FOUR SIMULTANEOUSLY STABLE POLYMORPHIC
EQUILIBRIA I N TWO-LOCUS TWO-ALLELE MODELS
ALAN HASTINGS
Department of Mathematics and Division of Environmental Studies, University of Calqornia, Davis, Calqornia 95616
Manuscript received July 9, 1984 Revised copy accepted September 17, 1984
ABSTRACT
The existence of four simultaneously stable equilibria with both loci poly- morphic is shown for the Lewontin-Kojima version of the two-locus two-allele symmetric viability model, using bifurcation theory. This exceeds the previ- ously claimed bound of two stable polymorphisms. Biological implications of
the result are discussed.
NE of the methods used to determine whether a polymorphism is main-
0
tained by selection is to perturb the population under study and see whether it returns to its previous equilibrium. A failure to return to the pre- vious equilibrium is often taken as evidence that selection is not responsible for the maintenance of the polymorphism. Behind this reasoning is the sup- position that any polymorphic equilibrium maintained by selection is unique, which is true in single-locus models (KINGMAN 196 1). For two-locus two-allele models, KARLIN (1975) showed that with sufficiently small recombination rates at most two polymorphic (at both loci) equilibria can be simultaneously stable. He also conjectured that at most two polymorphic equilibria can be simulta- neously stable for any recombination rate (see also KARLIN and CARMELLI1975).
Additionally, on the basis of a detailed analysis of some of the equilibria possible, KARLIN (1975) and KARLIN and FELDMAN (1970) have claimed that at most two polymorphic equilibria can be simultaneously stable in the two- locus two-allele symmetric viability model (LEWONTIN and KOJIMA 1960). Re- cently, FELDMAN and LIBERMANN (1979) have shown that as many as four boundary equilibria and two polymorphic equilibria can be simultaneously sta- ble in this model.
In the current paper, I will show that, even in the special case of the sym- metric viability model first studied by LEWONTIN and KOJIMA (1960), four polymorphic equilibria can be simultaneously stable in a two-locus two-allele model. This suggests that extreme sensitivities to initial conditions are possible in two-locus models. I will use techniques based on bifurcation theory, as in
HASTINGS (1981, 1982).
MODEL AND ANALYSIS
T h e model studied here is the standard two-locus two-allele discrete time symmetric viability model (reviewed by KARLIN 1975; EWENS 1979). Let A and a be the alleles at the A locus and B and b be the alleles at the B locus. Let the frequencies (and “names”) of the four gametes AB, Ab, aB, ab be xl, x2, x3,
x 4 , respectively. Denote the viability of an individual with genotype xixj by w ~ ,
where wq = wj, and w14 = w23 = 1. Define the disequilibrium,
(1)
(2)
(3)
D = x 1 x 4
-
~ 2 x 3and the frequencies of the allele A ,
P A = XI
+
x 2 ,ps = X I
+
x3. and the allele B ,Finally, denote the recombination rate by r. T h e evolution of this system is described by
xi = (xiwi
+.
rD)/iij fori
= 1 to 4, (4)where the sign is minus for
i
= 1 and 4 and plus fori
=2
and 3. T h e marginal mean fitness of xi is4
(5) w . =
1
w . . x . Y .l’j = I
and the mean fitness of the population is
4
iij =
1
w j x j , j = IIn the Lewontin-Kojima version of the symmetric viability model one as- sumes that the fitnesses of all of the homozygotes are the same. So, let
w11 = wpp = w33 = w44 = 1 - ff.
(7)
(8)
(9) Additionally, it is assumed that
w12 = w34 = 1
-
p
and
~ 1= 3~ 2 = 4 1
-
7.Here a,
p,
y are assumed positive.This model has been extensively studied (KARLIN 1975; EWENS 19’79 provide reviews) and the following facts will be useful. There is always an equilibrium of the form
X I = x p = XQ = x 4 = %, (10)
IP
-
Y l’
(11)Additionally, if
r
<
( P
+
-
a)/4,there are two “symmetric” equilibria of the form
X ] = x4 = (1
+
4)/4
xp = x3 = (1
-
4)/4and
X ] = x4 = (1
-
4)/4x p = x3 = (1
+
4)/4
where
257
(12)
EWENS ( 1 968) showed that these equilibria are stable if and only if
7 ( r ) = 4 r 2 ( p
+
y-
a)+
2r(2a2-
P2
-
y 2-
a(p
+
7))(16)
+
a(@+
y-
a)’>
0. Finally, KARLIN and FELDMAN (1970) discovered the existence of as many as four further “unsymmetric” equilibria and provided formulas for them. They gave a detailed analysis, including a stability analysis, for the special caseP
= y. Under the conditions they analyzed, no more than two simultaneously stable equilibria are possible.Analysis of the model using bqurcation theory
In this section I, will describe a partial analysis of the unsymmetric equilibria in the case not studied in detail by KARLIN and FELDMAN (1970), namely,
P
# 7 (subject to the restriction that the homozygotes all have the same fitness) using bifurcation theory, as in HASTINGS (1982). In fact, the calculation given in HASTINGS (1982) is just the calculation needed here. T h e results given by bifurcation theory are not as complete as those possible by an exact determi- nation of the equilibria, but the algebra is much simpler and the stability of the equilibria follows immediately.T h e analysis will depend on an analysis of the corner equilibrium (see BOD-
MER and FELSENSTEIN 1967 for preliminary calculations on the stability of this equilibrium)
p A = ps = 0. (17)
Note that, because of the symmetry assumed here, the same analysis is valid for the same values of r for all four corner equilibria. If
a
<
P,
7, (18)the stability of the corner equilibria depends on r , and they are stable if r
>
7c and unstable if r<
r e , where in the symmetric viability modelIn HASTINGS (1982), I determined the conditions under which a curve of stable polymorphic equilibria (depending on r ) bifurcates from the corner equi- libria. There will be such a curve of stable equilibria for r
<
rc approaching each corner equilibrium as r approaches r,, if and only if the quantityfi,
defined in equation (23) of HASTINGS (1982) is negative. For the corner equi- librium (17),
these equilibria take the form given by equation (22) in HASTINGS (1982). However, the exact form of this first order approximation is not im- portant here. In the symmetric viability case studied here the quantityfi,
simplifies greatly tof i l = a b 2
-
( P
-
r>'I/[(l
-
a)(@-
-
.)I
(20)Under the assumption (18) the denominator in (20) is always positive. Thus,
f i l is negative if and only if
IP
-
rl
>
a. (21)Hence, the results in HASTINGS (1982) show that the following surprising result holds.
If(21) holds and r is less than a, and r
-
CY is suficiently small, the symmetricviability model has four stable polymorphic equilibria.
Here, I am using the fact that none of the symmetric equilibria can be simultaneously stable with these four polymorphic equilibria, which can be shown after some algebra using (1 1 ) and (16). The dependence of these equi- libria on r will be discussed in the numerical example presented in the next section.
A numerical example
Here, I will present a numerical example illustrating the results of the pre- vious section. This example also answers the question of what happens to these four equilibria as r decreases from rc. Let a = 0.1, @ = 0.2, = 0.4. Then, the condition (21) of the previous section is satisfied and there are four stable polymorphic equilibria for r
<
0.1 and r sufficiently close to 0.1. Note also that in this case the symmetric equilibria (13), (14) are stable for 0<
r<
0.076 [the last figure is approximate and follows from (16)].
TABLE 1
Four simultaneously stable polymorphisms
259
~~
0.0955719 0.0294281 0.0737092 0.801 2908
0.0294281 0.0955719 0.8012908 0.0737092
0.0737092 0.8012908 0.0955719 0.0294281
0.8012908 0.0737092 0.0294281 0.0955719
Gametic frequencies at the four stable polymorphic unsymmetric equilibria of the
= 0.4, r = 0.09.
Lewontin-Kojima symmetric viability model for a = 0.1, B = 0.2,
Each row represents an equilibrium.
TABLE 2
Four simultaneously stable polymorphisms
PA PB D
0.1250000 0.1692811 0.0744118
0.1250000 0.8307189 -0.07441 18
0.8750000 0.169281 1 -0.0744118
0.8750000 0.8307189 0.07441 18
Allele frequencies and disequilibria at the four stable poly- morphic unsymmetric equilibria of the Lewontin-Kojima sym-
metric viability model for a = 0.1, 0 = 0.2, y = 0.4, r = 0.09.
Each row represents an equilibrium.
a
W
J
W
-I
J
LL
0
t u
z
w 3
0
w
e
LL
a
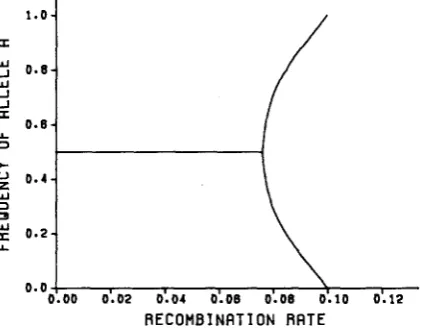
R E C O M B I N A T I O N R R T E
FIGURE 1 .-Allele frequencies at locus A at the four stable polymorphic unsymmetric equilibria
of the Lewontin-Kojima symmetric viability model for a = 0.1, p = 0.2, y = 0.4, as a function of
r . T h e frequency p~ varies similarly. T h e symmetric equilibria from which the unsymmetric equi- libria arise are also shown.
DISCUSSION
behavior possible in multilocus multiallele models. When hitherto unknown complexity in the two-locus two allele model is revealed, the results here sug- gest the possibility of further complexities in more complex models.
Furthermore, I have no reason to be convinced that four is the upper limit to the number of stable equilibria possible in a two-locus two-allele model, since 15 equilibria may be possible (KARLIN 1975). It may in fact be possible to generate an example with even eight stable equilibria, arising through a sequence of pitchfork bifurcations.
Surprisingly, the existence of four stable polymorphisms was not found for some bizarre fitness scheme but was found for the well-studied symmetric viability model. Additionally, the phenomenon can occur for arbitrarily weak selection and for arbitrarily small values of the recombination rate r .
Observations of different populations, thought to be under similar selective pressures, with different gametic frequencies, have been claimed as evidence for the role of drift and founder effect (e.g., SELANDER and FOLTZ 1981). T h e results obtained here render this kind of argument even more suspect.
Finally, the dependence of the stable equilibria studied here on r is most striking. As r is reduced from rc to the root of the function T ( r ) given in (16), the values of the allele frequencies change from 0 or 1 to 0.5. As this range of r values is typically very small, the change is quite dramatic (see Figure 1). In fact, as a approaches
I
/3-
yI,
the range of r values actually goes to zero. Also, the range of r values is certainly bounded by rc, which is a in the case studied here. Thus, for weak selection the change in allele frequencies with r is most dramatic. This is perhaps not surprising. However, the same can hold for strong selection (Figure 1).This research was supported by United States Public Health Service grant R 0 1 GM32 130-01.
LITERATURE CITED
BODMER, W . F. and J. FELSENSTEIN, 1967 Linkage and selection: theoretical analysis of the
deterministic two locus random mating model. Genetics 57: 237-265.
EWENS, W. J., 1968
140-143.
EWENS, W . J., 1979
FELDMAN, M. W. and U. LIBERMANN, 1979
A genetic model having complex linkage behavior. Theor. Appl. Genet. 38:
Mathematical Population Genetics. Springer-Verlag, Berlin.
On the number of stable equilibria and the simul-
taneous stability of fixation and polymorphism in two-locus models. Genetics 92: 1355-1 360.
Nonlinear Oscillations, Dynamical Systems, and Bijkcations of Vector Fields. Springer-Verlag, News York.
Simultaneous stability of D = 0 and D # 0 for multiplicative viabilities at
t w o loci: an analytical study. J. Theor. Biol. 8 9 69-81.
Unexpected behavior in two-locus genetic systems: an analysis of marginal
underdominance at a stable equilibrium. Genetics 102: 129-1 38.
General two-locus selection models: some objectives results and interpretations.
Numerical studies on two-loci selection models with general
GUCKENHEIMER, J. and P. HOLMES, 1983
HASTINGS, A., 1981
HASTINGS, A., 1982
KARLIN, S., 1975
Theor. Pop. Biol. 7: 399-421.
viabilities. Theor. Pop. Biol. 7: 364-398.
Linkage and selection: two locus symmetric viability
A mathematical problem in population genetics. Proc. Camb. Phil. Soc.
T h e evolutionary dynamics of complex polymorphisms.
Gametic disequilibrium between esterase loci in pop-
Communicating editor: W. J. EWENS
KARLIN, S. and M. W. FELDMAN, 1970 model. Theor. Pop. Biol. 1: 39-71.
KINGMAN, J. F. C., 1961
LEWONTIN, R. C. and K . KOJIMA, 1960
SELANDER, R. K. and D. W . FOLTZ, 1981
57: 574-582.
Evolution 14: 458-472.