 1 m l 3 - - 3
C m ~ m m m m
c \ v c \ w w w v
- m $ E $ E E E
E
'3
c a ' =
v_-
w V w u v u w Ue e
U h
$
g 5
a
;
;
.E
;
;
.?
.E
.E
.E
mm
+- n n w w w w w v w c m m m m m m m n n n n n .E m
.- m 0
z z z z z z z -
E5
x
I
;;'=a?
% L M % % % Z'2
C F; .2
.s
g
B B B B B B BE , e
0 %- 0 -
g!--
u l n a n n n n n n n n 0 . 5-
a0 ; 0e - n z z g m z z z z z z z & % o Cs , g - -
0o = : z z = : z z z . i ?
O n ,2
4 4 2 2 2 2 2 2 2 2 2 2 2 2 2 2 2
a;
% % % 8
8
.z
.e
C.e
C l o a %0 0 5 a o o c U E E m
P 2 O 0 O O a ' 2
w a m
k k k ! - . e ! -
U O O V C t i & o . ~ o o E o o ~- 2
e w e 0 R . Q Mw
z g X g s t \ "
" O ? ? r j :C wE wE vE wE ~~
~ z z z z m o o o
~E ~ E M E X Y ~ ~ ~ ~ m m m m m m m - c E mE : % %
E E L D L D L O a m m Q ZE T T
.- y1
2
m l
-3 8
4
2
m m a a ab e
Ua
.3 C .3 c
m c m Q v 3 c
5 %
g %
5 . E E-E
-
5 5 8 5
'2
E E m m m O5
.e .e
.e
Q C C G M
c c o
::
x
3
E . E . E . ~ ~ . E . s . E ~ B ~ ~ r ; ~ ~ ~
x ~ 4 4 4 4 4 x 4 x 4 4 4 x 4 4 ~
C
m
'2 a m .E
h l c I
i
a
2
2 %
lE
-
- 0 . . : ' 2 29
.-p
5
LD'2
.C.-
n
z
v u
;;s
w Z G - %
m z % $ z
5
U m % Y E y %
2
-g
-
h l cz z z g z z z
. i Z m
U u u w c
z z z z z z z . g o y y
m $ $ : %
z z z z z z z c p a
w
c m
.*
...
z
.E: c m .-. 2 . 5c . * z o $ a
E a 0 C2
5 2 %
s
z
g S . 5
~
~
~
~
~
y1 .-
m .E: w w n n - n n a m
;
g
; 0L L 0 . e
''
5
m.O
.O .O .O .O .O .OE
m mQ ~ Q ~ Qo bQ> ~
~ ~ ~ ~ E m m m m m m m m g m n m
a . - . - O E E 9 o . e = : = : = : = : = : = : = : n . r ! . o . o 0 0 c n n a a a a n s a a n
4 4 x 4 4 4 4 x 4 4 4 4 4 x 4 4 4
304
2 2
.s
g
i 2
m '&
- 5
s;
% = E E
a;;;
5 5
2's
5 5
22
E2
s 5
G 2
S $
g :
2s
$ C L
$ +
2 2 c : m %$ 2
5 3Z Z g
a m 0 r;
m C 3 m
m z $
5
~ E
g
Zg 3 5
-
S E C 0 - 5
*
g
+ 2 e
3
-
j:
.z
'$
ggz
E* ! j 5
$ C F . 2 hz
2 . 5 -c: ; 2
a;e,,
2
E 3 Z
2
igz6g
. : : o a m' Z , 2 % 2
V Z 3 3
r O 2 - g g
3 2 : ; ;
U 0
I)
3 0
+ o
0 2
w - . w o n
-
vJ
.e
Z E
a,
E 5
Y J W
O h
m a ;
a $
E a ;
r *
b a ;
a d
.--
%
zz
eU " O
J=
.-
* mil "C: e
L l G U 0
5
2
z
m z(a m , s z
a m o w w
+ & z m m
C O m = *
m Q
H I G H NEGATIVE INTERFERENCE IN LAMBDA
TABLE 3
Buoyant densities of bacteriophages
305
Buoyant density (g/cm") Phage
Anin5 AOam29 nin5 Av1v3 nin5 hvlv3 Oam29 nin5 AcIAKH54 nin5 hcIAKH54 Oam29 nin5 himm434 cI- nin5 himm434 cI- OamZ9 nin5 Abio30-7 nin5
hbio30-7 vlv3 nin5 Abio30-7 Oom29 nin5 hbio30-7 vlv3 OamZ9 nin5 hb515b519 bioKH341 nin5 hb515b519 bioKH341 Oam29 nin5 Ab525b519 bioKH341 clAKH54 nin5 hb515b519 bioKH341 cIAKH54 Oam29 nin5 Ab515b519 bioKH341 imm434 cl- nin5 Ab515b519 bioKH341 imm434 cl- Oom29 nin5
1.5002 1.5001 1.5000 1.5001 1.4944 1.4946 1.4969 1.4973 1.4855 1.4836 n.d. 1.4836 1.4967 1.4973 n.d. n.d. 1.4937 1.4939
~ ~
a Buoyant densities determined by analytical ultracentrifugation in CsCl gradient versus A+ (1.508
g/cm3) and Ab2 (1.491 g/cm3) (see SZYBALSKI and SZYBALSKI 1971). All of the phages used in crosses 1-12 are listed here. n.d. = not determined.
TABLE 4
Physical positions of markers a n d lengths of intervals
Marker Position Reference
bio30-7 (right end) bioKH341 (right end) vlv3 (point mutations) cIAKH54 (deletion ends) imm434 (substitution ends) Oam29 (point mutation)
77.7 BLATTNER et al. (1974a) 72.5 M. FIANDT, unpublished data 78.55"
74.1-78.4 BLATTNER et 01. (1974b) 73.5-79.1
81.15b
BLATTNER et 01. (1974b); MANIATIS et al. (1975)
WESTMORELAND, SZYBALSKI and RIS (1969); BLATTNER et BLATTNER et al. (1974a and unpublished data)
al. (197413)
Intervals, %A units
d! di d I -d2
bio markers imm marker bio to imm imm to Oom.29 bio to Oam29
bio30-7 vlv3
bioKH341 cIAKH54
bioKH341 imm434
0.85
1.6 1.0
2.60 2.75 2.05
3.45 4.35 3.05 Combination of electron microscopic and sequencing data. 1 %A = 485 bp.
Derived from genetic mapping data in relation to electron microscopic maps.
, 'b 0
.c
-
EE; y h q o m m P \ v @ l c ? q q
i i " r i + . -
2
:
. E 6 2?3a
- 1
b
z w 0 0 z ' k 4
ii
E x13
p
.E%,
e r c e ~ \ m ~ ) c o p \ m m m m o m o m m mq 9 l n q T q O O r q " . U ) N c q W q p ? v T ?
'6'
C i M O O e O r J N O O O r i 0 0 0 0 0 0 o ?-v) n .-
3
C
0-
.o
z
L
o .a.a
g g
Oo2 s d r i r i r i 3 3 . -
2 .-
-
W
-.-
%
Ir, ~p :
x x x x x xC
9
. - E O ? ? l p ? *.- E
B
r i + r i c o r o l n0)
;
E-
"0g g
"0g
q03 2 5 %
0: x x x x x x
g g m s E 2 g Z
x x x x--
0 . E 9 0 " . d : L " O c q c q ~ d : h e e e: m
.-
E 2 % % % % + r i d . -E Q
g
x x x x U U U+=
v19 9 v 0
m m m e.a "
U
a
U
3
E : r ( + + r i M . -
2
Y
.-
i ) @ Y N + r i 1 I @ ?
B h -0
B
"vi
.- U I
C E c b so -om0 -ar b
3 2 d i + + r i + m o o o o o o o
m u
g
x x x x x x
g % % S % 8 2 2
+o q r ' ? r ' ? I ' O b
z 3 e z % w k s
g
p \ r i m r i m mL r J L
a P
3
.5
E
+ h
39
2 2
$
.E p? O? 0! ". W c! c? r! q C9 U) ". b r'? '4 % p? 0s m m, e m m e m m e e m m m m e e ~ w m m
g2L
-
U
al w
5
-
M 5 0s s w e
e Q ' w w l n l n l n l n w w w eZ Z "
z z z z
""E n - , ,
0 E > > 0 U
.E
.E
y
T 2 2
z
z
';jz
.E
.z
.E.E
3 e$ . - m l n p \ m - r i w m e m w h m m 0 - w .- " 42
TT%L4;z
T T T T % W % %
- $ ' " E 2
U vi 3 a--
u Ue
-
- +-C
U U
c G 2
c:
c:
E 2 -8
9 -
D
.o
2
.m
2
. ? E
rijU L l C
-
3 E
C
5
-
2 ;1p
2 2
U
-
Z
D
-
$ 2
a l a C GTi
;Ox= E $
5
,"z
,vixj
.-
o c,"
= E $
3
g e
.o
42 L O
-
m 2$ 5
5
z g
$ 5
2 9
- 3a 3 2 -03
2
s E $
z g 33 2
5
u -5
U,mI
v) c
2
c C U c nU Y ,
- 1 a
c -
i - w
U 0 0
a l o
5 5
-
, L ; m
- G - I n
- 0
0 T 4 c
306
A C 58.5.; C
5
5"
Am c E g g S x D
U:gz,,m
c 35
v m C Q X8
5
2gz
? ? E , . ,
- E c s z &
, : s
m: g c :
g a
> + 2 U;?:" 2 2 & E . 5 z z : Q $ c E
S
p.5
E$%.,S"a; 3p c z g c : 5 z ; 2 z
o m c o wg m
ci;g g 2 ;
: z g
Z4+&"_h.s
E
m 2
Z S Z U m zg
3z
c 3
g ;
22j
:Qz
3
&kg$;
$ & g g
g
U - 0 - c + . ~ W g Y m
g
a l o " " E G , m E ; 2
> -
g c2':
a2
2 2 %
c e
L O E 2
2.2
UU " z g , g p ~ ; E E o
o c gv
z:
c 2 2 2 z z
$3
G;.;E5
m s tE
2 8 ' ; ; s
$+-&% z + g e 3 - 2 . ; 2 2
;;
g n
U i - a l D - -;= c
a 5 2 -
;;;
z",srjz=-Oq:zhl
m u z:;
.&
$ 2
&Z+EE-
E, a a h m D g E : $ $ w
&
E - 0c!j
:;
2
:ay1%m4- qg,.&,Oe:CCd al U
> S I
E
oa
-
2
m
,
e : . - Z
32.f, E,
1
5 -
8
& m u'2.m.'.
" C g . s & e z
g
z -
32 -
" 3 - 02 2 . 5
2
2 Z . F
& c $ " o - - G $ E 2 , 5" " 2 5 W 0
3 Q e : V C W Z 2 - m m.5
s ' - z z . -
g-cu
,
G E:,"
33
% Z S C A 2& g
g
e 2 s
s
:s::
g
2.5u O ~ , ~ ~ - D Z ~ B ~
E-:
m g 0g
8
g
M z
0""-;O" U m
2 3
LOM12
E $;
$ 3
$ 2
m b w a %'"C " + - & b m c 0 %
2 . s
% -g - c
Ln
"
c:
g;
0e:A2
h U sc c 0 6. m . 2
k w u D
s g ~ g ~ ~ ~ 3 ~ 2 $ f 3
c
5 g g m
$ Zc y - 2 2
. - g m
m 22
g z g :
5
.
gz",s-&.u
a"; 2 U 0al
9
e C L 0 2 - o . 5 . 2gz;?
W " 3 2
.s$;cw;
m2
3gZS;:E
m E O , Es % e g 2 : o U U
U - Q O E ~ L Oc m " 3 g 2 0 m + . G g -
3w-32
~ . z G c . i $ 2
c:
-
C . 5 m . g -m
5
8
s , $ z g g +
0 0.C 2 z . s E . C " r , o mU L - 0
a 3
- U
ri c E C ~ m ~U o z C
-
. AzE
G.-v ) s z $ z
E g h So m w c - 0 E G t U Z
5
.e
e
0 0 % c $ z E O ?
2 ~ ? <
C E m + v L Or+ cr, 74"
0 a =
-Q;:w$p+$:
O " w c o ~ m E 3 s m g
" m m s c w
E E % : . S ~ ~ ~ C v j a i -I 3 3 al..-D E-:"
' 2
2LO W m 00GZ
.zm
sz
M Z 5
m w LO2%
E2
n
.e
E ' " 5 -C2 - z
as ?
co.sD
-
- 0 . 9 "U g U * e + C 6 2 X Z Z
m 0 3 a l Z e m
--e:
"
0 m z h C , s E s8
HIGH NEGATIVE INTERFERENCE IN LAMBDA
307
recombinant plaques might introduce such a bias since these might tend to be preferentially of one of the 0 genotypes. Each cross w a s analyzed several times, and the results are combined in Table 6.
RESULTS
Design
of
crosses
Three-factor crosses were made to test the effect of various kinds of nonho-
mologies on the outcome of a recombinational event in bacteriophage
A.
The
marker arrangements for the crosses are represented in Figure
1.
The left marker
in each case was a bio deletion-substitution with the Fec- phenotype (absence
of growth on recA- hosts) due to the loss of the red and gam genes. The right
marker was the Oam29 point mutation. The central marker was either
a
point
mutation (actually a double point mutant v l v3) or a major genomic modifica-
tion, i.e., deletion or substitution. Different bio substitutions were used to keep
the intervals, denoted dl and dz, between the three markers reasonably constant
(Table
4).
This was done since previous studies of
AMATI
and
MESELSON
(1965)
indicated that the magnitude of the high negative interference effect varied with
the interval length. Thus, we were able to compare recombination frequencies
directly without correction for interval length, as is done in Figure
1
and Table
6.
Recombinants in the dl (bio-imm) interval were selected as Fec+ phages with
the desired immunity phenotype using indicator strains nonselective for the
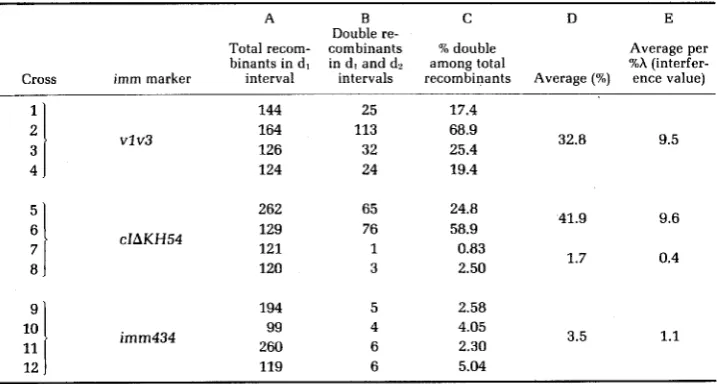
TABLE 6
Frequency of double recombinants among total recombinants in dl interval"
A B
c
n
ETotal recom- combinants % double Average per binants in d l in d , and dl among total Xh (interfer- Cross imm marker interval intervals recombinants Average (?A) ence value)
Double re-
1 2 3 4 .
7 8
!
11l l
12
144 164 126
vlv3
124
262 129 121
cIAKH54
120
194 99 260
imm434
119
25 113 32 24
65 76 1 3
17.4 68.9 25.4 19.4
32.8 9.5
24.8
58.9 41.9 9.6
1.7 0.4
0.83 2.50
2.58
1.1 4.05
2.30 5.04
3.5
V . MAKIN, W. SZYBALSKI A N D F. R. BLATTNER
308
FIGURE 1.-Diagrammatic representation of crosses used in this study. The left marker was always a bio DNA substitution (represented as an open rectangle) and the right marker was always a n OamZ9 point mutation. The central marker was varied to include a (double) point mutation v l
v3 (crosses 1-4), simple deletion cIAKH54 (crosses 5-8) and substitution of imm region of X with nonhomologous imm DNA of phage 434 represented as a rectangle (crosses 9-12). The outside heavy lines represent the parental phage genomes (all carrying the nin5 deletion) and the inside lines the single and double Fec+ recombinants. Each column includes all four possible orientations of the central and right markers. The "percent doubles" for each cross are taken from Table 6 , column C .
righthand markers. In crosses
1-8,this was done by plating the cross outputs on
recA- su3 bacteria and choosing the appropriate clear or turbid plaques. In
crosses 9-12, recA- su3 lysogens carrying either
X
or
Ximm434 prophages were
used as a lawn to select the appropriate immunity type among the Fec+
recombinants, and the plaques were subsequently purified on nonlysogens. In
all cases the procedure was nonselective with respect to the righthand (Oam29)
markers.
Recombinants in the dl interval were then scored for the 0 phenotype on
recA-
su Ocells to determine whether a second crossover had occurred in the da
interval. The percentage of double recombinants occurring in the populations
selected for at least a single crossover in dl is summarized in Figure
1.
Results
ofthe crosses
The results of 12 experiments numbered a s in Figure
1
are shown in Tables
5and 6. All crosses were done on the same day using the same batch of YmeJ
cells infected with each parent at a m.0.i. of approximately
5.
HIGH NEGATIVE INTERFERENCE IN LAMBDA
309
frequencies in this interval, i.e., an idiosyncratic increase (“hot spot”)
or
de-
crease. This is of importance for analysis of data derived from Table 6.
The frequencies of crossover per
%A
length in the selected (dl) interval (Table
5, column G) tended to be somewhat lower when the interval was bounded by
two major nonhomologies. However, this lesser degree of uniformity should not
affect the conclusions derived from Table 6, because this was not the interval
in which the second (unselected) recombination event was scored.
Analysis
ofcrosses with respect
to
high negative interference: The results
shown in Table 6 show a very high excess of double recombinants when the
point mutations vlv3 serve as the central marker (columns
C
and
D,
crosses
1-
4). For example, in cross
2
more than half (68.9%) of the recombinants in inteval
dl recombined also in interval dz, which is more than 100 times the expected
frequency of single crossovers in interval dz [equal to about 0.6%,3 derived as
the product of the interval length
(2.6
%A,
Table 4) and recombination frequency
per
%A
of 2.17
Xlow3,
Table 5, column
H,
line
2, and expressed as a percent-
age]. When the central marker is a substitution nonhomology, the double
recombinant frequency is dramatically lowered by factors up to
30
with an
average of ten (Table 6, columns
C and
D,
crosses 1-4 vs. 9-12). When the
central marker is a deletion, the recombination frequencies fall into two classes:
one resembling the point mutant crosses
1-4,
in cases in which the deletion
is
on the same strand as the bio substitution (crosses 5 and 6), and the other
resembling the
imm434
substitution (crosses 9-12), in cases in which the deletion
is on the Fec+ strand (crosses 7 and
8).
To
normalize the data to the total interval length (as to generate the so-called
interference value), the values in column
D
were divided by the physical
distance dl
+
dz and are listed in column
E. These interference values are
included mainly for historical interest, so as to permit a comparison with the
results of
AMATI
and MESELSON
(1965). However, we divided the data in column
3 by the total interval dl
+
d2 rather than by the measured single recombination
frequency in the interval d2, as was done by AMATI
and
MESELSON
(1965). Since
recombination rates are generally proportional to distance, our values in column
E should be proportional to their interference index.
As
stated before, our
unadjusted data in columns
C
and
D can be used directly, since the interval
lengths d l and dz were chosen to be approximately constant (Table
4).
DISCUSSION
The principal result of this study is the observation that major nonhomologies
within a recombining region exert a profound effect on HNI, measured as the
excess of double recombinants in the three-factor crosses. In those cases when
the central marker was a point mutation, about one-third of the preselected
” T h e value 0.65: is based on a model in which the second recombination event is independent of the first. The first crossover requires two chromosomes to enter a mating pool and subsequently undergo the necessary molecular events, whereas the second crossover requires only the latter factor. MAKIN (1976) has estimated that the probability of entering the mating pool is about 15% in this experiment, assuming that frequencies in crosses
310
V. MAKIN, W. SZYBALSKI A N DF. R.
BLATTNERsingle recombinants in region d l were also recombinant in the dz region (Table
6, crosses 1-4, column D), whereas the frequency of such double recombinants
was about tenfold lower in cases in which the central marker was a substitution
(crosses 9-12, column D). This result is in agreement with the simple prediction
of the strand transfer models, already discussed. Thus, w e believe that recom-
bination in phage
X
(under wild-type recombination and replication conditions:
Red’, Gam+, Rec+) proceeds predominantly by a strand transfer mechanism.
However, in crosses in which the central marker is a simple deletion, we
observed a profound asymmetry, with crosses
5and 6 yielding more than a
20
times higher percentage of double recombinants than crosses 7 and
8
(Table 6,
column D). This result might relate to the seemingly opposite results of DOERMAN
and PARMA
(1967), who observed no affect of central deletion on the recombi-
nation frequency, and of
BERGER
and WARREN
(1969), who recorded a depression
in recombination frequency in the case of central deletion (see also the discus-
sion of these results by MOSIG
1970).
This asymmetry observed for the
cI
deletion should permit a more detailed
examination of various strand transfer mechanisms of recombination. As shown
in Figure
2,
there are two main modes of strand transfer, symmetric (a) and
asymmetric (b). The present asymmetric results could be explained by asym-
metric strand transfer
or by mismatch repair. If such enzymatic repair of
heteroduplex intermediates formed by strand transfer were itself asymmetric,
both symmetric and asymmetric models could explain our results. However,
we shall first discuss only the asymmetric models and determine which of these
could explain our data without invoking mismatch repair. Subsequently, we
will also consider mismatch repair as a n auxiliary mechanism.
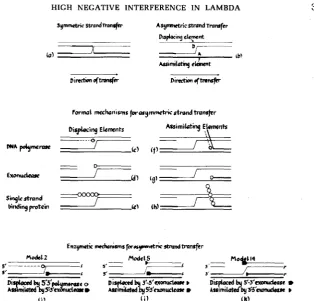
A general formulation of all asymmetric models of strand transfer is depicted
in Figure
2.
Such models (2b) imply a mechanism of displacement and assimi-
lation of one strand in any of several possible enzymatic ways. Displacement,
as mediated by element D, must occur in general by breaking the hydrogen
bonds between the strand to be displaced and its complementary strand. This
could occur either by replacing the displaced strand with one newly synthesized
by DNA polymerase (Zc), by removing the partner strand altogether with a n
exonuclease (2d), or by tying it up with a single strand DNA-binding protein
(Ze), such as the coliphage T4 gene 32 protein (ALBERTS
and FREY
1970),or one
of the DNA helicases
(KUHN, ABDUL-MONEM
and HOFFMANN-BERLING
1979).
Similarly, either polymerase, exonuclease or a single-strand-binding protein
could remove the appropriate strand on the recipient parent to allow assimila-
tion as shown in Figure Zf, g and h. As there are two strands, two strand
polarities and three possible displacing or assimilating elements, it is possible to
propose
2
X32 formal models for asymmetric strand transfer, a s listed in Table
7.
Note that all of these models would result in nonreciprocal recombination, a s
has been shown to be the case for the
X
Red system
(JACOBand WOLLMAN
1954;
SARTHY
and MESELSON
1976).
HIGH NEGATIVE INTERFERENCE IN LAMBDA
symmetric swondmnsfer AsymMn'cnmnd Transfer
bplacing Jcmnt D
A
( 0 )
x
tb)Mmimi r&t
Dimtbnoftrwofcr D;rutbn
+"
311
FIGURE Z.-Strand transfer mechanisms for genetic recombination. (a), Symmetric strand trans- fer according to HOLLIDAY (1964) and SIGAL and ALBERTS (1972). (b), Asymmetric strand transfer a s proposed by MESELSON and RAnDrNC (1975) requiring displacing (D) a n d assimilating (A) elements. Diagrams c-h illustrate all possible roles of the DNA polymerases, exonucleases and single strand DNA-binding proteins (SSDBP) acting a s displacing c, d and e or assimilating f , g and h elements, with DNA strand polarities not specified. Three specific examples, which correspond to models 2,
5 and 14 (see Table 7) with DNA strand polarities indicated, are represented in diagrams i, j and k. Note that in models 2 and 5 the 5' end of the DNA strand is displaced (5' + 5' overlap) and in model 14 the 3' end is displaced (3' + 3' overlap; see Table 7).
in
h
recombination, albeit under nonreplicating conditions, the inserted strand
of DNA begins with a 3'
end (3'
+
3'
overlap: see e.g., Figure 2k). As a
3'
+=5'
DNA polymerase most probably does not exist
(KORNBERG
1974), models 10,
11,
12, 13 and 16 are unlikely. Of the remaining models, only models 14 and 15
utilize an exonuclease of the same polarity as
X
exonuclease
(LITTLE
1967;
RADDING 1969), which is known to be involved in Red-mediated recombination
(SIGNER
1971). Model 15 requires a hitherto undiscovered single strand binding
protein of
3'
-+
5'polarity, whereas known examples are either nonpolar (phage
T4 gene
32
protein) or migrate in a 5'
-+
3'
direction (DNA helicases; KUHN,
ABDUL-MONEM
and HOFFMANN-BERLING
1979). On this basis, we favor model
14312
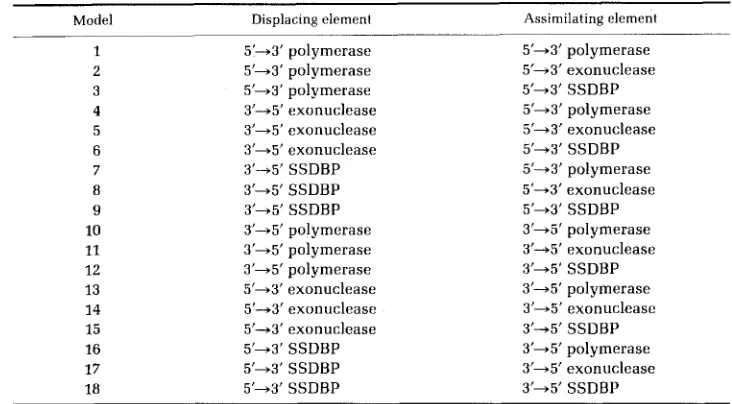
V. M A K I N , W. SZYBALSKI A N D F. R. BLATTNER TABLE 7Enzyme combinations capable of driving an asymmetric strand transfer reaction“
Model Displacing element Assimilating element
1 5’+3’ polymerase 5’+3’ polymerase
2 5 i 3 ’ polymerase 5 1 3 ’ exonuclease
3 5’+3’ polymerase 5 1 3 ’ SSDBP
4 3’+5’ exonuclease 5’+3’ polymerase
5 3’+5’ exonuclease 5’+3’ exonuclease
6 3’+5’ exonuclease 5 1 3 ’ SSDBP
7 3’+5’ SSDBP 5’43’ polymerase
8 3’+5’ SSDBP 5’+3’ exonuclease
9 3’+ 5’ SSDB P 5’+3’ SSDBP
10 3’+5’ polymerase 3 1 5 ’ polymerase
11 12 13 14 15 16 17
18
3’+5’ polymerase 3’+5’ polymerase 5’+3’ exonuclease 5’-+3’ exonuclease 5 1 3 ’ exonuclease 5 i 3 ’ SSDBP 5’+3’ SSDBP 5’+3’ SSDBP
3’45’ exonuclease 3’+5’ SSDBP 3’+5’ polymerase 3’+5’ exonuclease 3 ’ 4 5’ S S D BP 3’+5’ polymerase 3 1 5 ’ exonuclease
3 1 5’ SSDBP
Displacing and assimilating elements are diagrammatically represented in Figure 2, where also the specific model 2 (attributed to CASSIJTO and RADDING by SICAL and ALBERTS 1972), model 5 and model 14 (favored by this study; see Figure 3) are shown. The symbol SSDBP represents a single strand DNA-binding protein, e.g., the product of phage T4 gene 32 (ALBERTS and FREY 1970) or one of the DNA helicases (KUHN, ABDUL-MONEM and HOFFMAN-BERLING 1979); the latter could be considered as the 5’+3’ SSDBP. Note that models 9 and 18 are equivalent if the SSDBP exhibits no polarity. In models 1-9, a 5’ end of the DNA strand is displaced (5’45’ overlap), and in models 10- 18, a 3’ end is displaced (3’+3’ overlap, suggested by the results of WHITE and Fox 1974).
protein, then model 15 may represent the Red-mediated mechanism. Alterna-
tively,
bet
protein may act a s a cofactor of the host 3’
+5’
exonuclease or of
h
exonuclease of model 14 or of a host DNA-binding protein.
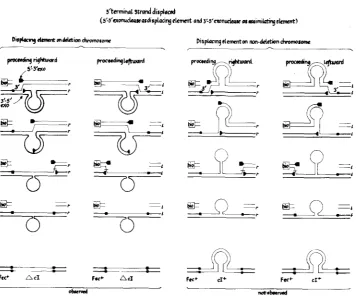
Figure 3
shows in detail several stages in the proposed mechanism of model
14 as it applies to the crosses involving the deletion nonhomology (see Figure
1,
crosses 5-8). The first two columns represent the crosses productive of double
recombinants (crosses
5and
6)
and the last two columns the nonproductive
crosses (crosses
7and 8). It is possible to rationalize this asymmetry in several
ways (MAKIN
1976), e.g., by a n inability of the 3’-
5’
exonuclease to wait at the
base of the
c l +
loop (Figure 3, columns
3and
4,line
2)for the 5’-
3‘ exonuclease
to complete its excursion, while the opposite combination is functional (columns
1
and 2, line 2).
The asymmetry observed in crosses 5-8 could also be due to destruction of
heteroduplex intermediates of the type shown in Figure
3(fourth line) in spite
of their successful formation. Such intermediates could be formed either by
symmetric or asymmetric strand transfer process. If the
c l +
single strand loops
of such heteroduplexes were enzymatically converted to the deletion genotype,
then the excess of double recombinants in crosses
5and
6(in relation to crosses
1-4)
and corresponding deficiency in crosses
7and 8 (relative to crosses
9-12)HIGH NEGATIVE INTERFERENCE IN LAMBDA
3't"inat strand d i r p h d
(s'-~'~onuc(uucuedispla~th~ element and ~'-5'nmnudavc os arimilatity &me)
--
---
F u + Ad
313
Dirpbcingdemcntm rm-dactiondxomoronw
,-.
.
c)'
'.
r
FIGURE 3.-Detailed mechanism of model 14 (5' + 3'-exonuclease as displacing element and 3' + 5'-exonuclease as assimilating element; see Table 7 and Figure 2) for crosses 5-8 as represented in Figure 1. The first and second columns (displacing element on deletion chromosome) represent crosses 5 or 6, whereas the third and fourth columns represent crosses 7 or 8 (location of 0 - marker not specified). The orientations of strands 1 and r are 5' + 3' and 3' + 5', respectively. The DNA of the bio parent is represented by thin lines and of the Fec+ parent by thick lines. The top line of the figure represents an early stage of the transfer process with a 3' end of a DNA strand of the bio parent being transferred to the Fec+ parent, the joint being represented by the solid square. The second line shows the progression of the transfer across the deletion nonhomology with the loop representing the cIc DNA. The third line formally illustrates the termination of the transfer process, with the short horizontal slash representing the required endonucleolytic cleavage of the displaced strand. The fourth line shows the end products of the transfer process, with the inserted strand bonded by two solid squares. The bottom line shows the double recombinant chromosome segre- gated during the first round of DNA replication. Note that only the Fec+AcI double recombinants (bottom line of first two columns) were observed at high frequency (see crosses 5 and 6 in Figure 1
and Table 6, column C).
Table 6 (column D:
41.9%
doubles vs. 32.8%,
and
1.7%doubles vs. 3.5%). Such a
phenomenon would be consistent with the evidence supporting a role for
mismatch repair cited in the introduction. Although repair must remain a
mechanism to be considered, preliminary data to test this possibility under our
wild-type experimental conditions have tended to negate it (see
MAKIN
1976),
especially since no obvious asymmetry was observed for the single crossovers
(ratio of values in columns
F
and E of Table
5
are
.140
and .167 for crosses 5 and
3 14
V. MAKIN, W. SZYBALSKI A N D F. R. BLATTNERobserved in crosses 5-8. Such deficiency, if seen in Table 5, might have indicated
a ‘‘repair’’-mediated
loss of the cI+ “loops” in the cI+/KH54 heteroduplexes, the
mechanism that was invoked by
BENZ
and
BERGER
(1973) to explain the loss of
the rII+ allele in crosses with rII deletion mutants of phage T4. Such repair in
T4 was observed only when
DNA
synthesis was reduced and the products of
T4 genes v and
x
were present. This is expected, since
v
and
x
are thought to be
required for repair.
If
there is no significant mismatch repair, as is considered likely, the asym-
metry in
HNI
is consistent with an asymmetric initiation of recombination. We
favor the asymmetric,
X
exonuclease-mediated strand transfer model described
by
BROKER
and
LEHMAN
(1971) (Figure Zk), as it successfully reconciles the 3’
+
3’ overlap observed by
WHITE
and Fox (1974), and the nonreciprocity for the
Red system observed by
JACOBand
WOLLMAN
(1954),
WOLLMAN
and
JACOB(1954) and
SARTHY
and
MESELSON
(1976), with known enzyme polarities. This
mechanism probably predominates under
our wild-type conditions when
A
replicates freely and the RecBC system of the host is suppressed by the
X
Gam
function
( K A R U ,ABDUL-MONEM
and
HOFFMAN-BERLING
1974). However, when
X
DNA
replication is artificially blocked, as is the case for most other analogous
published data (e.g.,
WHITE
and Fox 1974;
Russo
1973;
WEINSTOCK
1974;
WIL-
DENBERGand
MESELSON
1975) mismatch repair may be a predominant process,
especially since under such conditions the error-prone repair functions of the
SOS pathway are known to be induced
(WITKIN
1976). Moreover, the results of
Fox,
DUDNEY
and
SODERGREN
(1979; see also
MOSIG
et al.
1979) raise the question
of whether heteroduplexes across nonhomologies are generated by the same
mechanism as heteroduplexes involving extensive and perfect homologies. We
realize that several recombinational systems are operational in the
E.
coli-X
system, and depending on the conditions one or another of those seems to
predominate.
The experimental studies, which were carried out in years 1974-1976, were supported by the National Cancer Institute Program-Project grant CA-07175. W e would like to acknowledge the critical reading and comments of DRS. T. BROKER, A. H. DOERMANN, R. HOLLIDAY, C. M. RADDING, H.
M. SOBELL and F. W. STAHL. W e would also like to thank our colleagues J. S. SALSTROM, A. HONIGMAN, S.- L. Hu, E. ROSENVOLD and M. FIANDT for their help and advice. The excellent technical assistance of M. METZDORFF and K. HOLT is gratefully acknowledged.
LITERATURE CITED
ALBERTS, B. M. and L. FREY, 1970
AMAT;, P. and M. MESELSON, 1965
BENZ, W. C. and H. BERGER, 1973
BERGER, H. and A. J. WARREN, 1969
BLATTNER, F. R., J., D. BOREL, T. M. SHINNICK and W. SZYBALSKI, 1974a
T4 bacteriophage gene 32: a structural protein in the replication
Localized negative interference in bacteriophage A. Genetics
Selective allele loss in mixed infections with T4 bacteriophage.
Effects of deletion mutations on high negative interference in T4D bacteriophage. Genetics 63: 1-5.
Mapping of point mutations on the physical map of coliphage lambda: absence of clustering for odd-numbered exchanges. pp. 57-67. In: Mechanisms in Recombination. Edited by R. F. GRELL. Plenum Press, New York.
and recombination of DNA. Nature 227: 1313-1318.
51: 369-379.
HIGH NEGATIVE INTERFERENCE IN LAMBDA
315
BLATTNER, F. R., M. FIANDT, K. K. HASS, P. A. TWOSE and W. SZYBALSKI, 1974b Deletions and insertions in the immunity region of coliphage lambda: revised measurement of the promoter- startpoint distance. Virology 62: 458-471.
Molecular and genetic recombination of bacteriophage
T4. Annu. Rev. Genet. 9 213-244.
BROKER, T. R. and I. R. LEHMAN, 1971 Branched DNA molecules: intermediates in T 4 recombina- tion. J. Mol. Biol. 60: 131-139.
CAMPBELL, A., 1961
CHASE, M. and A. H. DOERMANN, 1958
COURT, D. and K. SATO, 1969
DOERMANN, A. H. and D. H. PARMA, 1967
EMERSON, S., 1969
FIANDT, M., Z. HRADECNA, H. A. LOZERON and W. SZYBALSKI, 1971
BROKER, T. R. and A. H. DOERMANN, 1975
Sensitive mutants of bacteriophage A. Virology 14: 22-32.
High negative interference over short segments of the
Studies of novel transducing variants of lambda: dispensability of
Recombination in bacteriophage T4. J. Cell. Physiol. 70
Linkage and recombination at the chromosomal level. pp. 267-360. In: Genetic Organization. Edited by E. W. CASPARI and A. R. RAVIN. Academic Press, New York.
Electron micrographic mapping of deletions, insertions, inversions, and homologies in the DNAs of coliphages lambda and phi80. pp. 329-354. In: The Bacteriophage Lambda. Edited by A. D. HERSHEY. Cold Spring Harbor Laboratory, Cold Spring Harbor, NY.
Gene conversion in unselected tetrads from multipoint crosses. pp. 89-110. In: Stadler Genetics Symposia, Vol. 1 and 2. Edited by G.
KIMBER and G. REDEI. University of Missouri, Columbia, MO.
On the mechanism of integration of transforming deoxyribonucleate. J. Gen. Physiol. 49 (Suppl.): 183-196.
Heteroduplex regions in unduplicated bacteriophage h recombinants. Cold Spring Harbor Symp. Quant. Biol. 4 3 999-1007.
Integration-negative mutants of bacteriophage lambda. J. Mol. Biol. 31: 487-505.
The combination of linkage values, and the calculation of distances between the loci of linked factors. J. Genet. 9 299-309.
genetic structure of bacteriophage T4. Genetics 43: 332-353.
genes
N
and Q. Virology 3 9 348-352.(Suppl. 1): 147-164.
FOGEL, S., D.
D.
HURST and R. K. MORTIMER, 1971FOX, M. S., 1966
FOX, M. S., C. S. DUDNEY and E. F. SODERGREN, 1979
GOTTESMAN, M. E. and M. B. YARMOLINSKY, 1968
HALDANE, J. B. S., 1919
HOLLIDAY, R., 1964
HOTCHKISS, R. D., 1974
JACOB, F. and E. -L. WOLLMAN, 1954
KARU, A., Y. SAKAKI, H. ECHOLS and S. LINN, 1974
A mechanism for gene conversion in fungi. Genet. Res. 5: 232-304.
Models of genetic recombination. Annu. Rev. Microbiol. 28: 445-468.
Etude gknetique d’un bacteriophage tempere d’Escherichia coli. Ann. Inst. Pasteur 8 7 653-673.
In vitro studies of the gam gene product of bacteriophage A. pp. 95-106. In: Mechanisms in Recombination. Edited by R. F. GRELL. Plenum Press, New York.
KORNBERG, A. 1974
KUHN, B., M. ABDUL-MONEM and H. HOFFMANN-BERLING, 1979
LITTLE, J. W., 1967
DNA synthesis. W. H. Freeman Company, San Francisco.
DNA helicases. Cold Spring Harbor
An exonuclease induced by bacteriophage A. 11. Nature of the enzymatic
MAKIN, G. J. V., 1976 Studies on the mechanism of Red-mediated recombination in bacteriophage
MANIATIS, T., M. PTASHNE, K. BACKMAN, D. KLEID, S. FLASHMAN, A. JEFFREY and
R.
MAUER, Recognition sequences of repressor and polymerase in the operators of bacteriophage Symp. Quant. Biol. 43: 63-67reaction.
1.
Biol. Chem. 242: 679-686.lambda. M. S. thesis, University of Wisconsin, Madison, WI.
1975
316
V. MAKIN, W . SZYBALSKI AND F. R. BLATTNERMESELSON, M. S. and C. M. RAnnrNc, 1975 A general model for genetic recombination. Proc. Natl.
Genetical interference and gene conversion. pp. 263-275. In: Acad. Sci. USA 72: 358-361.
MORTIMER, R. K. and S. FOGEL, 1974
Mechanisms in Recombination. Edited by R. F. GRELL. Plenum Press, New York. Mosrc, G., 1970
Mosrc, G., R. DANNENBERG, D. GHOSAL, A. LUDER,
s.
B E N E D ~ C T ands.
BOCK, 1979 Recombination in bacteriophage T4. Adv. Genet. 15: 1-53.General genetic
The genetic control of phage-induced enzymes. Annu. Rev. Genet. 3: 363-
Molecular mechanisms in genetic recombination. Annu. Rev. Genet. 7: 87-
Genetic recombination: strand transfer and mismatch repair. Annu. Rev.
On the physical structure of h recombinant DNA. Mol. Gen. Genet. 122
Single burst study of rec- and red-mediated recombination
Genetic recombination: the nature of a crossed strand-exchange
General recombination, pp. 139-174. In: The Bacteriophage Lambda. Edited by
A replicator’s view of recombination (and repair). pp. 421-432. In: Mechanisms
Symmetry in protein-nucleic acid interaction and its genetic implications. recombination in bacteriophage T4. Stadler Symp. 11: 31-56.
394.
RADDING, C. M., 1969
RADDING, C. M., 1973
RADDING, C. M., 1978
Russo, V. E. A., 1973
SARTHY, P. V. and M. MESELSON, 1976 111.
Biochem. 47: 847-880.
353-366.
in bacteriophage lambda. Proc. Natl. Acad. Sci. USA 73: 4613-4617.
between two homologous DNA molecules. J. Mol. Biol. 71: 789-793.
A. D. HERSHEY. Cold Spring Harbor Laboratory, New York.
in Recombination. Edited by R. F. GRELL. Plenum Press, New York. SIGAL, N. and B. ALBERTS, 1972
SIGNER, E., 1971
SKALKA, A., 1974
SOBELL, H. M., 1973 Adv. Genet. 17: 411-490.
STAHL, F. W. and M. M. STAHL, 1974 Red-mediated recombination in bacteriophage lambda. pp.
407-419. In: Mechanisms in Recombination. Edited by R. F. GRELL. Plenum Press, New York. Structural modifications of DNA: crosslinking, circularization and single- strand interruptions. Abhand. d. Deutsch. Akad. d. Wissenschaft. zu Berlin, Klasse Med., Nr. 4,
Equilibrium density gradient centrifugation. pp. 311- 354. In: Procedures in Nucleic Acid Research, Vol2. Edited by G. L. CANTONI and D. R. DAVIES. Harper and Row, New York.
THOMAS, C. A., JR., 1966 Recombination of DNA molecules. Prog. Nucleic Acid Res. Mal. Biol. 5:
THOMPSON, B. J., M. N. CAMIEN and R. C. WARNER, 1976 Kinetics of branch migration in double-
WA GNE R, R., JR. and M. MESELSON, 1976 Repair tracts in mismatched DNA heteroduplexes. Proc.
WEINSTOCK, E. J., 1974 Formation of bacteriophage lambda recombinants. Ph.D. thesis, Massa- chusetts Institute of Technology, Cambridge, MA.
WESTMORELAND, B. C., W. SZYBALSKI and H. RE, 1969 Mapping of deletions and substitutions in heteroduplex DNA molecules of bactcriophage lambda by electron microscopy. Science 163:
1343-1348.
On the molecular basis of high negative interference. Proc. Natl. SZYBALSKI, W., 1964
pp. 1-19.
SZYBALSKI, W. and E. H. SZYBALSKI, 1971
315-348.
stranded DNA. Proc. Natl. Acad. Sci. USA 73: 2299-2303.
Natl. Acad. Sci. USA 73: 4135-4139.
HIGH NEGATIVE INTERFERENCE IN LAMBDA
317
A theory of crossing-over by means of hybrid deoxyribonucleic acid.
Mismatch repair in heteroduplex DNA. Proc. Natl. Acad. WHITEHOUSE, H. L. K., 1963
Nature 199 1034-1040.
WILDENBERG,
1.
and M. MESELSON, 1975 Sci. USA 72: 2202-2206.WITKIN, E., 1976 Ultraviolet mutagenesis and inducible DNA repair in Escherichia coli. Bacteriol
Etude gCnCtique d'un bacteriophage temper6 d'Escherichia
Corresponding editor: G. MOSIG
Rev. 40: 869-907.
WOLLMAN, E. L. and F. JACOB, 1954