1443
T
he human coagulation system has evolved over the past 450 million years to function through a complex coordi-nated interaction involving the vessel wall, circulating cells, and a protein cascade of procoagulants that are regulated by a series of anticoagulant mediators.1 In normal hemostaticresponses, these various components combine to generate a protective fibrin clot, limiting blood loss from the circula-tion. In pathological states, deficiencies, dysfunction, or aber-rant regulation of these hemostatic elements result in either bleeding or thrombotic phenotypes. These pathologies are the
consequence of either rare genetic traits or, more often, a com-plex interaction of genetic and acquired influences.
Current Therapeutic Approaches
to Coagulation Disorders
Current treatments for bleeding and thrombotic diseases com-prise either targeted single factor approaches in which specific elements of the hemostatic process are replaced or inhibited or combination strategies that either restore or interfere with multiple components of the hemostatic response. Although an
© 2016 American Heart Association, Inc.
Circulation Research is available at http://circres.ahajournals.org DOI: 10.1161/CIRCRESAHA.115.307015
Abstract:
Molecular genetic details of the human coagulation system were among the first successes of the geneticrevolution in the 1980s. This information led to new molecular diagnostic strategies for inherited disorders of hemostasis and the development of recombinant clotting factors for the treatment of the common inherited bleeding disorders. A longer term goal of this knowledge has been the establishment of gene transfer to provide continuing access to missing or defective hemostatic proteins. Because of the relative infrequency of inherited coagulation factor disorders and the availability of safe and effective alternative means of management, the application of gene therapy for these conditions has been slow to realize clinical application. Nevertheless, the tools for effective and safe gene transfer are now much improved, and we have started to see examples of clinical gene therapy successes. Leading the way has been the use of adeno-associated virus–based strategies for factor IX gene transfer in hemophilia B. Several small phase 1/2 clinical studies using this approach have shown prolonged expression of therapeutically beneficial levels of factor IX. Nevertheless, before the application of gene therapy for coagulation disorders becomes widespread, several obstacles need to be overcome. Immunologic responses to the vector and transgenic protein need to be mitigated, and production strategies for clinical grade vectors require enhancements. There is little doubt that with the development of more efficient and facile strategies for genome editing and the application of other nucleic acid–based approaches to influence the coagulation system, the future of genetic therapies for hemostasis is bright. (Circ Res. 2016;118:1443-1452. DOI: 10.1161/CIRCRESAHA.115.307015.)
Key Words: bleeding ■ coagulation ■ factor IX ■ factor VIII ■ gene therapy ■ gene transfer ■ hemophilia
Gene Therapy for Coagulation Disorders
Laura L. Swystun, David Lillicrap
Circulation Research
Compendium on Thrombosis
Advances in Thrombosis and Hemostasis: An Introduction to the Compendium Global Burden of Thrombosis: Epidemiologic Aspects
Systems Analysis of Thrombus Formation
Animal Models of Thrombosis From Zebrafish to Nonhuman Primates Platelet-Mediated Thrombosis: From Bench to Bedside
Cross Talk Pathways Between Coagulation and Inflammation
Evolving Treatments for Arterial and Venous Thrombosis: Role of the Direct Oral Anticoagulants Evolving Treatments for Acute Ischemic Stroke
Gene Therapy for Coagulation Disorders
Jeffrey I. Weitz and John W. Eikelboom, Editors
Original received October 5, 2015; revision received December 9, 2015; accepted December 16, 2015. From the Department of Pathology and Molecular Medicine, Queen’s University, Kingston, Ontario, Canada.
Correspondence to David Lillicrap, MD, Department of Pathology and Molecular Medicine, Richardson Laboratory, Queen’s University, Kingston, Ontario, Canada K7L 3N6. E-mail [email protected]
initial vasoconstrictive response to vascular damage is crucial for the early phase of hemostasis (eg, reduced blood flow, platelet margination, and unfolding of von Willebrand factor), specific therapies aimed at this early element of hemostasis are not routinely used. In contrast, cellular therapies to enhance the contributions of platelets (adhesion and aggregation) and red cells (contributing to platelet margination and fibrin clot strength)2 to the generation of an optimal hemostatic response
are frequently applied to the treatment of bleeding. Currently, most therapies used for coagulation disorder management are aimed at replacing, enhancing, or inhibiting components of the procoagulant protein cascade and the corresponding anti-coagulant and fibrinolytic response.
Procoagulant Treatment Approaches
Interventions aimed at preventing or treating bleeding can in-volve either single factor strategies or combination replace-ment therapy. The more common inherited bleeding disorders, hemophilia A (factor VIII [FVIII] deficiency) and B (factor IX [FIX] deficiency), von Willebrand disease, and factor XI defi-ciency, can be treated with single factor replacement therapies through the infusion of protein concentrates derived from ei-ther plasma or recombinant DNA technology.3In contrast, most acquired bleeding disorders, such as vitamin K deficiency, liver disease, and consumptive coagu-lopathies, are managed with the infusion of either defined mul-tifactor concentrates (eg, prothrombin complex concentrates, factors II, VII, IX, and X,4 and cryoprecipitate, von Willebrand
factor, factors VIII, XIII, and fibrinogen5) or plasma.
The efficacy of these various treatment regimens can be monitored through clinical assessment and by the perfor-mance of either global tests of hemostasis, such as the pro-thrombin, partial thromboplastin, and thrombin times, or by quantification of single factor levels as in the replacement of FVIII and FIX in the hemophilias.
Gene Therapy Concepts and Historical Context
The concept of using genetic approaches to treat and po-tentially cure disease originated in the early 1980s with the cloning of the first human genes. The first, approved, and suc-cessful gene therapy study in a human was conducted in 2 pa-tients with inherited adenosine deaminase deficiency in 1990.6Since then, the field of gene therapy has had a chequered his-tory, but in 2012, the European Medicines Agency approved for the first time a gene therapy drug Glybera, a gene therapy treatment for inherited lipoprotein lipase deficiency.7 Now in
2015, there is realistic optimism that this therapeutic modality can offer significant opportunities for long-term benefits in a range of disorders.
The coagulation genes were among the first to be char-acterized in the 1980s, with major contributions derived from Earl Davie’s laboratory at the University of Washington (Figure 1A).8–11 These discoveries confirmed that the
coagula-tion proteins were derived from genes which encoded modular domains that were shared between related families of coagu-lation factors (eg, the vitamin K–dependent factors and fac-tors V and VIII). Soon after the cloning of the FVIII and FIX genes, translational application of this knowledge was initi-ated through the introduction of molecular diagnostics,12 the
development of recombinant clotting factors for replacement therapy,13 and the initial preclinical trials of gene therapy.
The hemophilias are an ideal model genetic disease for the application of gene therapy. They result from recessive mu-tations in 2 well-characterized genes with minimal influence from other genetic modifiers. Furthermore, it was well recog-nized from the natural history of moderately severe and mild hemophilia (clotting factor levels between 1% and 40%) that small increments in plasma clotting factor levels would result in significant benefits in reducing the risk of bleeding. Thus began the 30-year quest to convert what seems to be a simple therapeutic goal, to deliver a normal gene copy to compensate for the existing mutant gene.
Spectrum of Gene Therapy Initiatives
for Coagulation Disorders
It is clear, for the reasons detailed earlier, that the hemophil-ias have been leading candidates for the application of gene therapy since the dawn of molecular medicine. However, ad-vances in the development of gene therapy strategies for other pro- or anticoagulant deficiencies has been modest, a fact that relates to a combination of factors, including the infrequent incidence of monogenic inherited bleeding and thrombotic disorders, the existence of safe and effective alternative thera-pies, and in some instances, inadequate biological knowledge to effectively design gene therapy strategies.
Aside from FVIII and FIX gene therapy, the only other procoagulant targets that have been explored to any extent in preclinical studies have been von Willebrand factor14 and
factor VIIa.15 The former target might find a place for the
treatment of the rare type 3 form of von Willebrand disease (incidence 1 per million), but even in this population, with no detectable circulating von Willebrand factor, spontaneous bleeding rates can often be surprisingly low.16 The limited
activated factor VII gene therapy studies have been focused predominantly on the potential for developing a sustainable, long-term strategy for bypassing the presence of FVIII anti-bodies (FVIII inhibitors) in hemophilia A.
Gene therapy for anticoagulant purposes has been simi-larly underdeveloped. Again, this relates to the availability of safe and effective alternative treatments and to the fact that monogenic traits directly responsible for initiating thrombosis are rare. Finally, in contrast to the hemophilias, the levels of transgenic protein expression required to produce a phenotyp-ic benefit are likely to be beyond the capability of current gene transfer technologies.
One thrombotic disorder where gene therapy has pro-duced promising preclinical results is the inherited form of
Nonstandard Abbreviations and Acronyms AAV adeno-associated virus
ADAMTS13 a disintegrin and metalloproteinase with a throm-bospondin type 1 motif, member 13
FVIII factor VIII
FIX factor IX
a disintegrin and metalloproteinase with a thrombospondin type 1 motif, member 13 (ADAMTS13) deficiency (Upshaw– Schulman Disease). Studies in the mouse model of this condi-tion, using an adeno-associated viral (AAV) vector–mediated approach have documented sustained therapeutic levels of transgenic ADAMTS13 expression and a positive effect on the clinical features of the disorder.17 However, with safe and
ef-fective treatment already available for this rare genetic disease through infrequent (every 3–4 weeks) infusion of plasma18
and the development of a recombinant ADAMTS13 protein,19
the likelihood of a gene therapy approach reaching the clinic in the foreseeable future is small.
Gene Therapy Strategies for Coagulation
Disorders
The therapeutic goal for coagulation factor gene therapy is the delivery of a normal gene copy to produce therapeutically ef-fective levels of the protein that is either deficient or dysfunc-tional as a result of a germ line mutation (Figure 1B).
Several strategies for genome editing, using technologies such as zinc finger nucleases and transcription activator-like endonucleases, have the potential for in situ mutation correc-tion. The application of these approaches has proven effective in vitro and in animal studies,20,21 but none of the
methodolo-gies are close to clinical introduction. Zinc finger nuclease ed-iting has also been applied successfully to target therapeutic transgene insertion to a safe and transcriptionally favorable locus.22 The latest technology to be applied to genome editing
uses guide RNA molecules to target the site for revision, and the Cas9 endonuclease to mediate the double strand cleavage event required for sequence correction with a normal donor molecule (clustered regularly interspaced short palindromic repeats/clustered regularly interspaced short palindromic
repeat–associated 9 [CRISPR-Cas9]).23 There is considerable
enthusiasm in the molecular biology community concerning the utility of this technology, and the momentum of CRISPR-Cas9 advances has been dramatic.24 However, despite the
fact that mutant coagulation gene editing has already been reported in vitro,25 several critical limitations to this
technol-ogy remain. The achievable efficiency of mutation correction is still low, and perhaps more problematic is the potential for damaging off-target editing events elsewhere in the genome.26
Similar weaknesses exist for earlier editing approaches involv-ing zinc finvolv-inger nuclease repair and TALEN-based strategies.27
For the present, substantial improvements in both efficiency and safety will need to be developed before any genome edit-ing approach could be considered for clinical application as a treatment for coagulation factor deficiency or dysfunction.
Current gene therapy initiatives for coagulation patholo-gies are essentially gene supplementation stratepatholo-gies in which the mutant gene is untouched and a copy of the normal gene is added to somatic cells of the patient. By its nature, this type of approach is best suited to the treatment of recessive traits in which interference from a dominant mutant product is not a limitation.
Transgene Delivery Approaches
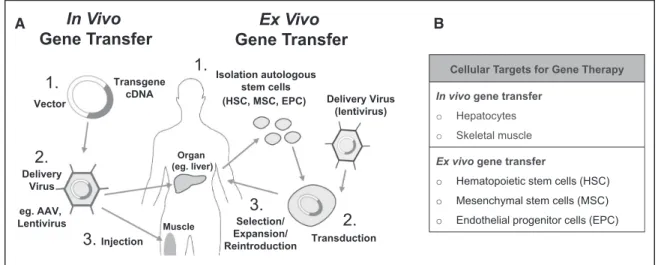
The first critical hurdle to overcome in any gene transfer protocol is the efficient delivery of the therapeutic transgene to somatic cells of the patient (Figure 2A). This goal can be achieved by either in vivo or ex vivo approaches, and there are advantages and weaknesses to both strategies. Ex vivo transgene delivery requires the isolation of a recipient cell population that can be readily transduced and has the potential for prolonged survival after delivery back into the recipient. The advantages of this approach are that efficient rates and Figure 1.Gene therapy as a treatment for coagulation factor disorders. A, Gene therapy is the ultimate goal for treatment of
inherited coagulation disorders. Characterization of the clotting factor genes/proteins in the 1980s allowed for the molecular diagnosis of inherited bleeding and thrombophilic disorders. Subsequently, recombinant protein technologies provided the first clotting factor– specific replacement therapies. Current strategies for improving treatment of inherited coagulation disorders include improving the pharmacokinetics of recombinant protein therapies and increasing the safety and efficacy of gene transfer protocols. B, Potential strategies for gene therapy for coagulation factor deficiencies include viral and nonviral in vivo, methods of delivery of a normal gene copy, ex vivo modification of host cells to induce normal gene expression, and in situ mutation correction. CRISPR-Cas9, clustered regularly interspaced short palindromic repeats/clustered regularly interspaced short palindromic repeat–associated 9; iPS, induced pluripotent stem cells; and TALENS, transcription activator-like effector nucleases.
2013 1990 1986 1984 Recombinant Protein Therapy Gene Therapy Molecular Diagnostics Basic Science Coagulation Factor Gene Therapy Viral Gene Transfer
Retrovirus Adenovirus
Adeno-associated Virus
Mutation Repair
Zinc finger nucleases TALENs
CRISPR-Cas9
Cell-based Gene Therapy
Embryonic stem cells
Adult stem/progenitor cells Patient-derived iPS cells
Non Viral Gene Transfer
Hydrodynamic delivery Targeted nanoparticles
A B
cell-type specificity of transgene delivery can be achieved (Figure 2B) and that the immunologic complications of in vivo vector administration are avoided. However, ex vivo strategies are inherently complex and resource-intensive, and re-establishment of the genetically modified cells may require some form of conditioning to ensure engraftment.28 In
con-trast, in vivo gene transfer is straightforward to perform, but ensuring efficient transgene delivery to specific cell types, without inciting immune reactions to the delivery vehicle, is challenging.
A single human clinical trial for ex vivo delivery of a FVIII transgene has been performed for hemophilia A. This study, involving 12 patients, used autologous fibroblasts that were electroporated ex vivo with a FVIII transgene, selected for op-timal FVIII expression, expanded, and subsequently injected into the omental membrane.29 There were no adverse events
associated with these studies, but also no evidence of sus-tained FVIII transgene expression, although several patients showed evidence of transient (2–3 days) minimal increments of plasma FVIII levels (2%–6%). Other preclinical studies of ex vivo transgene delivery for hemophilia have involved he-matopoietic stem cells, mesenchymal stem cells, and endo-thelial progenitors. Results from the hematopoietic stem cell studies have been especially promising30,31 and may advance
to a phase I/II clinical trial in the near future, but the endo-thelial progenitor investigations have been complicated by the generation of immune responses to the transgene product.32
One particular ex vivo strategy for ectopic hemophilia gene therapy deserves further commentary. The generation of FVIII transgenes that are delivered to and expressed exclusively in megakaryocytes and platelets has shown promise in promot-ing an effective hemostatic response in standard animal mod-els of hemophilia A and in the presence of FVIII inhibitory antibodies.28,33–35 Whether this strategy can be sufficiently
op-timized in terms of transgene delivery and protein production
without the requirement of prohibitive conditioning regimens remains to be established. One potential approach to circum-vent the requirement for bone marrow conditioning is the use of intraosseus vector infusion to mediate in vivo hematopoi-etic cell transduction. Using a lentiviral-based strategy with a platelet-specific regulatory sequence, long-term persistence of intra-platelet FVIII has been demonstrated at sufficient levels to mediate hemostatic protection in the presence of anti-FVIII antibodies.36
Modes of Transgene Delivery
The goal of transgene delivery is to introduce the therapeutic gene construct as efficiently and safely to the recipient, usu-ally to selected types of somatic cells. Importantly, avoid-ance of delivery to germ cells is a requirement for all clinical studies because the safety of germ line modification remains unresolved.
There are essentially 2 modes of transgene delivery: the application of viral vectors or nonviral vector–mediated strat-egies. Some of the latter approaches have been used for in vi-tro molecular biology studies since the 1970s, but in general, physicochemical protocols are relatively inefficient delivery approaches for achieving long-term persistence of transgenes. Electroporation has also been applied in some ex vivo proto-cols29 but again lacks sufficient efficiency for routine clinical
translation. Finally, localized application of hydrodynamic gene transfer protocols has been proposed as a further ap-proach to deliver expression plasmids to specific organs or regions of organs.37,38 However, the inevitable, albeit transient,
vascular and local tissue damage caused by this mode of deliv-ery initiates a strong innate immune response, thus increasing the likelihood of a secondary adaptive response to the neoan-tigenic transgene protein.
In marked contrast to the limitations of the various nonviral modes of transgene delivery, viral vectors use
Figure 2.Methods for transfer of coagulation factor genes. A, Currently used strategies (clinical or preclinical) for transfer of coagulation factor genes include in vivo and ex vivo approaches. In vivo methodologies involve inserting the coagulation factor cDNA into a viral vector genome, with subsequent introduction of the vector into the patient by intravenous or intramuscular injection. Ex vivo gene transfer involves isolation of autologous stem cells. Isolated cells are transduced with the normal coagulation factor gene, selected for protein expression, expanded, and then reintroduced into the patient. B, Cellular targets for in vivo transfer of coagulation factor genes include hepatocytes and skeletal muscle. Cellular targets for ex vivo gene transfer approaches involve isolation and transduction of autologous stem cells including hematopoietic stem cells (HSCs), mesenchymal stem cells (MSCs), and endothelial progenitor cells (EPCs). AAV indicates adeno-associated virus.
Transgene cDNA Vector
In Vivo
Gene Transfer
Organ (eg. liver) MuscleEx Vivo
Gene Transfer
Delivery Virus eg. AAV, Lentivirus1.
2.
3.
Injection1.
Isolation autologous stem cells Transduction2.
Selection/ Expansion/ Reintroduction3.
Delivery Virus (lentivirus) (HSC, MSC, EPC) A Bproperties that have evolved over millions of years to fa-cilitate the efficient entry of viruses into cells.39 The
genera-tion of viral vectors involves the modificagenera-tion of the viral genome to eliminate the possibility of viral replication and to provide sufficient space for insertion of the therapeutic transgene in a viral particle that can still be efficiently pack-aged. Subsequent to the generation of the modified vector genome, the vector particles are produced in cell culture, providing packaging and structural proteins from cotrans-fected helper plasmids.
Gene therapy clinical trials for hemophilia have used 3 types of viral vector (Figure 3A). One patient received an ad-enoviral-mediated treatment for hemophilia A; there has been one trial of retroviral gene transfer of FVIII, and the remain-ing studies have used AAV vectors to deliver FIX transgenes (Table).
In contrast to AAV vectors that are discussed later, ad-enoviral vectors are not appropriate for the delivery of co-agulation transgenes, in large part because of the significant proinflammatory innate immune response that their capsid proteins incite.48 This response can be limited to the
develop-ment of fever, thrombocytopenia, and the elevation of liver transaminases, but can, under certain circumstances, advance to a fatal systemic inflammatory state such as that which killed Jesse Gelsinger during a trial of ornithine transcarbamylase gene therapy in 1999.49–51 Unless this obstacle can be
elimi-nated (which may simultaneously reduce the transduction ef-ficacy of the vector), adenoviral vectors will not find a place in clinical coagulation factor gene therapy.
The single clinical trial using an FVIII retrovirus in he-mophilia A involved the systemic delivery of a classical γ-retroviral vector that would require cellular replication for successful transduction.41 None of the 12 patients entered in
this phase 1 study developed complications, but only low level (≈4%) and transient levels of FVIII were documented, indicat-ing either inadequate transduction and transgene expression.
Although γ-retroviral vectors are no longer being pursued for hemophilia therapy, the development of lentiviral vectors, capable of transducing both dividing and nondividing cells, has continued. Lentiviral vectors can be used as the means for transgene delivery either in ex vivo strategies for modify-ing stem cell populations52,53 or via systemic in vivo delivery.54
Both strategies have been used successfully in animal models of hemophilia,54,55 and some form of lentiviral-based treatment
may well enter the clinic in the future. Nevertheless, in 2015, the clearly favored vector system for clinical coagulation gene therapy was AAV.
AAV-Mediated Gene Therapy for Hemophilia
The development of AAV-based protocols for gene transfer of FIX has been one of the highlights of clinical gene ther-apy advances in the past 5 years. AAVs are small infectious agents (≈20 nm) that belong to the family of parvoviruses. In humans, infection with AAV is asymptomatic. AAV has a small single-stranded DNA genome of ≈4.7 kb, with 2 open reading frames that encode capsid and other structural pro-teins and propro-teins facilitating integration of the wild-type virus into a specific region on chromosome 19. In the con-struction of AAV vectors, both these coding regions are de-leted, allowing the insertion of transgenes of ≤≈5 kb without adversely influencing the packaging efficiency of vector par-ticles. This packaging limitation has not affected the genera-tion of FIX transgenes, in which the coding sequence spans ≈1.3 kb. In contrast, the FVIII cDNA occupies ≈8.5 kb, and even after deletion of the nonessential B domain–encoding sequence, the cDNA is still ≈5 kb. Thus, promoter/enhancer elements for AAV FVIII constructs have to be compact, and even then, a reduction in vector packaging efficiency is a significant concern (Figure 3B).To date, human clinical trials of skeletal muscle43 and
liv-er-directed44,45 AAV FIX gene therapy have been conducted.
In each instance, strong tissue-specific promoter/enhancer
Figure 3.Viral vectors for in vivo gene transfer. A, Gene therapy clinical trials for hemophilia have used adenovirus (1 patient), γ-retrovirus (12 patients), and adeno-associated virus (AAV; 31 patients).
B, Adenovirus-associated inflammation and inadequate transduction and transgene expression by γ-retrovirus have discontinued their use in coagulation factor gene therapy. One challenge of AAV-mediated gene therapy is the limited packaging size of the recombinant vector genome. Strategies for overcoming this limitation include cDNA contraction and the use of compact promoter/enhancer elements. ITR indicates inverted terminal repeat.
Viral Vector Characteristics Advantages Limitation Adenovirus 36 Kb dsDNANon-enveloped
Non-integrating
Large genome Easy to produce high titer
Infects many cell types High immunogenicity
Retrovirus (lentivirus) 8 Kb ssRNA Enveloped Integrating Larger genome High infection efficiency
Stable gene transfer Insertional mutagenesis
Adeno-associated virus (AAV) 4.7 Kb ssDNA Non-enveloped Non-integrating Low immunogenicity Infects many cell types
Long-term gene transfer Small genome
Rep Cap
ITR ITR
4.7 kb ss DNA
WT AAV Genome AAV Vector
Promoter Transgene cDNA
ITR Enhancer ITR
~5 kb ss DNA
5’ Regulatory Elements 3’ Regulatory Elements • Constitutive • Regulated Cell-specific Developmental-specific Drug-inducible • Enhancer • Poly A tail Codon Optimization/ cDNA contraction
A
B
elements have been used, and the vectors have been delivered by either intramuscular injection or portal or peripheral vein injection for the skeletal muscle and liver studies, respectively. Tissue tropism of the AAV vectors is influenced by the differ-ential efficacy for transduction mediated by serotype-specific capsids.56 Thus, AAV8 possesses strong hepatotropism and
can be delivered efficiently to the liver via peripheral vein infusion without the risk of significant transduction of other tissue types.
Fate of AAV Vectors
Transduction by AAV vectors involves an interaction with a range of serotype-specific cell surface receptors.56–58 The
vector particles are taken into endocytic vesicles, where, in the acidic milieu of the endosome, the capsid is removed and the nucleic acid released for transport to the nucleus.59 The
capsid proteins are then degraded by the proteasome, and the resultant peptides presented on the transduced cell’s sur-face, in association with major histocompatibility complex class I molecules (Figure 4B). The vector genome enters the nucleus and persists long term, largely as circular extrachro-mosomal concatemers. However, AAV-based transgenes will integrate into host cell sequences at sites of natural chromo-some fragility, a process that is accompanied by small ad-jacent insertion/deletion events.60 Although integration into
the host genome is a rare phenomenon for AAV, the large vector particle doses that are administered in clinical gene therapy (≈1012 particles/kg) results in millions of transduced
cells with AAV insertions. There has been a single report of increased tumor incidence (hepatocellular carcinoma) in a mouse AAV gene transfer model,61 but these observations
were made in old normal and mucopolysaccharidosis VII mice after injection of the vectors in neonates. This outcome has not been replicated in any other long-term small or large animal model of AAV therapy.
Results of AAV-Mediated Gene
Therapy for Hemophilia
To date, ≈31 hemophilia B patients have been treated in 5 AAV gene therapy trials (Table). A clinical study using AAV to deliver FVIII has just enrolled its first patient. In the he-mophilia B trials, initial studies involved intramuscular injec-tion of the vector, showing that long-term expression of the FIX transgene was achievable.43,62,63 Latterly, hemophilia gene
therapy trials have targeted the liver for transgene delivery using either portal vein infusion or, more recently, peripheral vein injection of hepatotropic AAV8 vectors.
The FIX transgene constructs that have been generated have used small and potent tissue-specific enhancer/pro-moter elements, and one study has used a codon optimized FIX cDNA.45 In the most recent FIX clinical trial, the FIX
transgene also contains a gain-of-function missense variation (Arg338Leu–FIX Padua64,65) that increases the specific
coagu-lant activity of the transgenic protein by ≈7-fold.
For native single-stranded AAV transgenes to mediate mRNA expression, a second complementary DNA strand has first to be synthesized. In efforts to accelerate this process, some investigators have constructed self-complementary AAV transgenes that mediate earlier expression after delivery.57
Vector doses in the clinical trials have ranged between 2×1011
and 5×1012 vector genomes/kg.
After transgene delivery, FIX plasma levels start to in-crease after 2 to 3 weeks and have, until the most recent study using the FIX Padua transgene, resulted in FIX levels of 2% to 10% in the short term and persistent levels of 1% to 6%.43,45,46 Preliminary results from the FIX Padua study,
in patients receiving one of the higher vector doses, have shown transient levels of FIX >50% that have subsequently fallen to levels similar to the other studies that have been reported.66 In the context of hemophilia, these relatively
low levels of FIX have had significant benefits in terms of Table. Summary of Completed and Ongoing Phase I/II Clinical Trials for Hemophilia Gene Therapy
Vehicle
Transfer Method Gene
Patients
Treated Best Outcome Complications Reference Gamma-retrovirus transduction of
autologous fibroblasts
Ex vivo F9 2 Transient expression (2%) No adverse effects reported
Xinfang et al40
Electroporation of autologous fibroblasts (BDD-FVIII)
Ex vivo F8 12 Transient expression (4%) No adverse effects reported
Roth et al29
Gamma-retrovirus (BDD-FVIII)–IV In vitro F8 12 Transient expression (4%) No adverse effects reported
Powell et al41
“Gutless” adenovirus In vitro F8 1 Transient expression (1%–3%) Transaminitis, thrombocytopenia
No published reports42
ssAAV2/2–Intramuscular In vitro F9 8 Transient expression (<2%) No adverse effects reported
Manno et al43
ssAAV2/2–hepatic artery infusion In vitro F9 7 Transient expression (10%) over 8 wk
Transient transaminitis Manno et al44
scAAV2/8: (FIXco)–IV In vitro F9 10 Persistent expression (1%–6%) ≤3 y
Transient transaminitis Nathwani et al45,46
scAAV2/8: (FIX Padua)–IV In vitro F9 6 Variable expression (0.5%–25%) ≤6 mo
No adverse effects reported
Monahan et al47
BDD indicates B-domain deleted; FIXco, codon optimized FIX; sc, self-complementary; and ss, single strand.
reducing bleeding frequency, with patients being able to stop their routine FIX protein prophylactic infusions. In the study with the longest follow up of 10 hemophilia B patients, therapeutically effective levels of FIX between 1% and 5% have been maintained for a median of 3.2 years after single peripheral vein vector infusions.46 Follow-up of patients in
all of these trials is ongoing.
Challenges to Gene Therapy for Hemophilia
The first obvious fact is that progress with FIX gene therapy has been significantly advanced compared with FVIII. This is because of a combination of limitations presented by FVIII. First, the FVIII cDNA, even with the B domain–encoding region deleted, is ≈4.5 kb, and thus, generation of FVIII transgene constructs that can be efficiently packaged in AAV particles is difficult. Second, achieving high-level expression of FVIII has been notoriously problematic, for reasons that have been only partially explained. Finally, the immunogenic potential of FVIII is significantly greater than that of FIX, and preclinical studies of FVIII gene transfer have frequently been complicated by the development of an anti-FVIII immune re-sponse.32,67,68 In contrast, liver-directed AAV gene therapy inhemophilic dogs with anti-FVIII antibodies has also shown potential for inducing immunologic tolerance to FVIII.69
Despite these various challenges, a new AAV-mediated FVIII gene therapy study has just been initiated, and the outcome of this trial will be eagerly awaited.
In contrast to the challenges presented by FVIII gene transfer, the studies of FIX delivery have not been complicat-ed by immune responses to the transgenic FIX protein, and in the latest clinical trials, plasma levels of transgenic FIX have been adequate to produce a significant long-term clinical ben-efit. Nevertheless, the predictability and consistency of these FIX levels are still not well understood.
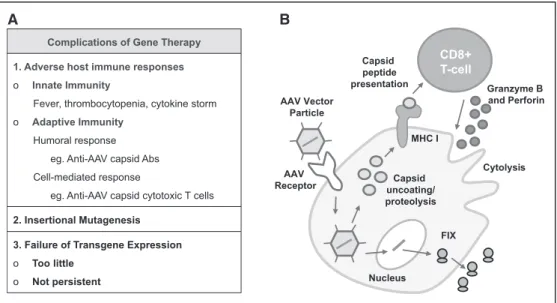
The most significant problem that has arisen in the recent AAV clinical trials has been the development, in ≈25% of
patients, of an anti-AAV capsid immune response that has resulted in transduced hepatocyte cytotoxicity.56,70,71 This
complication is the result of CD8+ cytotoxic T cell responses
to AAV capsid peptides being presented on the surface of transduced hepatocytes (Figure 4A). The timing of the sub-sequent transaminitis that develops has varied from as early as 3 weeks to as late as 10 weeks post vector delivery, prob-ably reflecting either a secondary recall response or a later primary immune response to the AAV capsid. With the aim of minimizing this immunologic problem, patients with evi-dence of preexisting anti-AAV immunity, as demonstrated by the presence of anti-AAV antibodies, are currently ex-cluded from participation in these clinical trials. This affects different numbers of patients for the different AAV serotypes but ranges from ≈30% of the population with antibodies to AAV8 to ≈60% of the population with preexisting anti-AAV2 immunity.72 Unfortunately, even with the generation
of novel AAV capsids, the issue of cross reactivity of the immune response may still be an obstacle to avoiding the subsequent cellular immune attack.73
With the development of a cytotoxic T cell response to AAV capsid peptides, transduced liver cells are killed and the transgene is lost, thus prevention or rapid intervention of this immune attack is essential to achieve long-term transgene ex-pression. To date, mitigation and eventual extinction of the immune response has been possible with transient cortico-steroid administration started after early recognition of liver transaminase increases.45 Although the use of prophylactic
im-munosuppressive therapy for this complication has been dis-cussed repeatedly, this strategy has not yet been used. Finally, in addition to the cell-mediated immune response to the AAV capsid, a potent humoral response is consistently documented after AAV vector administration that prevents effective repeat AAV delivery. This obstacle to vector readministration might be circumvented by strategies such as infusing capsid decoys to divert the antibody blockade.74
Figure 4.Complications of gene therapy. A, Complications of gene therapy include innate and adaptive immune responses, insertional mutagenesis, and failure of transgene expression. B, Approximately 25% of patients treated with adeno-associated virus (AAV)–mediated gene therapy develop an anti-AAV capsid cell–mediated immune response. Presentation of AAV capsid protein to cognate CD8+ T-cells
results in cytolysis of transgene-expressing cells. MHC indicates major histocompatibility complex.
Complications of Gene Therapy 1. Adverse host immune responses
o Innate Immunity
Fever, thrombocytopenia, cytokine storm o Adaptive Immunity
Humoral response eg. Anti-AAV capsid Abs Cell-mediated response
eg. Anti-AAV capsid cytotoxic T cells
2. Insertional Mutagenesis 3. Failure of Transgene Expression
o Too little o Not persistent AAV Vector Particle Nucleus Capsid uncoating/ proteolysis Capsid peptide presentation MHC I CD8+ T-cell Granzyme B and Perforin Cytolysis AAV Receptor FIX A B
Future Considerations for the Application of
Gene Therapy for Coagulation Disorders
After a 30-year period of in vitro experimentation and pre-clinical assessment, the field of gene therapy is beginning to show robust evidence of clinical benefit in a range of genetic disorders. The recent success of gene transfer for hemophilia B highlights the potential of this therapeutic modality for management of coagulation pathologies. As further evidence of the promise of gene therapy initiatives, the partnership in hemophilia gene therapy trials by the biopharmaceuti-cal industry has increased dramatibiopharmaceuti-cally in the past 2 years. Nevertheless, before gene therapy can be extended to wide-spread clinical utility, several critical hurdles need to be overcome. First, is the development of gene therapy vectors that can be produced in large scale with high and reproduc-ible quality.75 Evidence that the AAV vectors used in a recentFIX clinical trial contained only ≈10% of transgene contain-ing particles illustrates the need for improved and more effi-cient vector production protocols. With current clinical trials being limited to study populations of 5 to 10 patients, there is a long way to go before the widespread application of gene transfer can be envisaged.76
Next is the issue of immune obstacles to successful gene transfer. With levels of preexisting immunity to current AAV-based vectors ranging from 30% to 60%, many otherwise eligible patients are excluded from this form of treatment. Whether AAV capsid modifications or the use of novel AAV serotypes will circumvent this obstacle remains to be seen. Similarly, the use of other vector types, such as lentiviral con-structs, would substantially reduce this problem. Aside from the problems posed by immune responses to the vector, im-mune reactions to the novel transgenic protein may also com-plicate some applications of gene transfer, particularly when the transgenic protein presents peptide sequences that are novel to the recipient.
Although immediate adverse effects of gene transfer us-ing AAV and lentiviral vectors have been negligible, the long-term outcome of gene therapy will require formal monitoring, particularly for genotoxicity outcomes. Studies performed in small and, more critically, large animal models with greater longevity have not shown any evidence of an enhanced inci-dence of chronic pathologies (most importantly, no eviinci-dence of increased cancer development).77 Nevertheless, these
ob-servations will need to be strengthened by formal long-term, multi-year surveillance in human gene therapy recipients. Finally, the efficacy of gene therapy in contrast to currently available or next-generation infused factor replacement thera-pies will need to be evaluated in randomized clinical trials.78
The promise of genetic therapies for improved manage-ment of coagulation disorders is now beginning to be realized. Although gene replacement strategies are the most promi-nent of these approaches, the application of inhibitory oligo-nucleotides79 and small inhibitory RNA molecules80 to alter
the hemostatic balance has demonstrated how other nucleic acid–based strategies have shown considerable potential in recent clinical trials. With enhanced access to genome editing technologies, this momentum toward translational benefits is likely to continue.
Sources of Funding
D. Lillicrap is the recipient of a Canada Research Chair in Molecular Hemostasis. L.L. Swystun is the recipient of a CIHR fellowship.
Disclosures
None.References
1. Davidson CJ, Tuddenham EG, McVey JH. 450 million years of hemosta-sis. J Thromb Haemost. 2003;1:1487–1494.
2. Cines DB, Lebedeva T, Nagaswami C, Hayes V, Massefski W, Litvinov RI, Rauova L, Lowery TJ, Weisel JW. Clot contraction: compres-sion of erythrocytes into tightly packed polyhedra and redistribution of platelets and fibrin. Blood. 2014;123:1596–1603. doi: 10.1182/ blood-2013-08-523860.
3. Key NS, Negrier C. Coagulation factor concentrates: past, present, and future. Lancet. 2007;370:439–448. doi: 10.1016/S0140-6736(07)61199-4. 4. Rodgers GM. Prothrombin complex concentrates in emergency bleeding
disorders. Am J Hematol. 2012;87:898–902. doi: 10.1002/ajh.23254. 5. Nascimento B, Goodnough LT, Levy JH. Cryoprecipitate therapy. Br J
Anaesth. 2014;113:922–934. doi: 10.1093/bja/aeu158.
6. Blaese RM, Culver KW, Chang L, Anderson WF, Mullen C, Nienhuis A, Carter C, Dunbar C, Leitman S, Berger M. Treatment of severe combined immunodeficiency disease (SCID) due to adenosine deaminase deficiency with CD34+ selected autologous peripheral blood cells transduced with a human ADA gene. Amendment to clinical research project, Project 90-C-195, January 10, 1992. Hum Gene Ther. 1993;4:521–527. doi: 10.1089/ hum.1993.4.4-521.
7. Ylä-Herttuala S. Endgame: glybera finally recommended for ap-proval as the first gene therapy drug in the European union. Mol Ther. 2012;20:1831–1832. doi: 10.1038/mt.2012.194.
8. Degen SJ, MacGillivray RT, Davie EW. Characterization of the comple-mentary deoxyribonucleic acid and gene coding for human prothrombin.
Biochemistry. 1983;22:2087–2097.
9. Chung DW, Rixon MW, Que BG, Davie EW. Cloning of fibrinogen genes and their cDNA. Ann N Y Acad Sci. 1983;408:449–456.
10. Leytus SP, Chung DW, Kisiel W, Kurachi K, Davie EW. Characterization of a cDNA coding for human factor X. Proc Natl Acad Sci U S A. 1984;81:3699–3702.
11. Kurachi K, Davie EW. Isolation and characterization of a cDNA coding for human factor IX. Proc Natl Acad Sci U S A. 1982;79:6461–6464. 12. Kogan SC, Doherty M, Gitschier J. An improved method for prenatal
diagnosis of genetic diseases by analysis of amplified DNA sequences. Application to hemophilia A. N Engl J Med. 1987;317:985–990. doi: 10.1056/NEJM198710153171603.
13. Wood WI, Capon DJ, Simonsen CC, Eaton DL, Gitschier J, Keyt B, Seeburg PH, Smith DH, Hollingshead P, Wion KL, Delwart E, Tuddenham EG, Vehar GA, Lawn RM. Expression of active human factor VIII from recombinant DNA clones. Nature. 1984;312:330–337.
14. De Meyer SF, Vanhoorelbeke K, Chuah MK, Pareyn I, Gillijns V, Hebbel RP, Collen D, Deckmyn H, VandenDriessche T. Phenotypic correction of von Willebrand disease type 3 blood-derived endothelial cells with len-tiviral vectors expressing von Willebrand factor. Blood. 2006;107:4728– 4736. doi: 10.1182/blood-2005-09-3605.
15. Margaritis P. Gene-based continuous expression of FVIIa for the treatment of hemophilia. Front Biosci (Schol Ed). 2012;4:287–299.
16. Eikenboom JC. Congenital von Willebrand disease type 3: clinical mani-festations, pathophysiology and molecular biology. Best Pract Res Clin Haematol. 2001;14:365–379. doi: 10.1053/beha.2001.0139.
17. Jin SY, Xiao J, Bao J, Zhou S, Wright JF, Zheng XL. AAV-mediated ex-pression of an ADAMTS13 variant prevents shigatoxin-induced throm-botic thrombocytopenic purpura. Blood. 2013;121:3825–3829, S1. doi: 10.1182/blood-2013-02-486779.
18. Scully M, Hunt BJ, Benjamin S, Liesner R, Rose P, Peyvandi F, Cheung B, Machin SJ; British Committee for Standards in Haematology. Guidelines on the diagnosis and management of thrombotic thrombocy-topenic purpura and other thrombotic microangiopathies. Br J Haematol. 2012;158:323–335. doi: 10.1111/j.1365-2141.2012.09167.x.
19. Plaimauer B, Kremer Hovinga JA, Juno C, Wolfsegger MJ, Skalicky S, Schmidt M, Grillberger L, Hasslacher M, Knöbl P, Ehrlich H, Scheiflinger F. Recombinant ADAMTS13 normalizes von Willebrand factor-cleaving activity in plasma of acquired TTP patients by
overriding inhibitory antibodies. J Thromb Haemost. 2011;9:936–944. doi: 10.1111/j.1538-7836.2011.04224.x.
20. Park CY, Kim J, Kweon J, Son JS, Lee JS, Yoo JE, Cho SR, Kim JH, Kim JS, Kim DW. Targeted inversion and reversion of the blood coagulation factor 8 gene in human iPS cells using TALENs. Proc Natl Acad Sci U S A. 2014;111:9253–9258. doi: 10.1073/pnas.1323941111.
21. Li H, Haurigot V, Doyon Y, et al. In vivo genome editing restores haemo-stasis in a mouse model of haemophilia. Nature. 2011;475:217–221. doi: 10.1038/nature10177.
22. Sharma R, Anguela XM, Doyon Y, et al. In vivo genome editing of the albumin locus as a platform for protein replacement therapy. Blood. 2015;126:1777–1784. doi: 10.1182/blood-2014-12-615492.
23. Sander JD, Joung JK. CRISPR-Cas systems for editing, regulating and targeting genomes. Nat Biotechnol. 2014;32:347–355. doi: 10.1038/ nbt.2842.
24. Zhang F, Wen Y, Guo X. CRISPR/Cas9 for genome editing: progress, implications and challenges. Hum Mol Genet. 2014;23:R40–R46. doi: 10.1093/hmg/ddu125.
25. Park CY, Kim DH, Son JS, Sung JJ, Lee J, Bae S, Kim JH, Kim DW, Kim JS. Functional Correction of Large Factor VIII Gene Chromosomal Inversions in Hemophilia A Patient-Derived iPSCs Using CRISPR-Cas9.
Cell Stem Cell. 2015;17:213–220. doi: 10.1016/j.stem.2015.07.001. 26. Gori JL, Hsu PD, Maeder ML, Shen S, Welstead GG, Bumcrot D. Delivery
and Specificity of CRISPR-Cas9 Genome Editing Technologies for Human Gene Therapy. Hum Gene Ther. 2015;26:443–451. doi: 10.1089/ hum.2015.074.
27. LaFountaine JS, Fathe K, Smyth HD. Delivery and therapeutic applica-tions of gene editing technologies ZFNs, TALENs, and CRISPR/Cas9. Int J Pharm. 2015;494:180–194. doi: 10.1016/j.ijpharm.2015.08.029. 28. Shi Q, Fahs SA, Wilcox DA, Kuether EL, Morateck PA, Mareno N, Weiler
H, Montgomery RR. Syngeneic transplantation of hematopoietic stem cells that are genetically modified to express factor VIII in platelets re-stores hemostasis to hemophilia A mice with preexisting FVIII immunity.
Blood. 2008;112:2713–2721. doi: 10.1182/blood-2008-02-138214. 29. Roth DA, Tawa NE Jr, O’Brien JM, Treco DA, Selden RF; Factor VIII
Transkaryotic Therapy Study Group. Nonviral transfer of the gene encod-ing coagulation factor VIII in patients with severe hemophilia A. N Engl J Med. 2001;344:1735–1742. doi: 10.1056/NEJM200106073442301. 30. Ide LM, Gangadharan B, Chiang KY, Doering CB, Spencer HT.
Hematopoietic stem-cell gene therapy of hemophilia A incorporating a porcine factor VIII transgene and nonmyeloablative conditioning regi-mens. Blood. 2007;110:2855–2863. doi: 10.1182/blood-2007-04-082602. 31. Zakas PM, Spencer HT, Doering CB. Engineered hematopoietic stem cells
as therapeutics for hemophilia A. J Genet Syndr Gene Ther. 2011;1:2410. doi: 10.4172/2157-7412.S1-003.
32. Ozelo MC, Vidal B, Brown C, Notley C, Hegadorn C, Webster S, Harpell L, Ahlin J, Winterborn A, Handforth J, Arruda VR, Hough C, Lillicrap D. Omental implantation of BOECs in hemophilia dogs results in circulating FVIII antigen and a complex immune response. Blood. 2014;123:4045– 4053. doi: 10.1182/blood-2013-12-545780.
33. Yarovoi HV, Kufrin D, Eslin DE, Thornton MA, Haberichter SL, Shi Q, Zhu H, Camire R, Fakharzadeh SS, Kowalska MA, Wilcox DA, Sachais BS, Montgomery RR, Poncz M. Factor VIII ectopically expressed in plate-lets: efficacy in hemophilia A treatment. Blood. 2003;102:4006–4013. doi: 10.1182/blood-2003-05-1519.
34. Shi Q, Wilcox DA, Fahs SA, Fang J, Johnson BD, DU LM, Desai D, Montgomery RR. Lentivirus-mediated platelet-derived factor VIII gene therapy in murine haemophilia A. J Thromb Haemost. 2007;5:352–361. doi: 10.1111/j.1538-7836.2007.02346.x.
35. Shi Q, Montgomery RR. Platelets as delivery systems for disease treat-ments. Adv Drug Deliv Rev. 2010;62:1196–1203. doi: 10.1016/j. addr.2010.06.007.
36. Wang X, Shin SC, Chiang AF, Khan I, Pan D, Rawlings DJ, Miao CH. Intraosseous delivery of lentiviral vectors targeting factor VIII expression in platelets corrects murine hemophilia A. Mol Ther. 2015;23:617–626. doi: 10.1038/mt.2015.20.
37. Sawyer GJ, Rela M, Davenport M, Whitehorne M, Zhang X, Fabre JW. Hydrodynamic gene delivery to the liver: theoretical and practical issues for clinical application. Curr Gene Ther. 2009;9:128–135.
38. Herrero MJ, Sabater L, Guenechea G, Sendra L, Montilla AI, Abargues R, Navarro V, Aliño SF. DNA delivery to ‘ex vivo’ human liver segments.
Gene Ther. 2012;19:504–512. doi: 10.1038/gt.2011.144.
39. Kay MA, Glorioso JC, Naldini L. Viral vectors for gene therapy: the art of turning infectious agents into vehicles of therapeutics. Nat Med. 2001;7:33–40. doi: 10.1038/83324.
40. Xinfang Q, Daru L, Jiemin Z, Jianmin T, Jianmin Y, Peilin M, Hsueh J. Implantation of autologous skin fibroblas genetically modified to secrete clotting factor IX partially corrects the hemor-rhagic tendencies in two hemophilia B patients. Chin Med J (Engl). 1996;109:832–839.
41. Powell JS, Ragni MV, White GC II, Lusher JM, Hillman-Wiseman C, Moon TE, Cole V, Ramanathan-Girish S, Roehl H, Sajjadi N, Jolly DJ, Hurst D. Phase 1 trial of FVIII gene transfer for severe hemophilia A us-ing a retroviral construct administered by peripheral intravenous infusion.
Blood. 2003;102:2038–2045. doi: 10.1182/blood-2003-01-0167. 42. Mannucci PM. Hemophilia and Related Bleeding Disorders: A Story of
Dismay and Success. Hematology. 2002;1–9.
43. Manno CS, Chew AJ, Hutchison S, et al. AAV-mediated factor IX gene transfer to skeletal muscle in patients with severe hemophilia B. Blood. 2003;101:2963–2972. doi: 10.1182/blood-2002-10-3296.
44. Manno CS, Pierce GF, Arruda VR, et al. Successful transduction of liver in hemophilia by AAV-Factor IX and limitations imposed by the host im-mune response. Nat Med. 2006;12:342–347. doi: 10.1038/nm1358. 45. Nathwani AC, Tuddenham EG, Rangarajan S, et al. Adenovirus-associated
virus vector-mediated gene transfer in hemophilia B. N Engl J Med. 2011;365:2357–2365. doi: 10.1056/NEJMoa1108046.
46. Nathwani AC, Reiss UM, Tuddenham EG, et al. Long-term safety and efficacy of factor IX gene therapy in hemophilia B. N Engl J Med. 2014;371:1994–2004. doi: 10.1056/NEJMoa1407309.
47. Monahan P, Walsh C, Powell J, et al. Update on a phase 1/2 open-label trail of BAX335, an adeno-associated virus 8 (AAV8) vector-based gene therapy program for hemophilia B. J Thromb Haem. 2015;13:87. 48. Zhang Y, Chirmule N, Gao GP, Qian R, Croyle M, Joshi B, Tazelaar
J, Wilson JM. Acute cytokine response to systemic adenoviral vec-tors in mice is mediated by dendritic cells and macrophages. Mol Ther. 2001;3:697–707. doi: 10.1006/mthe.2001.0329.
49. Raper SE, Chirmule N, Lee FS, Wivel NA, Bagg A, Gao GP, Wilson JM, Batshaw ML. Fatal systemic inflammatory response syndrome in a orni-thine transcarbamylase deficient patient following adenoviral gene trans-fer. Mol Genet Metab. 2003;80:148–158.
50. Somia N, Verma IM. Gene therapy: trials and tribulations. Nat Rev Genet. 2000;1:91–99. doi: 10.1038/35038533.
51. Marshall E. Gene therapy death prompts review of adenovirus vector.
Science. 1999;286:2244–2245.
52. Aiuti A, Biasco L, Scaramuzza S, et al. Lentiviral hematopoietic stem cell gene therapy in patients with Wiskott-Aldrich syndrome. Science. 2013;341:1233151. doi: 10.1126/science.1233151.
53. Biffi A, Montini E, Lorioli L, et al. Lentiviral hematopoietic stem cell gene therapy benefits metachromatic leukodystrophy. Science. 2013;341:1233158. doi: 10.1126/science.1233158.
54. Cantore A, Ranzani M, Bartholomae CC, et al. Liver-directed lenti-viral gene therapy in a dog model of hemophilia B. Sci Transl Med. 2015;7:277ra28. doi: 10.1126/scitranslmed.aaa1405.
55. Chen Y, Schroeder JA, Kuether EL, Zhang G, Shi Q. Platelet gene therapy by lentiviral gene delivery to hematopoietic stem cells restores hemosta-sis and induces humoral immune tolerance in FIX(null) mice. Mol Ther. 2014;22:169–177. doi: 10.1038/mt.2013.197.
56. Arruda VR, Xiao W. It’s all about the clothing: capsid domination in the adeno-associated viral vector world. J Thromb Haemost. 2007;5:12–15. doi: 10.1111/j.1538-7836.2006.02262.x.
57. Wu Z, Asokan A, Samulski RJ. Adeno-associated virus serotypes: vec-tor toolkit for human gene therapy. Mol Ther. 2006;14:316–327. doi: 10.1016/j.ymthe.2006.05.009.
58. Van Vliet KM, Blouin V, Brument N, Agbandje-McKenna M, Snyder RO. The role of the adeno-associated virus capsid in gene transfer. Methods Mol Biol. 2008;437:51–91. doi: 10.1007/978-1-59745-210-6_2. 59. Ding W, Zhang L, Yan Z, Engelhardt JF. Intracellular trafficking of
ade-no-associated viral vectors. Gene Ther. 2005;12:873–880. doi: 10.1038/ sj.gt.3302527.
60. Nakai H, Wu X, Fuess S, Storm TA, Munroe D, Montini E, Burgess SM, Grompe M, Kay MA. Large-scale molecular characterization of adeno-associated virus vector integration in mouse liver. J Virol. 2005;79:3606– 3614. doi: 10.1128/JVI.79.6.3606-3614.2005.
61. Donsante A, Vogler C, Muzyczka N, Crawford JM, Barker J, Flotte T, Campbell-Thompson M, Daly T, Sands MS. Observed incidence of tu-morigenesis in long-term rodent studies of rAAV vectors. Gene Ther. 2001;8:1343–1346. doi: 10.1038/sj.gt.3301541.
62. Jiang H, Pierce GF, Ozelo MC, de Paula EV, Vargas JA, Smith P, Sommer J, Luk A, Manno CS, High KA, Arruda VR. Evidence of multiyear factor IX expression by AAV-mediated gene transfer to skeletal muscle in an
individual with severe hemophilia B. Mol Ther. 2006;14:452–455. doi: 10.1016/j.ymthe.2006.05.004.
63. Buchlis G, Podsakoff GM, Radu A, Hawk SM, Flake AW, Mingozzi F, High KA. Factor IX expression in skeletal muscle of a severe he-mophilia B patient 10 years after AAV-mediated gene transfer. Blood. 2012;119:3038–3041. doi: 10.1182/blood-2011-09-382317.
64. Simioni P, Tormene D, Tognin G, Gavasso S, Bulato C, Iacobelli NP, Finn JD, Spiezia L, Radu C, Arruda VR. X-linked thrombophilia with a mutant factor IX (factor IX Padua). N Engl J Med. 2009;361:1671–1675. doi: 10.1056/NEJMoa0904377.
65. Monahan PE, Sun J, Gui T, et al. Employing a gain-of-function factor IX variant R338L to advance the efficacy and safety of hemophilia B human gene therapy: preclinical evaluation supporting an ongoing adeno-asso-ciated virus clinical trial. Hum Gene Ther. 2015;26:69–81. doi: 10.1089/ hum.2014.106.
66. Monahan PE. Gene therapy in an era of emerging treatment options for hemophilia B. J Thromb Haemost. 2015;13(suppl 1):S151–S160. doi: 10.1111/jth.12957.
67. Rawle FE, Shi CX, Brown B, McKinven A, Tinlin S, Graham FL, Hough C, Lillicrap D. Heterogeneity of the immune response to adenovirus-medi-ated factor VIII gene therapy in different inbred hemophilic mouse strains.
J Gene Med. 2004;6:1358–1368. doi: 10.1002/jgm.624.
68. McIntosh J, Lenting PJ, Rosales C, et al. Therapeutic levels of FVIII following a single peripheral vein administration of rAAV vector encod-ing a novel human factor VIII variant. Blood. 2013;121:3335–3344. doi: 10.1182/blood-2012-10-462200.
69. Finn JD, Ozelo MC, Sabatino DE, Franck HW, Merricks EP, Crudele JM, Zhou S, Kazazian HH, Lillicrap D, Nichols TC, Arruda VR. Eradication of neutralizing antibodies to factor VIII in canine hemophilia A after liver gene therapy. Blood. 2010;116:5842–5848. doi: 10.1182/ blood-2010-06-288001.
70. Mingozzi F, Maus MV, Hui DJ, Sabatino DE, Murphy SL, Rasko JE, Ragni MV, Manno CS, Sommer J, Jiang H, Pierce GF, Ertl HC, High KA.
CD8(+) T-cell responses to adeno-associated virus capsid in humans. Nat Med. 2007;13:419–422. doi: 10.1038/nm1549.
71. Mingozzi F, High KA. Immune responses to AAV in clinical trials. Curr Gene Ther. 2011;11:321–330.
72. Calcedo R, Morizono H, Wang L, McCarter R, He J, Jones D, Batshaw ML, Wilson JM. Adeno-associated virus antibody profiles in newborns, children, and adolescents. Clin Vaccine Immunol. 2011;18:1586–1588. doi: 10.1128/CVI.05107-11.
73. Gurda BL, DiMattia MA, Miller EB, et al. Capsid antibodies to differ-ent adeno-associated virus serotypes bind common regions. J Virol. 2013;87:9111–9124. doi: 10.1128/JVI.00622-13.
74. Mingozzi F, Anguela XM, Pavani G, et al. Overcoming preexisting humor-al immunity to AAV using capsid decoys. Sci Transl Med. 2013;5:194ra92. doi: 10.1126/scitranslmed.3005795.
75. Wright JF, Zelenaia O. Vector characterization methods for quality con-trol testing of recombinant adeno-associated viruses. Methods Mol Biol. 2011;737:247–278. doi: 10.1007/978-1-61779-095-9_11.
76. Wright JF. Adeno-associated viral vector manufacturing: keeping pace with accelerating clinical development. Hum Gene Ther. 2011;22:913– 914. doi: 10.1089/hum.2011.2514.
77. Li H, Malani N, Hamilton SR, Schlachterman A, Bussadori G, Edmonson SE, Shah R, Arruda VR, Mingozzi F, Wright JF, Bushman FD, High KA. Assessing the potential for AAV vector genotoxicity in a murine model.
Blood. 2011;117:3311–3319. doi: 10.1182/blood-2010-08-302729. 78. Sharma A, Easow M, Sriganesh V, Neely J, Kalipatnapu S. Gene therapy
for haemophilia. Cochrane Database Syst Rev. 2014;1–15.
79. Büller HR, Bethune C, Bhanot S, Gailani D, Monia BP, Raskob GE, Segers A, Verhamme P, Weitz JI; FXI-ASO TKA Investigators. Factor XI antisense oligonucleotide for prevention of venous thrombosis. N Engl J Med. 2015;372:232–240. doi: 10.1056/NEJMoa1405760.
80. Sehgal A, Barros S, Ivanciu L, et al. An RNAi therapeutic targeting anti-thrombin to rebalance the coagulation system and promote hemostasis in hemophilia. Nat Med. 2015;21:492–497. doi: 10.1038/nm.3847.