CHARACTERIZATION OF THE ROLE OF PSEUDOMONAS SYRINGAE TYPE III EFFECTOR HopAF1 IN VIRULENCE
Erica Jania Washington
A dissertation submitted to the faculty of the University of North Carolina at Chapel Hill in partial fulfillment of the requirements for the degree of Doctor of Philosophy in the
Department of Biology.
Chapel Hill 2013
Approved by: Jeff Dangl
© 2013
ABSTRACT
Erica Jania Washington: Characterization of the role of Pseudomonas syringae type III effector HopAF1 in virulence
(Under the direction of Jeff Dangl)
Many plant pathogens, including Pseudomonas syringae, encode the type III secretion system for translocating effector proteins into the host during infection. Strains of P. syringae which are not capable of delivering the type III effectors are nonpathogenic. Therefore, the functions of type III effectors are essential for disease. Although several type III effectors have been demonstrated to block components of the plant defense response, the functions of most type III effectors are unknown. Our lab is interested in the type III effector HopAF1, a type III effector that is present in eleven of the nineteen sequenced strains of P. syringae and other plant pathogens. Although the presence of HopAF1 in multiple strains of P. syringae suggests that it plays an important role in virulence, no
function has yet been associated with HopAF1. We generated a tertiary structural prediction for HopAF1, which suggests that HopAF1 is structurally related to bacterial deamidases. Deamidation, the irreversible substitution of an amide group with a carboxylate group, is the mechanism by which several bacterial virulence factors manipulate the activity of a specific substrate. To identify a potential target of HopAF1 activity, we employed a yeast two-hybrid screen and identified Arabidopsis methylthioadenosine nucleosidase (MTN1) as a putative target of HopAF1. This interaction, which we extended to include Arabidopsis MTN2, was confirmed in planta using coimmunoprecipitations and bimolecular fluorescence
levels of ethylene biosynthesis. Ethylene is a key plant hormone for plant developmental processes, such as senescence and flowering. Ethylene is also induced during PTI. Therefore, we hypothesized that HopAF1 inhibits PTI by manipulating MTNs and levels of ethylene in plants. To this end, we used gas chromatography to measure ethylene
biosynthesis in plants treated with bacterially-delivered HopAF1. We determined that
HopAF1 inhibits ethylene biosynthesis in a manner dependent on putative catalytic residues. Additionally, Yang cycle mutants mtn1 mtn2 and mtk are more susceptible to weak
ACKNOWLEDGEMENTS
I am grateful to many colleagues, friends, and family members for the support and encouragement that have made this dissertation possible. It would be impossible to
adequately thank each individual; therefore I would like to specifically thank a few individuals for their contributions to my graduate school career.
First, I would like to thank my advisor, Jeff Dangl, for his ongoing support and enthusiasm. Jeff taught me how to fearlessly follow science where it takes you and in doing so, always provided me with the resources necessary to help my project progress. In
addition, Jeff took the time to teach me how to think scientifically and critically. I also strongly believe that without Jeff’s support during a critical time, I would not be able to write this dissertation. Finally, I would like to thank Jeff for providing me a home in the Dangl-Grant lab.
During my time in the Dangl-Grant lab I have worked with so many wonderful individuals that taught me about science and life. I would like to thank Sarah Grant for the time she invested in supporting me and my research. I would also like to thank Petra Epple for her guidance, Terry Law for her friendship, Marc Nishimura for mentoring me when I first joined the lab and Nuria Sanchez Coll for being a role model and a friend. I would also like to thank Vera Bonardi, Tatiana Mucyn, and Derek Lundberg for many good times over the years.
addition to spending many hours at the microscope with me, Tony has always been
generous with his encouragement and support. Thank you to Ashutosh Tripathy of the UNC Macromolecular Interactions Facility, Brenda Temple of the UNC Center for Structural Biology, Lee Graves and John Sondek.
Thank you to my committee for your patience as I developed as a scientist. Thanks to Miriam Braunstein and Matt Wolfgang for teaching me bacteriology in my first year at UNC and for continuing to support me while on my committee. I would also like to Thank you to Jason Reed for being thoughtful and constructive during committee meetings and journal clubs. I would also like to thank Joe Kieber. I rotated in the Kieber lab and always felt like an honorary member of the Kieber lab. The Kieber lab doors remained open to me when I needed to use the gas chromatography machine to do some of my most important experiments. Thank you to Gyeong Mee Yoon and Smadar Harpaz-Saad for being so patient and kind while teaching me how to perform these experiments.
I would also like to thank Sharon Milgram, who recruited me to UNC through the Interdisciplinary Biomedical Sciences program. Sharon was really invested in helping me succeed as a person and a scientist. I particularly want to thank her for support in regards to my participation in the Cell and Molecular Biology training grant program.
I would not be the person or the scientist I am today without my best friend and my love, David Hubert. I want to thank David for his unconditional love and understanding.
Finally, I would like to thank my family. First, I want to thank my big brother, Derek, for giving me brotherly advice from across the country and reminding me of what it means to lead a well-balanced and successful life. My parents have given me the best parental
TABLE OF CONTENTS
LIST OF TABLES ... x
LIST OF FIGURES ... xi
LIST OF ABBREVIATIONS ... xiii
Chapter 1 Introduction ... 20
A two-tiered model of the plant innate immune system. ... 21
The phytohormone ethylene plays multiple roles in the plant defense response. ... 26
Successful pathogens suppress PTI /ETI activation and signaling. ... 28
The type III secretion system and its effectors are the main virulence determinants of Pseudomonas syringae. ... 28
References ... 39
Chapter 2 Bacterial virulence factors modulate eukaryotic host cell signaling systems via deamidation ... 50
Preface ... 50
Abstract ... 50
Introduction ... 51
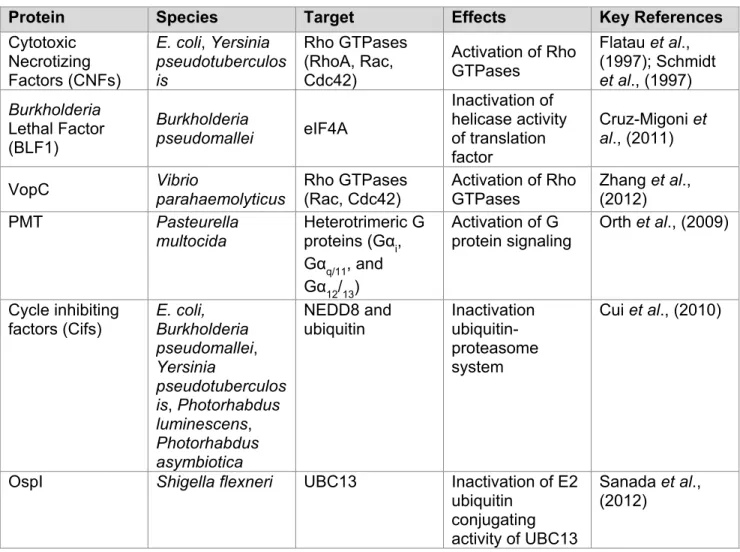
Cytotoxic Necrotizing Factor Toxins ... 53
Burkholderia Lethal Factor 1 inhibits activity of translation factor eIF4A. ... 56
Vibrio parahaemolyticus type III effector VopC deamidates small GTPases. ... 57
Pasteurella multocida toxin ... 58
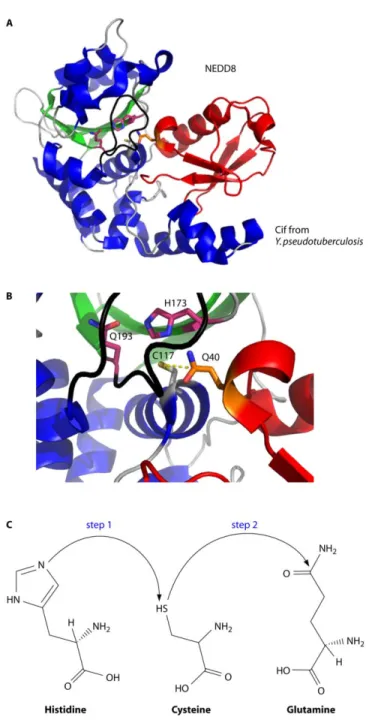
Cycle Inhibiting Factors ... 61
References ... 81
Chapter 3 P. syringae type III effector HopAF1 suppresses plant immunity by targeting the methionine recycling pathway ... 90
Preface ... 90
Abstract ... 91
Introduction ... 92
Results ... 95
Discussion ... 105
Materials and Methods ... 109
Figures ... 118
References ... 135
Chapter 4 Conclusions and Future Directions ... 142
Using a shotgun approach and mass spectrometry to determine if HopAF1 modulates MTN function via deamidation. ... 143
Determine whether a functional deamidation site variant of MTN1 is impervious to HopAF1 activity in planta. ... 145
Generate a HopAF1/MTN1 co-crystal to determine the molecular mechanism behind HopAF1 inhibition of MTN activity. ... 147
Figures ... 150
LIST OF TABLES
Table 1.1 P. syringae type III effectors: defense suppression, localization,
activities and targets ... 38
LIST OF FIGURES
Figure 1.1 P. syringae type III effectors target multiple host cellular components. ... 37
Figure 2.1 A schematic representation of enzymatic deamidation in proteins. ... 72
Figure 2.2 The catalytic domains of diverse deamidases. ... 73
Figure 2.3 Crystal structure of the C-terminal region of Pasteurella multocida toxin. ... 74
Figure 2.4 PMT deamidates and activates heterotrimeric Gα proteins. ... 75
Figure 2.5 Model of Cif inactivation of CRL activity and ubiquitination. ... 76
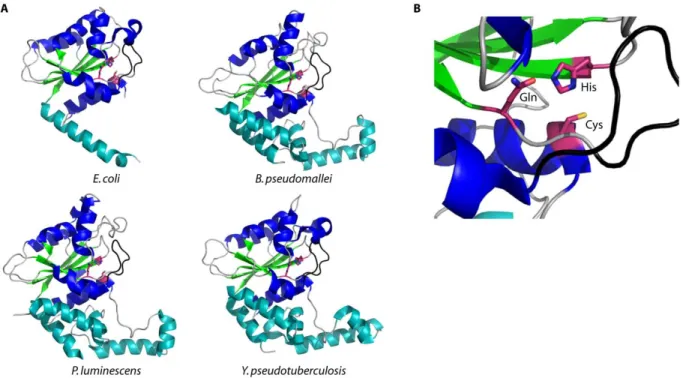
Figure 2.6 Crystal structures of cell cycle-inhibiting factors. ... 77
Figure 2.7 Crystal structure of the CifYp deamidase/NEDD8 complex. ... 78
Figure 2.8 Crystal structures of unbound OspI and the OspI/Ubc13 complex. ... 79
Figure 3.1 The predicted structure of HopAF1 exhibits structural homology to CheD, a bacterial deamidase. ... 118
Figure 3.2 Alignment of HopAF1 sequences from P. syringae pathovars and other plant pathogens. ... 119
Figure 3.3 HopAF1 transgenic lines are susceptible to weak pathogens. ... 120
Figure 3.4 Arabidopsis transgenic lines expressing wild-type HopAF1 inhibit flg22-mediated protection against P. syringae pv. tomato DC3000. ... 121
Figure 3.5 HR triggered by AvrXv3 is dependent on putative catalytic residues. ... 122
Figure 3.6 Establishing a system to study localization of transiently expressed HopAF1 in N. benthamiana. ... 123
Figure 3.7 HopAF1 is targeted to the plasma membrane via acylation. ... 124
Figure 3.8 The acylation-minus variant of HopAF1 co-localizes with free YFP, a soluble protein. ... 125
Figure 3.9 The Yang cycle contributes to ethylene biosynthesis, which is suppressed by Pto DC3000. ... 126
Figure 3.10 HopAF1 associates with Arabidopsis MTN proteins at the plasma membrane ... 127
Figure 3.12 HopAF1 inhibits PAMP-induced ethylene biosynthesis. ... 129
Figure 3.13 Pfo-1 is unable to induce high levels of ethylene biosynthesis
in Yang cycle mutants, mtn1 mtn2 and mtk. ... 130
Figure 3.14 HopAF1 domain architecture and circular dichroism spectra of HopAF1,
HopAF1H186A, and MTN1. ... 131
Figure 3.15 HopAF1 inhibits MTN1 activity in vitro in a manner dependent on catalytic activity. ... 132
Figure 3.16 The Yang cycle is required for PTI in Arabidopsis. ... 133
Figure 3.17 Proposed model of HopAF1 suppression of plant innate immunity. ... 134
Figure 4.1 Alignment and structural localization of MTN1 conserved
putative deamidase target residues. ... 150
Figure 4.2 MTN1 and MTN2 variants display loss-of-function phenotypes
when residues N113 and N194 are ‘deamidated’. ... 151
Figure 4.3 Circular dichroism spectra of MTN1 ‘deamidated’ variants does not
vary from wild-type MTN1. ... 152
Figure 4.4 Purification of wild-type MTN1. ... 153
LIST OF ABBREVIATIONS
A/E attaching and effacing
ABA abscisic acid
ACS aminocyclopropane carboxylate synthase
ADP adenosine diphosphate
ADR1 activated disease resistance 1
AMP adenosine monophosphate
APX ascorbate peroxidase
ARF-GEF ADP-ribosylation factor guanine nucleotide exchange factor
ATP adenosine triphosphate
avr avirulence
BAK1 bri1-associated receptor kinase 1
BCL-10 B-cell lymphoma 10
BiFC bimolecular fluorescence complementation
BIK1 Botrytis-induced kinase 1
BKK1 BAK1-like kinase 1
BLF1 Burkholderia Lethal Factor 1
BRI1 brassinosteroid insensitive 1
C caryboxyl
CaCo2 human colon carcinoma cell line
cAMP cyclic adenosine monophosphate
CARD caspase recruitment domain
CATERPILLER CARD transcription enhancer, purine binding, pyrine, lots of leucine repeats
CBL3 calcineurin B-like protein 3
CD circular dichroism
Cdc42 cell division control protein 42
CDK cyclin dependent kinase
cDNA complementary DNA
Cdt1 cdc10-dependent transcript 1
CeBIP chitin elicitor binding protein
CERK1 chitin elicitor receptor kinase 1
cfu colony forming units
CHBP Cif homolog Burkholderia pseudomallei
CHPA Cif homolog Photorhabdus asymbiotica
CHPL Cif homolog Photorhabdus luminescens
CHYP Cif homolog Yersinia pseudotuberculosis
Cif cell inhibiting factor
CNF1 cytotoxic necrotizing factor 1
CNF2 cytotoxic necrotizing factor 2
CNF3 cytotoxic necrotizing factor 3
CNFY cytotoxic necrotizing factor isolated from Yersinia
Col-0 Arabidopsis ecotype Col-0
COP9 constitutive photomorphogenesis
CRL cullin ring ligases
CSN COP9 signalosome
cYFP carboxyl terminus of YFP
DAG diacylglycerol
DNA deoxyribonucleic acid
DNT dermonecrotic toxin
EDTA ethylenediamine tetraacetic acid
EFR elongation factor Tu receptor
EF-Tu elongation factor Tu
EHEC enterohemorrhagic E. coli
eIF4A eukaryotic initiation factor 4A
EIL1 ethylene insensitive 3-like 1
EIL2 ethylene insensitive 3-like 2
EIN2 ethylene insensitive 2
EIN3 ethylene insensitive 3
elf18 18 amino acid peptide derived from elongation factor Tu
EPEC enteropathogenic E. coli
ERF ethylene response factor
ETI effector-triggered immunity
ETS effector-triggered susceptibility
Fla 216 Florida 216
Fla 7060 Florida 7060
flg22 22 amino acid peptide derived from flagellin
FLS2 flagellin sensing 2
GDP guanosine diphosphate
GmHID1 2-hydroxyisoflavanone dehydratase from Glycine Max
GMP guanosine monophosphate
GRP7 glycine-rich RNA binding protein 7
GRP8 glycine-rich RNA binding protein 8
GR-RBP glycine-rich RNA binding protein
GTP guanosine triphosphate
HeLa cell line derived from Henrietta Lacks
Hop Hrp-dependent outer protein
HR hypersensitivity response
hrc HR and conserved genes
hrcC HR and conserved protein C; structural type III secretion system protein
hrp HR and pathogenicity genes
HrpL HR and pathogenicity protein L; alternative sigma factor
Hsp70 heat shock protein 70
Hsp90 heat shock protein 90
IPTG isopropyl β-D-1-thiogalactopyranoside
JA jasmonic acid
JAK/STAT Janus kinase/ signal transducer and activator of transcription
kb kilobase
LEE locus of enterocyte effacement
LPS lipopolysaccharide
LRR leucine-rich repeat
LysM lysin motif
MALTI mucosa-associated-lymphoid-tissue-lymphoma-translocation gene 1
MAMP microbe-associate molecular pattern
MAPK mitogen-activated kinase
MBP maltose binding protein
MES 2-(4-morpholino)ethanesulfonic acid
MTA methylthioadenosine
MTK methylthioribose kinase
N amino
NADPH nicotinamide adenine dinucleotide phosphate
NB nucleotide-binding
NEDD8 neural precursor cell expressed, developmentally down-regulated 8
NLR nucleotide-binding domain and leucine-rich repeat-containing
NOD nucleotide-binding oligomerization domain
nYFP amino terminus of YFP
OD optical density
ORF open reading frame
PBL PBS1-like protein
PBS1 AvrPphB susceptible 1
PDB protein databank
PEPR1 Pep1 receptor 1
Pfo-1 Pseudomonas fluorescens
PI3K phosphoinositide-3-kinase
PLC2 phospholipase C 2
PLCβ phospholipase C β
PMT Pasteurella multocida toxin
PR pathogenesis-related
PRR pattern recognition receptor
PsbQ oxygen-evolving enhancer protein 3
PTI PAMP-triggered immunity
Pto DC3000 Pseudomonas syringae pv. tomato DC3000
pv pathovar
Rac Ras-related C3 botulinum toxin substrate 1
RbohD respiratory burst oxidase homolog D
Rbx RING box protein 1
RhoA Ras homology A
RIN4 RPM1-interacting protein 4
RING really interesting new gene 1
RIPK RPM1-induced protein kinase 1
RLK receptor-like kinase
RLP receptor-like protein
rmsd root-mean-square deviation
RNA ribonucleic acid
ROS reactive oxygen species
rpm revolutions per minute
RPM1 resistance to Pseudomonas syringae pv. maculicola 1
RPS2 resistance to Pseudomonas syringae pv. syringae 2
RPS5 resistance to Pseudomonas syringae pv. syringae 5
SA salicylic acid
SID2 salicylic acid induction deficient 2
STAND signal transduction ATPases with numerous domains
T3E type III effector
T3SS type III secretion system
TAB1 TAK1-binding protein 1
TAB2 TAK1-binding protein 2
TAK1 TGFβ-activated kinase 1
TEV tobacco etch virus
TIR Toll, interleukin-1 receptor
TTSS1 type III secretion system 1
TTSS2 type III secretion system 2
UBC13 ubiquitin-conjugating enzyme 13
UBL ubiquitin-like protein
UEV1A ubiquitin-conjugating enzyme variant 1a
WB western blot
Xcv Xanthomonas campestris pv. vesicatoria
YFP yellow fluorescent protein
Chapter 1 Introduction
Plants constantly encounter pathogens such as insects, oomycetes, viruses, fungi, and bacteria in their natural habitats. However, many of these plant-microbe interactions do not result in disease. Plant species may be resistant to microbes that are successful
pathogens on other plant species. This is called non-host resistance, and can be thought of as reflecting situations where the microbe in question is not evolutionarily adapted to life on that plant species (Senthil-Kumar and Mysore, 2013). Many mechanisms of non-host resistance involve physical barriers that prevent the pathogen from accessing nutrients in the plant apoplastic fluid. The waxy cuticle layer, the epidermis, and the plant cell wall are structural barriers common in many plants. Bacterial pathogens may access the apoplastic fluid through the stomatal openings in the plant leaf surface (Senthil-Kumar and Mysore, 2013). However, another mechanism of non-host resistance is the closure of stomata in the presence of microbes (Melotto et al., 2006). Moreover, the presence of non-host microbes also induces reinforcements in the cell wall in the form of callose, lignin, or suberin near sites of infection (Senthil-Kumar and Mysore, 2013).
Once microbes reach the apoplastic fluid, antimicrobials prevent further colonization. Non-host Arabidopsis apoplastic fluid extracts can restrict the growth of P. syringae
A two-tiered model of the plant innate immune system.
Adapted microbes successful at surpassing the passive non-host immunity detailed above must overcome an active two-tiered plant immune system (Jones and Dangl, 2006). The first stage is PTI or PAMP-triggered immunity. As a key virulence strategy, pathogens have evolved effector molecules, such as type III effector (T3E) proteins in P. syringae and RXLR effectors in oomycetes, that target PTI, resulting in disease and effector-triggered susceptibility (ETS) (Jones and Dangl, 2006) (Table 1.1). Plants evolved the second tier of the immunity termed ETI for effector-triggered immunity. In ETI, plants use NLR (nucleotide-binding domain and leucine-rich repeat-containing) proteins to detect effectors, often triggering a rapid and localized cell death response called HR.
PAMP-triggered immunity
The first line of active defense begins with transmembrane pattern recognition receptors (PRRs) recognizing conserved molecular patterns from pathogens called pathogen-associated molecular patterns or PAMPs (Boller and Felix, 2009). Since these microbial signatures are not limited to pathogenic microbes, they are perhaps more aptly referred to as microbial-associated molecular patterns or MAMPs. For the purposes of this introduction, we will use the terms PAMPs and PTI. PAMPs are often highly conserved across a wide range of microbes, fulfill a function essential to the pathogen’s lifecycle, and yet are not present in the host (Boller and Felix, 2009). Common bacterial PAMPs are lipopolysaccharide (LPS), peptidoglycan, elongation factor Tu (EF-Tu), and flagellin (Boller and Felix, 2009). Common fungal PAMPs include chitin and ergosterol, both derived from cell walls of fungi (Boller and Felix, 2009). A 22 amino acid peptide derived from a
PAMP recognition occurs via pattern recognition receptors (PRRs) that are
analogous to Toll-like receptors in animals. PRRs are either receptor-like kinases (RLKs) or receptor-like proteins (RLPs). RLKs contain an N-terminal extracellular domain, a single transmembrane domain, and a C-terminal kinase domain. RLPs contain an N-terminal extracellular domain, a single transmembrane domain, and a C-terminal short cytoplasmic tail. Some well-studied PRRs include FLS2, EFR, and CERK1 (Monaghan and Zipfel, 2012). FLS2 was the first PRR to be discovered and as a result, much of what we know about PAMP recognition stems from FLS2 studies (Gomez-Gomez and Boller, 2000). FLS2 is the RLK that recognizes flg22 (Gomez-Gomez and Boller, 2000). The C-terminal kinase domain of FLS2 is a Ser/Thr kinase, except that its catalytic loop contains a CD sequence instead of RD. For these reason, FLS2 is a non-RD kinase, which appears to be a hallmark of the kinase domain of immune system related PRRs (Dardick and Ronald, 2006). EFR is a Brassicaceae-specific RLK PRR that recognizes elf18 (Zipfel et al., 2006). CERK1, a RLK that recognizes fungal chitin and bacterial peptidoglycan in Arabidopsis, has three
extracellular LysM domains for carbohydrate binding, along with the transmembrane domain and the C-terminal kinase domain (Miya et al., 2007; Wan et al., 2008). In rice, chitin
recognition requires an additional RLP PRR called CeBiP (Kaku et al., 2006). CeBiP
interacts with rice CERK1 and this interaction is enhanced in the presence of chitin (Shimizu et al., 2010).
2010; Yamaguchi et al., 2006) Heterocomplex formation between the BAK1 and PRRs leads to trans-phosphorylation and signal transduction (Schulze et al., 2010). Schulze et al., demonstrated that triggering downstream flg22-mediated responses requires BAK1 with a functional kinase domain (Schulze et al., 2010). BAK1 is critical to PTI-related signaling because bak1 mutants have severely reduced responses to flg22 and are completely
insensitive to elf18 (Chinchilla et al., 2007). BAK1 is also required for responses triggered by other PAMPs such as LPS and peptidoglycan (Shan et al., 2000). However, not all PTI signaling requires BAK1. For example, instead of interacting with BAK1, CERK1 undergoes ligand-induced homo-dimerization leading to activation of downstream immune responses (Liu et al., 2012).
Perception of PAMPs triggers PTI (Jones and Dangl, 2006). PTI is a critical
component of the two-tiered immune system because plants unable to trigger this response are more susceptible to infection. For example, Arabidopsis and N. benthamiana plants encoding a non-functional fls2 are more susceptible to bacterial infection (de Torres et al., 2006; Hann and Rathjen, 2007; Li et al., 2005; Zipfel et al., 2004). Conversely, ectopic expression of EFR in non-Brassicaceae plants Nicotiana benthamiana and tomato increases their resistance to a range of bacterial pathogens (Lacombe et al., 2010).
The timing of PTI responses after PAMP recognition ranges from seconds to hours. Early PTI responses include an influx in ions such as H+, K+, Cl- and Ca2+. Ca2+ activates calcium-dependent protein kinases and RbohD (Ma and Berkowitz, 2007). RbohD is a NADPH oxidase that contributes to reactive oxygen species (ROS) burst in Arabidopsis (Ogasawara et al., 2008). This ROS burst occurs approximately 2 minutes after PAMP recognition. ROS burst contributes to PTI by activating defense gene expression,
strengthening the cell wall, and initiating cell death (Torres et al., 2006). PTI responses also include the activation of mitogen-activated kinases (MAPKs), which in turns leads to
a fraction of the massive reprogramming of gene expression that occurs after PAMP
recognition (Zipfel et al., 2006; Zipfel et al., 2004). Later PTI responses include the induction of ethylene biosynthesis and callose deposition. Callose deposition, an easily measured output and classic PTI response, consists of an accumulation of β-1,3-glucan polymers in the plant cell wall near the sight of infection to form physical barriers against further pathogen attack. We will address the role of ethylene further in the next section of this introduction.
Effector-triggered immunity
The second line of defense is the gene-for-gene recognition of pathogen effectors by plant disease resistance gene products. This line of defense evolved due to the ability of virulent pathogens to inhibit PTI and thus render the plant susceptible to infection. This condition is termed effector-triggered susceptibility or ETS (Jones and Dangl, 2006). Plants evolved disease resistance (R) proteins to combat ETS. Disease resistance proteins are often referred to as NB-LRR, or NLR, proteins due to their domain architecture, which
consists of a central nucleotide-binding (NB) domain and highly polymorphic C-terminal LRR region. The N-terminal domains of NLRs commonly consist of either a TIR (Toll, interleukin-1 receptor) domain or a CC (coiled-coil) domain. There are approximately interleukin-150 NLRs in the Arabidopsis Col-0 genome (Meyers et al., 2003).
NLRs can detect effectors from a wide variety of plant pathogens including fungi, oomycetes, viruses, and bacteria. NLR proteins may detect effectors directly or indirectly. For example, AvrPita, an effector protein from Magnaporthe grisea, interacts directly with the NLR Pi-ta from rice in yeast and in vitro (Jia et al., 2000). The indirect detection of an
hypothesis implies that multiple effectors modify the same host target in different ways and that the host target can guarded by multiple NLRs (Jones and Dangl, 2006). This is because ETI is a driving force of the host-pathogen arms race and is consistent with the finding that pathogen effectors converge on a subset of host proteins (Mukhtar et al., 2011). A classic example of the guard hypothesis that fulfills these tenets is the targeting of the Arabidopsis protein RIN4 (RPM1-Interacting Protein 4) by multiple P. syringae T3Es. AvrRpt2-mediated cleavage of RIN4 and phosphorylation of RIN4 in the presence of AvrRpm1 and AvrB, trigger the activation of NLRs RPS2 and RPM1, respectively (Axtell and Staskawicz, 2003; Mackey et al., 2003; Mackey et al., 2002).
The molecular mechanisms that govern NLR activation and signaling are not fully understood and there are few generalizable themes describing NLR activation (Bonardi et al., 2012; Bonardi and Dangl, 2012; Eitas and Dangl, 2010). This is likely due to their rapid evolution in the arms race and the ability to detect virulence effectors from a diverse range of plant pathogens. Plant NLRs are similar to CATERPILLER/NOD/LRR proteins and STAND ATPases (Leipe et al., 2004; Ting and Davis, 2005). STAND ATPases are
molecular switches that cycle between an ADP-bound off state and an ATP-bound on state. Similarly, NLRs contain a critical catalytic sequence, called the P loop, in their nucleotide-binding domain. The P loop domain is not required for the function of all NLRs. For example, NLR activation, basal resistance to virulent pathogens, and salicylic acid accumulation mediated by the ADR1 family of NLRs does not require P loop function (Bonardi et al., 2011). Proper activation of NLRs requires a conformational change to release the intramolecular interactions. Additionally, homotypic or heterotypic interactions or the
infection called the hypersensitivity response or HR often occurs once ETI is triggered (Jones and Dangl, 2006). Although the exact mechanism by which ETI restricts pathogen growth is unknown, mutation of R genes or in genes encoding chaperones required for NLR stability results in an increase in susceptibility to pathogens.
The phytohormone ethylene plays multiple roles in the plant defense response.
There are three main hormones in plants that regulate the plant defense response against abiotic and biotic stresses: salicylic acid (SA), jasmonic acid (JA), and ethylene. The simplest model of how these hormones function states that ethylene and JA mediate
resistance to necrotrophs and herbivorous insects, while SA mediates resistance to biotrophs (Robert-Seilaniantz et al., 2007). Although there are a few exceptions, the ethylene and JA pathways are synergistic to each other and antagonistic to SA (Ecker and Davis, 1987; Penninckx et al., 1998; Schenk et al., 2000). There is cross-talk between these pathways to allow the plant to fine tune its defense response against the multiple invaders it may encounter.
In recent years, there have been numerous examples of plant pathogens that hijack specific hormone-regulated signaling pathways, supporting the hypothesis that
campestris pv. vesicatoria, (Xcv) was recently found to target a tomato ethylene response factor (ERF) called SIERF4. SIERF4 mutants display reduced ethylene levels and an
increase in susceptibility to Xcv (Kim et al., 2013), supporting the hypothesis that ethylene is involved in plant defense.
Ethylene is a gaseous plant hormone that is required for normal plant development processes such as seed germination, leaf senescence, and leaf and petal abscission (Kieber, 1997). As mentioned above, ethylene biosynthesis is induced during the plant defense response. This occurs after recognition of PAMPs and the induction of the MAPK signaling cascade. Phosphorylation stabilizes ACS (aminocyclopropane carboxylate
synthase) proteins and thus increases the biosynthesis of ethylene (Liu and Zhang, 2004). There is a growing body of work focused on elucidating the role of ethylene in defense against pathogens. Ethylene up-regulates the expression of pathogenesis-related (PR) proteins and increases the accumulation of hydroxyproline-rich, cell-wall strengthening proteins (Ecker and Davis, 1987; Penninckx et al., 1998). Ethylene also increases SA-induced expression of PR-1. This suggests that in addition to functioning synergistically with JA, ethylene can also support SA-dependent defense responses (Lawton et al., 1994). Furthermore, ethylene directly up-regulates the expression of FLS2. FLS2 transcription is directly controlled by the binding of ERFs EIN3/EIL1/EIL2 to the FLS2 promoter and in ethylene insensitive mutants (ein2), both FLS2 expression and FLS2 protein accumulation are suppressed. As a result, ein2 plants are more susceptible to weakened strains of P. syringae (Boutrot et al., 2010; Mersmann et al., 2010).
promotes susceptibility to P. syringae (Chen et al., 2009). Furthermore, ethylene perception is also necessary for manifestation of disease symptoms, following Pto DC3000 infection in Arabidopsis (Bent et al., 1992). In conclusion, the role of ethylene in plant defense signaling pathways is complex and needs more attention. Further experimentation is required for complete understanding of how all phytohormones regulate defense pathways and how pathogens promote disease by disrupting hormone homeostasis.
Successful pathogens suppress PTI /ETI activation and signaling.
Well-adapted pathogens have evolved methods to breach the multiple lines of plant defense described above. Pathogens employ effectors to block components of PTI, ETI, or both. Fungi and oomycetes encode hundreds of effectors as part of their virulence
strategies. However, little is known about their function and mechanisms by which they thwart plant immune systems. For that reason, in this section, we will mainly focus on T3Es from bacterial phytopathogens, mostly from Pseudomonas syringae. Bacterial pathogen strains can each inject approximately 20 to 30 virulence effectors into the host cell during infections, and strains across one widespread species, Pseudomonas syringae deploy approximately 60 protein families, many of which have highly variable members (Baltrus et al., 2011) (Figure 1.1; Table 1.1).
The type III secretion system and its effectors are the main virulence determinants of
Pseudomonas syringae.
Before detailing how T3Es target various defense machinery in the plant cell to suppress immune system input signaling and outputs responses, it is important to digress to first introduce the type III secretion system. This will be followed by a detailed discussion of T3E functions, largely in the context of the plant pathogen P. syringae.
The type III secretion system
disrupt normal host processes and defense responses. One widespread and critical bacterial virulence mechanism is the type III secretion system (T3SS), which consists of a multi-ring base located in the bacterial inner and outer membranes and a protruding needle-like appendage. This needle-needle-like pilus is called the Hrp pilus in phytopathogenic bacteria. The Hrp pilus is used to deliver T3Es into the host cell to cause disease. The T3SS is conserved across a broad range of Gram-negative pathogens including animal pathogens such as Salmonella, Yersinia pestis, and Escherichia coli and phytopathogens
Pseudomonas syringae, Ralstonia solanacearum and various Xanthomonas species (Chatterjee et al., 2013; Deslandes and Rivas, 2012; Raymond et al., 2013).
Arabidopsis-P. syringae model system
P. syringae causes disease in agronomically important crops such as tomatoes, tobacco, and beans. P. syringae also infects Arabidopsis thaliana, a genetically tractable model plant system (Whalen et al., 1991). Since P. syringae was also considered to be genetically tractable, this established a new plant-pathogen system for studying molecular mechanisms utilized by pathogens to causes disease progression and the subsequent host defense responses.
(hypersensitivity response and conserved) genes, is up-regulated. Although the specific in planta signal that is responsible for up-regulating the hrp and hrc genes is unknown, Huynh et al., demonstrated that fructose, sucrose, and mannitol upregulate AvrB transcription in defined media (Huynh et al., 1989). Although P. syringae remains extracellular, the T3SS translocates T3Es into the host cell to suppress components of PTI and ETI and contribute to pathogenicity (Grant et al., 2006). P. syringae type III secretion system mutants are nonpathogenic; therefore the type III secretion system and its effectors are required for the pathogenicity of P. syringae (Lindgren et al., 1986). Furthermore, Lindgren et al., suggested that these same effectors are responsible for HR triggered in non-host plants (Lindgren et al., 1986).
Bacterial genomics, next generation sequencing, and definition of T3E families
The effort to identify T3E families in all strains of P. syringae is ongoing and has involved many approaches over the years. Scoring robust effector loss-of-function phenotypes using transposon mutagenesis screens is difficult because, due to functional redundancy of sequence un-related T3Es, there is often only a weak, or even undetectable, contribution of any single effector to virulence. Therefore, identification of T3Es has relied heavily on bioinformatics-based methods. Research on the molecular mechanisms of P. syringae T3Es has focused on the three ‘gold standard’ P. syringae strains that were genome sequenced first; Pto DC3000, P. syringae pv. syringae B728A, and P. syringae pv. phaseolicola 1448A (Buell et al., 2003; Feil et al., 2005; Joardar et al., 2005).
1448A (Chang et al., 2005). This was followed by validation of the translocation of the candidate effector proteins (Chang et al., 2005).
Next generation DNA sequencing has made it possible to quickly generate data on multiple, diverse P. syringae genomes. Using a combination of Illumina and 454 platforms, Baltrus et al., generated draft genome sequences for 14 previously unsequenced,
phylogenetically diverse strains of P. syringae (Baltrus et al., 2011). The search for type III effectors among these genomes led to the identification of eight new type III effectors families, resulting in a total of approximately 60 T3E families in this diverse collection of P. syringae strains that expands the five distinct phylogroups (Baltrus et al., 2011).
Effectors that target PTI
Multiple effectors target RLKs at the plasma membrane and downstream
components of PTI signaling pathways. AvrPto and AvrPtoB share no primary or structural homology, but they are redundant P. syringae T3Es that suppress the function of RLKs. AvrPtoB has an N-terminal kinase domain that is sufficient to suppress flg22-induced PTI (Gohre et al., 2008). The C-terminus of AvrPtoB is an E3 ubiquitin ligase domain that can ubiquitinate and degrade RLKs FLS2 and CERK1 (Gimenez-Ibanez et al., 2009; Gohre et al., 2008). AvrPto can bind and inhibit the activity of FLS2 and EFR (Xiang et al., 2008). Although, it has been reported that AvrPto and AvrPtoB can inhibit BAK1 function, there is conflicting evidence for this interaction (Shan et al., 2008; Xiang et al., 2011). AvrPphB cleaves kinases, such as BIK1, that function downstream of PAMP recognition and integrate signals from multiple PRRs to block PTI responses (Zhang et al., 2010). Interestingly,
phosphothreonine lysase activity to inhibit the function of MPK3 and MPK6 (Zhang et al., 2007). Similar to HopF2, disruption of PTI by targeting MAPK cascades triggers activation of a NLR and ETI (Zhang et al., 2012).
Using effectors as probes of plant cell biology and plant defense has led to the identification of components of the plant immune system that extend beyond PRRs and NLRs. HopM1 targets vesicle trafficking by inhibiting the function of an Arabidopsis ARF-GEF protein MIN7. HopM1 degradation of MIN7 is proteasome-dependent (Nomura et al., 2006). Interestingly, when ETI is triggered by AvrRpt2, AvrPphB, and HopA1, HopM1-mediated destabilization of MIN7 is inhibited (Nomura et al., 2011). This demonstrates a novel situation in which ETI contributes to plant defense by preventing bacterial suppression of a key component of PTI (Nomura et al., 2011). HopU1 is a P. syringae type III effector mono-ADP-ribosyltransferase that targets Arabidopsis glycine-rich RNA binding proteins (GR-RBPs) GRP7 and GRP8 (Fu et al., 2007). HopU1 negatively affects the ability of GRP7 to bind RNA (Jeong et al., 2011). Specifically, HopU1 blocks the interaction between GRP7 and FLS2 and EFR transcripts (Nicaise et al., 2013), leading to a reduction in FLS2
accumulation after infection with a virulent P. syringae strain expressing HopU1 (Nicaise et al., 2013). This is consistent with HopU1 inhibiting PAMP responses triggered by flg22, such as callose deposition (Fu et al., 2007). T3Es HopZ1a and HopZ1b have also been
implicated in PTI suppression. HopZ1a and HopZ1b degrade the soybean protein GmHID1 (2-hydroxyisoflavanone dehydratase), a protein involved in the biosynthesis of antimicrobial phytoalexins (Zhou et al., 2011). Soybean plants with GmHID1 silenced are more
in PAMP-induced ROS burst and callose deposition (Rodriguez-Herva et al., 2012). Finally, HopG1 also inhibits callose and ROS burst, although the molecular mechanism is currently unknown (Block et al., 2010).
Effectors that block ETI
The first evidence of type III effectors blocking HR came from studies in the 1990s in which Xanthomonas campestris pv. campestris hrp mutants induced a programmed cell death in the vascular system of crucifers (Jackson et al., 1999). This suggested that there are type III effectors in the hrp regulon that are blocking this HR-like response. More recent studies have identified P. syringae type III effectors that are involved in suppressing ETI. For example, Jackson et al. demonstrated that VirPphA, AvrPphC, and AvrPphF from P.
syringae pv. phaseolicola were able to inhibit NLR activation (Jackson et al., 1999). AvrPtoB suppresses ETI-associated programmed cell death in tomato plants via ubiquitin ligase activity in its C-terminus (Abramovitch et al., 2003). HopF2 targets RIN4 and blocks ETI (Wilton et al., 2010). AvrAC can also block ETI (Feng et al., 2012). Furthermore, Jamir et al., showed that effectors could inhibit cell death triggered in yeast, suggesting those effectors target programmed cell death pathways conserved in eukaryotes (Jamir et al., 2004). Subcellular localizations and enzymatic functions of P. syringae type III effectors
As described above, T3Es can inhibit PTI and ETI and thus contribute to virulence. Given the diversity of targets noted above for particular T3Es, it is not surprising to learn that they function in various subcellular locations. Thus, determining the role of any T3E in bacterial virulence requires multiple approaches and gathering pertinent forms of data, such as the subcellular localization of the effector, the enzymatic function, and a potential host target.
eukaryotic-specific posttranslational modification of fatty acid acylation at the N-terminus (Nimchuk et al., 2000). At the plasma membrane that AvrRpm1 and AvrB trigger the phosphorylation of RIN4, leading to the elicitation of RPM1-mediated ETI. Myristoylation sites of AvrRpm1 and AvrB are required to trigger ETI (Nimchuk et al., 2000). Since then multiple type III effectors such have been shown to localize to the plasma membrane via acylation and their functions are dependent on their proper targeting (Dowen et al., 2009; Lewis et al., 2008; Robert-Seilaniantz et al., 2006; Shan et al., 2000).
The chloroplasts, mitochondria, and nucleus are all organelles targeted by T3Es. HopI1 is targeted to the chloroplasts via a noncanonical targeting sequence (Jelenska et al., 2007). HopI1 alters thylakoid structure, suppresses SA accumulation and SA-related
defenses and interacts with Hsp70 (Jelenska et al., 2010; Jelenska et al., 2007). HopN1 also is targeted to the chloroplast, where it degrades a photosystem II protein, PsbQ (Rodriguez-Herva et al., 2012). HopG1 is targeted to the mitochondria and although its specific protein target it currently unknown, HopG1 causes a reduction in PAMP-induced callose deposition and ROS burst (Block et al., 2010). Although there are no published examples of P. syringae T3Es being targeted directly to the nucleus, the AvrBs3 family of T3Es from Xanthomonas spp. are targeted to the nucleus, where they act as transcriptional activators to directly regulate host gene expression (Boch and Bonas, 2010).
2003). HopU1 is an ADP ribosyltransferase (Fu et al., 2007). AvrAC is a uridylyl transferase from Xanthomonas campestris pv. campestris (Feng et al., 2012). HopX1 is predicted to be a transglutaminase (Nimchuk et al., 2007). While the transglutaminase function has not been shown, the putative catalytic residues are required for HopX1-triggered ETI in beans (Nimchuk et al., 2007).
Assigning enzymatic functions to T3Es is challenging. T3Es have relatively low conservation at the primary amino acid sequence level to proteins of known biochemical function. This suggests that convergent evolution into conserved folds to modify host
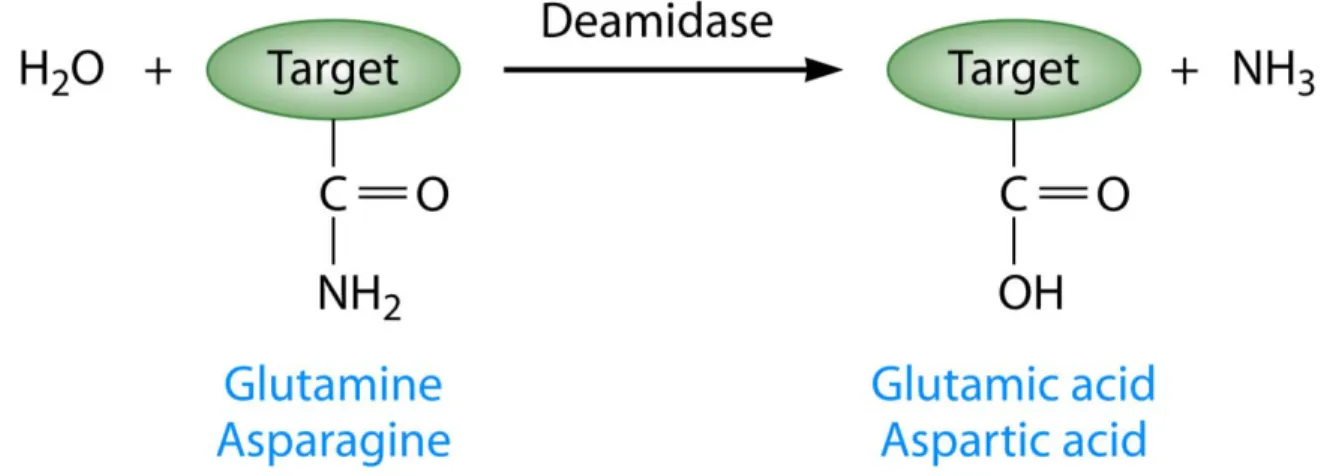
signaling pathways. Solving crystal structures of T3Es is way to help determine a conserved structure associated with a function and/or specific residues required for function (Boutemy et al., 2011; Jeong et al., 2011; Lee et al., 2004; Singer et al., 2004). In lieu of an actual crystal structure, generating tertiary structure predictions can also provide some information about the function of a T3E. AvrRpm1 is predicted to have a fold similar to the catalytic domain of poly-ADP-ribosyl polymerase and the central domains of HopQ1 is predicted to have homology to nucleoside hydrolase enzymes (Cherkis et al., 2012; Li et al., 2013). Additionally, SseI from Salmonella is predicted to have deamidase function based on homology to the bacterial deamidase PMT. However, despite many efforts, SseI has not been shown to have deamidation activity (Bhaskaran and Stebbins, 2012). Deamidation is the irreversible conversion of glutamine and asparagines to glutamic acid and aspartic acid, respectively. Since deamidation is emerging as a posttranslational modification utilized by multiple bacterial toxins to modify host signaling pathways, we will explore bacterial deamidases further in Chapter 2 (Washington et al., 2013).
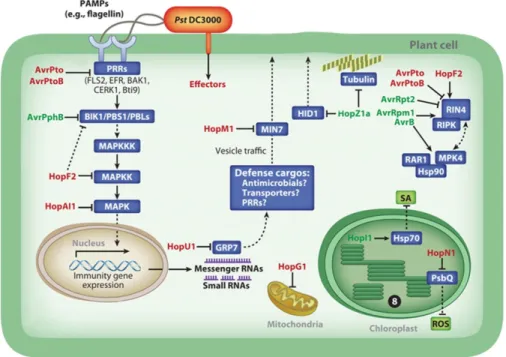
Figure 1.1. P. syringae type III effectors target multiple host cellular components.
Adapted from Xin and He, 2013. Bacterial PAMPs, such as flagellin, trigger PTI. P. syringae injects type III effector proteins into the plant cell, via the type III secretion system, to block components of PTI and ETI. AvrPto and AvrPtoB inhibit the activity of PRRs at the plasma membrane through inhibition of PRR kinase activity or degradation, respectively. AvrPphB cleaves BIK1, PBS1, and PBS1-like proteins (PBLs). HopF2 ADP-ribosylates MKK5 and inhibits its activity. HopF2 may also inhibit PTI at the site of the plasma membrane by blocking BIK1 phosphorylation. HopAI1 also targets the MAPK signaling cascade by using phosphothreonine lyase activity to inhibit the function of MPK3 and MPK6. HopU1 is a mono-ADP-ribosyltransferase that targets glycine-rich RNA-binding proteins (GR-RBPs). HopU1 prevents GRP7 from interacting with FLS2 and EFR transcripts, leading to a reduction in FLS2 transcript and protein accumulation. HopM1 targets vesicle trafficking by inhibiting the function of the ARF-GEF protein MIN7. HopZ1a has been reported to have multiple roles. HopZ1a degrades HID1, a protein involved in biosynthesis of antimicrobial phytoalexins in soybean. HopZ1a also acetylates tubulin. Recently, HopZ1a has been reported to target a pseudokinase, ZED1, which triggers ZAR1-mediated recognition in Arabidopsis. Multiple effectors target RIN4. AvrRpt2 cleaves RIN4, triggering RPS2-mediated ETI. HopF2 blocks ETI triggered by AvrRpt2 by interacting with RIN4.
Phosphorylation of RIN4 in the presence of AvrRpm1 and AvrB is mediated by RIPK. AvrPto and AvrPtoB can degrade RIN4. Finally, AvrB targets RIN4 in complex with
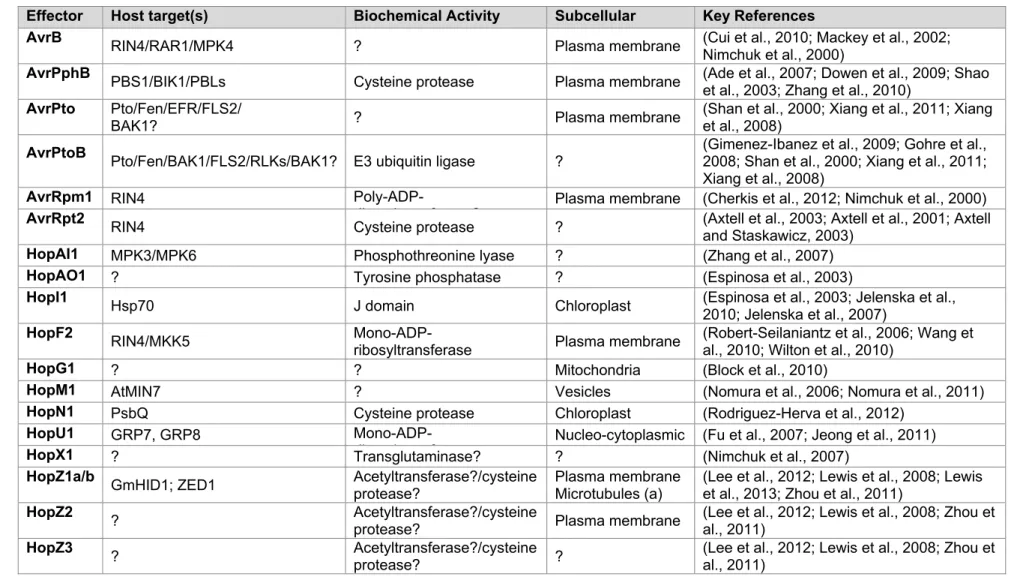
Table 1.1. P. syringae type III effectors: defense suppression, localization, activities and targets
Effector Host target(s) Biochemical Activity Subcellular Key References
AvrB RIN4/RAR1/MPK4 ? Plasma membrane (Cui et al., 2010; Mackey et al., 2002;
Nimchuk et al., 2000)
AvrPphB PBS1/BIK1/PBLs Cysteine protease Plasma membrane (Ade et al., 2007; Dowen et al., 2009; Shao
et al., 2003; Zhang et al., 2010)
AvrPto Pto/Fen/EFR/FLS2/
BAK1? ? Plasma membrane (Shan et al., 2000; Xiang et al., 2011; Xianget al., 2008)
AvrPtoB Pto/Fen/BAK1/FLS2/RLKs/BAK1? E3 ubiquitin ligase ? (Gimenez-Ibanez et al., 2009; Gohre et al., 2008; Shan et al., 2000; Xiang et al., 2011; Xiang et al., 2008)
AvrRpm1 RIN4
Poly-ADP-ib lt f ? Plasma membrane (Cherkis et al., 2012; Nimchuk et al., 2000)
AvrRpt2 RIN4 Cysteine protease ? (Axtell et al., 2003; Axtell et al., 2001; Axtell
and Staskawicz, 2003)
HopAI1 MPK3/MPK6 Phosphothreonine lyase ? (Zhang et al., 2007)
HopAO1 ? Tyrosine phosphatase ? (Espinosa et al., 2003)
HopI1 Hsp70 J domain Chloroplast (Espinosa et al., 2003; Jelenska et al.,
2010; Jelenska et al., 2007)
HopF2 RIN4/MKK5
Mono-ADP-ribosyltransferase Plasma membrane (Robert-Seilaniantz et al., 2006; Wang etal., 2010; Wilton et al., 2010)
HopG1 ? ? Mitochondria (Block et al., 2010)
HopM1 AtMIN7 ? Vesicles (Nomura et al., 2006; Nomura et al., 2011)
HopN1 PsbQ Cysteine protease Chloroplast (Rodriguez-Herva et al., 2012)
HopU1 GRP7, GRP8
Mono-ADP-ib lt f Nucleo-cytoplasmic (Fu et al., 2007; Jeong et al., 2011)
HopX1 ? Transglutaminase? ? (Nimchuk et al., 2007)
HopZ1a/b GmHID1; ZED1 Acetyltransferase?/cysteine
protease?
Plasma membrane Microtubules (a)
(Lee et al., 2012; Lewis et al., 2008; Lewis et al., 2013; Zhou et al., 2011)
HopZ2 ? Acetyltransferase?/cysteine
protease? Plasma membrane (Lee et al., 2012; Lewis et al., 2008; Zhou etal., 2011)
HopZ3 ? Acetyltransferase?/cysteine
protease? ? (Lee et al., 2012; Lewis et al., 2008; Zhou et al., 2011)
REFERENCES
Abramovitch, R.B., Kim, Y.J., Chen, S., Dickman, M.B., and Martin, G.B. (2003). Pseudomonas type III effector AvrPtoB induces plant disease susceptibility by inhibition of host programmed cell death. The EMBO journal 22, 60-69.
Ade, J., DeYoung, B.J., Golstein, C., and Innes, R.W. (2007). Indirect activation of a plant nucleotide binding site-leucine-rich repeat protein by a bacterial protease.
Proceedings of the National Academy of Sciences of the United States of America 104, 2531-2536.
Asai, T., Tena, G., Plotnikova, J., Willmann, M.R., Chiu, W.L., Gomez-Gomez, L., Boller, T., Ausubel, F.M., and Sheen, J. (2002). MAP kinase signalling cascade in Arabidopsis innate immunity. Nature 415, 977-983.
Axtell, M.J., Chisholm, S.T., Dahlbeck, D., and Staskawicz, B.J. (2003). Genetic and
molecular evidence that the Pseudomonas syringae type III effector protein AvrRpt2 is a cysteine protease. Molecular microbiology 49, 1537-1546.
Axtell, M.J., McNellis, T.W., Mudgett, M.B., Hsu, C.S., and Staskawicz, B.J. (2001). Mutational analysis of the Arabidopsis RPS2 disease resistance gene and the corresponding pseudomonas syringae avrRpt2 avirulence gene. Molecular plant-microbe interactions : MPMI 14, 181-188.
Axtell, M.J., and Staskawicz, B.J. (2003). Initiation of RPS2-specified disease resistance in Arabidopsis is coupled to the AvrRpt2-directed elimination of RIN4. Cell 112, 369-377.
Baltrus, D.A., Nishimura, M.T., Romanchuk, A., Chang, J.H., Mukhtar, M.S., Cherkis, K., Roach, J., Grant, S.R., Jones, C.D., and Dangl, J.L. (2011). Dynamic evolution of pathogenicity revealed by sequencing and comparative genomics of 19
Pseudomonas syringae isolates. PLoS pathogens 7, e1002132.
Bent, A.F., Innes, R.W., Ecker, J.R., and Staskawicz, B.J. (1992). Disease development in ethylene-insensitive Arabidopsis thaliana infected with virulent and avirulent
Pseudomonas and Xanthomonas pathogens. Molecular plant-microbe interactions : MPMI 5, 372-378.
Bhaskaran, S.S., and Stebbins, C.E. (2012). Structure of the catalytic domain of the Salmonella virulence factor SseI. Acta crystallographica Section D, Biological crystallography 68, 1613-1621.
Block, A., Guo, M., Li, G., Elowsky, C., Clemente, T.E., and Alfano, J.R. (2010). The Pseudomonas syringae type III effector HopG1 targets mitochondria, alters plant development and suppresses plant innate immunity. Cellular microbiology 12, 318-330.
Boller, T., and Felix, G. (2009). A renaissance of elicitors: perception of microbe-associated molecular patterns and danger signals by pattern-recognition receptors. Annual review of plant biology 60, 379-406.
Bonardi, V., Cherkis, K., Nishimura, M.T., and Dangl, J.L. (2012). A new eye on NLR
proteins: focused on clarity or diffused by complexity? Current opinion in immunology 24, 41-50.
Bonardi, V., and Dangl, J.L. (2012). How complex are intracellular immune receptor signaling complexes? Frontiers in plant science 3, 237.
Bonardi, V., Tang, S., Stallmann, A., Roberts, M., Cherkis, K., and Dangl, J.L. (2011). Expanded functions for a family of plant intracellular immune receptors beyond specific recognition of pathogen effectors. Proceedings of the National Academy of Sciences of the United States of America 108, 16463-16468.
Boutemy, L.S., King, S.R., Win, J., Hughes, R.K., Clarke, T.A., Blumenschein, T.M., Kamoun, S., and Banfield, M.J. (2011). Structures of Phytophthora RXLR effector proteins: a conserved but adaptable fold underpins functional diversity. The Journal of biological chemistry 286, 35834-35842.
Boutrot, F., Segonzac, C., Chang, K.N., Qiao, H., Ecker, J.R., Zipfel, C., and Rathjen, J.P. (2010). Direct transcriptional control of the Arabidopsis immune receptor FLS2 by the ethylene-dependent transcription factors EIN3 and EIL1. Proceedings of the National Academy of Sciences of the United States of America 107, 14502-14507.
Buell, C.R., Joardar, V., Lindeberg, M., Selengut, J., Paulsen, I.T., Gwinn, M.L., Dodson, R.J., Deboy, R.T., Durkin, A.S., Kolonay, J.F., et al. (2003). The complete genome sequence of the Arabidopsis and tomato pathogen Pseudomonas syringae pv. tomato DC3000. Proceedings of the National Academy of Sciences of the United States of America 100, 10181-10186.
Chang, J.H., Urbach, J.M., Law, T.F., Arnold, L.W., Hu, A., Gombar, S., Grant, S.R.,
Ausubel, F.M., and Dangl, J.L. (2005). A high-throughput, near-saturating screen for type III effector genes from Pseudomonas syringae. Proceedings of the National Academy of Sciences of the United States of America 102, 2549-2554.
Chatterjee, S., Chaudhury, S., McShan, A.C., Kaur, K., and De Guzman, R.N. (2013). Structure and biophysics of type III secretion in bacteria. Biochemistry 52, 2508-2517.
Chen, H., Xue, L., Chintamanani, S., Germain, H., Lin, H., Cui, H., Cai, R., Zuo, J., Tang, X., Li, X., et al. (2009). ETHYLENE INSENSITIVE3 and ETHYLENE INSENSITIVE3-LIKE1 repress SALICYLIC ACID INDUCTION DEFICIENT2 expression to negatively regulate plant innate immunity in Arabidopsis. The Plant cell 21, 2527-2540.
Cherkis, K.A., Temple, B.R., Chung, E.H., Sondek, J., and Dangl, J.L. (2012). AvrRpm1 missense mutations weakly activate RPS2-mediated immune response in Arabidopsis thaliana. PloS one 7, e42633.
Chinchilla, D., Zipfel, C., Robatzek, S., Kemmerling, B., Nurnberger, T., Jones, J.D., Felix, G., and Boller, T. (2007). A flagellin-induced complex of the receptor FLS2 and BAK1 initiates plant defence. Nature 448, 497-500.
Cohn, J.R., and Martin, G.B. (2005). Pseudomonas syringae pv. tomato type III effectors AvrPto and AvrPtoB promote ethylene-dependent cell death in tomato. The Plant journal : for cell and molecular biology 44, 139-154.
Cui, H., Wang, Y., Xue, L., Chu, J., Yan, C., Fu, J., Chen, M., Innes, R.W., and Zhou, J.M. (2010). Pseudomonas syringae effector protein AvrB perturbs Arabidopsis hormone signaling by activating MAP kinase 4. Cell host & microbe 7, 164-175.
Cui, J., Bahrami, A.K., Pringle, E.G., Hernandez-Guzman, G., Bender, C.L., Pierce, N.E., and Ausubel, F.M. (2005). Pseudomonas syringae manipulates systemic plant defenses against pathogens and herbivores. Proceedings of the National Academy of Sciences of the United States of America 102, 1791-1796.
Dangl, J.L., and Jones, J.D. (2001). Plant pathogens and integrated defence responses to infection. Nature 411, 826-833.
Dardick, C., and Ronald, P. (2006). Plant and animal pathogen recognition receptors signal through non-RD kinases. PLoS pathogens 2, e2.
de Torres-Zabala, M., Truman, W., Bennett, M.H., Lafforgue, G., Mansfield, J.W., Rodriguez Egea, P., Bogre, L., and Grant, M. (2007). Pseudomonas syringae pv. tomato hijacks the Arabidopsis abscisic acid signalling pathway to cause disease. The EMBO journal 26, 1434-1443.
de Torres, M., Mansfield, J.W., Grabov, N., Brown, I.R., Ammouneh, H., Tsiamis, G., Forsyth, A., Robatzek, S., Grant, M., and Boch, J. (2006). Pseudomonas syringae effector AvrPtoB suppresses basal defence in Arabidopsis. The Plant journal : for cell and molecular biology 47, 368-382.
Deslandes, L., and Rivas, S. (2012). Catch me if you can: bacterial effectors and plant targets. Trends in plant science 17, 644-655.
Dowen, R.H., Engel, J.L., Shao, F., Ecker, J.R., and Dixon, J.E. (2009). A family of bacterial cysteine protease type III effectors utilizes acylation-dependent and -independent strategies to localize to plasma membranes. The Journal of biological chemistry 284, 15867-15879.
Ecker, J.R., and Davis, R.W. (1987). Plant defense genes are regulated by ethylene. Proceedings of the National Academy of Sciences of the United States of America 84, 5202-5206.
Espinosa, A., Guo, M., Tam, V.C., Fu, Z.Q., and Alfano, J.R. (2003). The Pseudomonas syringae type III-secreted protein HopPtoD2 possesses protein tyrosine phosphatase activity and suppresses programmed cell death in plants. Molecular microbiology 49, 377-387.
Fan, J., Crooks, C., Creissen, G., Hill, L., Fairhurst, S., Doerner, P., and Lamb, C. (2011). Pseudomonas sax genes overcome aliphatic isothiocyanate-mediated non-host resistance in Arabidopsis. Science 331, 1185-1188.
Feil, H., Feil, W.S., Chain, P., Larimer, F., DiBartolo, G., Copeland, A., Lykidis, A., Trong, S., Nolan, M., Goltsman, E., et al. (2005). Comparison of the complete genome
sequences of Pseudomonas syringae pv. syringae B728a and pv. tomato DC3000. Proceedings of the National Academy of Sciences of the United States of America 102, 11064-11069.
Felix, G., Duran, J.D., Volko, S., and Boller, T. (1999). Plants have a sensitive perception system for the most conserved domain of bacterial flagellin. The Plant journal : for cell and molecular biology 18, 265-276.
Feng, F., Yang, F., Rong, W., Wu, X., Zhang, J., Chen, S., He, C., and Zhou, J.M. (2012). A Xanthomonas uridine 5'-monophosphate transferase inhibits plant immune kinases. Nature 485, 114-118.
Fu, Z.Q., Guo, M., Jeong, B.R., Tian, F., Elthon, T.E., Cerny, R.L., Staiger, D., and Alfano, J.R. (2007). A type III effector ADP-ribosylates RNA-binding proteins and quells plant immunity. Nature 447, 284-288.
Gimenez-Ibanez, S., Hann, D.R., Ntoukakis, V., Petutschnig, E., Lipka, V., and Rathjen, J.P. (2009). AvrPtoB targets the LysM receptor kinase CERK1 to promote bacterial virulence on plants. Current biology : CB 19, 423-429.
Gohre, V., Spallek, T., Haweker, H., Mersmann, S., Mentzel, T., Boller, T., de Torres, M., Mansfield, J.W., and Robatzek, S. (2008). Plant pattern-recognition receptor FLS2 is directed for degradation by the bacterial ubiquitin ligase AvrPtoB. Current biology : CB 18, 1824-1832.
Gomez-Gomez, L., and Boller, T. (2000). FLS2: an LRR receptor-like kinase involved in the perception of the bacterial elicitor flagellin in Arabidopsis. Molecular cell 5, 1003-1011.
Grant, S.R., Fisher, E.J., Chang, J.H., Mole, B.M., and Dangl, J.L. (2006). Subterfuge and manipulation: type III effector proteins of phytopathogenic bacteria. Annual review of microbiology 60, 425-449.
Hann, D.R., and Rathjen, J.P. (2007). Early events in the pathogenicity of Pseudomonas syringae on Nicotiana benthamiana. The Plant journal : for cell and molecular biology 49, 607-618.
central regulator of innate immunity in plants. Proceedings of the National Academy of Sciences of the United States of America 104, 12217-12222.
Huynh, T.V., Dahlbeck, D., and Staskawicz, B.J. (1989). Bacterial blight of soybean: regulation of a pathogen gene determining host cultivar specificity. Science 245, 1374-1377.
Jackson, R.W., Athanassopoulos, E., Tsiamis, G., Mansfield, J.W., Sesma, A., Arnold, D.L., Gibbon, M.J., Murillo, J., Taylor, J.D., and Vivian, A. (1999). Identification of a pathogenicity island, which contains genes for virulence and avirulence, on a large native plasmid in the bean pathogen Pseudomonas syringae pathovar phaseolicola. Proceedings of the National Academy of Sciences of the United States of America 96, 10875-10880.
Jamir, Y., Guo, M., Oh, H.S., Petnicki-Ocwieja, T., Chen, S., Tang, X., Dickman, M.B., Collmer, A., and Alfano, J.R. (2004). Identification of Pseudomonas syringae type III effectors that can suppress programmed cell death in plants and yeast. The Plant journal : for cell and molecular biology 37, 554-565.
Jelenska, J., van Hal, J.A., and Greenberg, J.T. (2010). Pseudomonas syringae hijacks plant stress chaperone machinery for virulence. Proceedings of the National Academy of Sciences of the United States of America 107, 13177-13182. Jelenska, J., Yao, N., Vinatzer, B.A., Wright, C.M., Brodsky, J.L., and Greenberg, J.T.
(2007). A J domain virulence effector of Pseudomonas syringae remodels host chloroplasts and suppresses defenses. Current biology : CB 17, 499-508.
Jeong, B.R., Lin, Y., Joe, A., Guo, M., Korneli, C., Yang, H., Wang, P., Yu, M., Cerny, R.L., Staiger, D., et al. (2011). Structure function analysis of an ADP-ribosyltransferase type III effector and its RNA-binding target in plant immunity. The Journal of biological chemistry 286, 43272-43281.
Jia, Y., McAdams, S.A., Bryan, G.T., Hershey, H.P., and Valent, B. (2000). Direct interaction of resistance gene and avirulence gene products confers rice blast resistance. The EMBO journal 19, 4004-4014.
Joardar, V., Lindeberg, M., Jackson, R.W., Selengut, J., Dodson, R., Brinkac, L.M.,
Daugherty, S.C., Deboy, R., Durkin, A.S., Giglio, M.G., et al. (2005). Whole-genome sequence analysis of Pseudomonas syringae pv. phaseolicola 1448A reveals divergence among pathovars in genes involved in virulence and transposition. Journal of bacteriology 187, 6488-6498.
Jones, J.D., and Dangl, J.L. (2006). The plant immune system. Nature 444, 323-329. Kaku, H., Nishizawa, Y., Ishii-Minami, N., Akimoto-Tomiyama, C., Dohmae, N., Takio, K.,
Minami, E., and Shibuya, N. (2006). Plant cells recognize chitin fragments for
defense signaling through a plasma membrane receptor. Proceedings of the National Academy of Sciences of the United States of America 103, 11086-11091.
Kim, H.S., Desveaux, D., Singer, A.U., Patel, P., Sondek, J., and Dangl, J.L. (2005). The Pseudomonas syringae effector AvrRpt2 cleaves its C-terminally acylated target, RIN4, from Arabidopsis membranes to block RPM1 activation. Proceedings of the National Academy of Sciences of the United States of America 102, 6496-6501. Kim, J.G., Stork, W., and Mudgett, M.B. (2013). Xanthomonas type III effector XopD
desumoylates tomato transcription factor SlERF4 to suppress ethylene responses and promote pathogen growth. Cell host & microbe 13, 143-154.
Kunze, G., Zipfel, C., Robatzek, S., Niehaus, K., Boller, T., and Felix, G. (2004). The N terminus of bacterial elongation factor Tu elicits innate immunity in Arabidopsis plants. The Plant cell 16, 3496-3507.
Lacombe, S., Rougon-Cardoso, A., Sherwood, E., Peeters, N., Dahlbeck, D., van Esse, H.P., Smoker, M., Rallapalli, G., Thomma, B.P., Staskawicz, B., et al. (2010). Interfamily transfer of a plant pattern-recognition receptor confers broad-spectrum bacterial resistance. Nature biotechnology 28, 365-369.
Laurie-Berry, N., Joardar, V., Street, I.H., and Kunkel, B.N. (2006). The Arabidopsis thaliana JASMONATE INSENSITIVE 1 gene is required for suppression of salicylic acid-dependent defenses during infection by Pseudomonas syringae. Molecular plant-microbe interactions : MPMI 19, 789-800.
Lawton, K.A., Potter, S.L., Uknes, S., and Ryals, J. (1994). Acquired Resistance Signal Transduction in Arabidopsis Is Ethylene Independent. The Plant cell 6, 581-588. Lee, A.H., Hurley, B., Felsensteiner, C., Yea, C., Ckurshumova, W., Bartetzko, V., Wang,
P.W., Quach, V., Lewis, J.D., Liu, Y.C., et al. (2012). A bacterial acetyltransferase destroys plant microtubule networks and blocks secretion. PLoS pathogens 8, e1002523.
Lee, C.C., Wood, M.D., Ng, K., Andersen, C.B., Liu, Y., Luginbuhl, P., Spraggon, G., and Katagiri, F. (2004). Crystal structure of the type III effector AvrB from Pseudomonas syringae. Structure 12, 487-494.
Leipe, D.D., Koonin, E.V., and Aravind, L. (2004). STAND, a class of P-loop NTPases including animal and plant regulators of programmed cell death: multiple, complex domain architectures, unusual phyletic patterns, and evolution by horizontal gene transfer. Journal of molecular biology 343, 1-28.
Lewis, J.D., Abada, W., Ma, W., Guttman, D.S., and Desveaux, D. (2008). The HopZ family of Pseudomonas syringae type III effectors require myristoylation for virulence and avirulence functions in Arabidopsis thaliana. Journal of bacteriology 190, 2880-2891. Lewis, J.D., Lee, A.H., Hassan, J.A., Wan, J., Hurley, B., Jhingree, J.R., Wang, P.W., Lo, T.,
Lewis, J.D., Wu, R., Guttman, D.S., and Desveaux, D. (2010). Allele-specific virulence attenuation of the Pseudomonas syringae HopZ1a type III effector via the Arabidopsis ZAR1 resistance protein. PLoS genetics 6, e1000894.
Li, W., Chiang, Y.H., and Coaker, G. (2013). The HopQ1 effector's nucleoside hydrolase-like domain is required for bacterial virulence in arabidopsis and tomato, but not host recognition in tobacco. PloS one 8, e59684.
Li, X., Lin, H., Zhang, W., Zou, Y., Zhang, J., Tang, X., and Zhou, J.M. (2005). Flagellin induces innate immunity in nonhost interactions that is suppressed by Pseudomonas syringae effectors. Proceedings of the National Academy of Sciences of the United States of America 102, 12990-12995.
Lindgren, P.B., Peet, R.C., and Panopoulos, N.J. (1986). Gene cluster of Pseudomonas syringae pv. "phaseolicola" controls pathogenicity of bean plants and hypersensitivity of nonhost plants. Journal of bacteriology 168, 512-522.
Liu, T., Liu, Z., Song, C., Hu, Y., Han, Z., She, J., Fan, F., Wang, J., Jin, C., Chang, J., et al. (2012). Chitin-induced dimerization activates a plant immune receptor. Science 336, 1160-1164.
Liu, Y., and Zhang, S. (2004). Phosphorylation of 1-aminocyclopropane-1-carboxylic acid synthase by MPK6, a stress-responsive mitogen-activated protein kinase, induces ethylene biosynthesis in Arabidopsis. The Plant cell 16, 3386-3399.
Lopez-Solanilla, E., Bronstein, P.A., Schneider, A.R., and Collmer, A. (2004). HopPtoN is a Pseudomonas syringae Hrp (type III secretion system) cysteine protease effector that suppresses pathogen-induced necrosis associated with both compatible and incompatible plant interactions. Molecular microbiology 54, 353-365.
Ma, W., and Berkowitz, G.A. (2007). The grateful dead: calcium and cell death in plant innate immunity. Cellular microbiology 9, 2571-2585.
Mackey, D., Belkhadir, Y., Alonso, J.M., Ecker, J.R., and Dangl, J.L. (2003). Arabidopsis RIN4 is a target of the type III virulence effector AvrRpt2 and modulates RPS2-mediated resistance. Cell 112, 379-389.
Mackey, D., Holt, B.F., 3rd, Wiig, A., and Dangl, J.L. (2002). RIN4 interacts with Pseudomonas syringae type III effector molecules and is required for RPM1-mediated resistance in Arabidopsis. Cell 108, 743-754.
Melotto, M., Underwood, W., Koczan, J., Nomura, K., and He, S.Y. (2006). Plant stomata function in innate immunity against bacterial invasion. Cell 126, 969-980.
Mersmann, S., Bourdais, G., Rietz, S., and Robatzek, S. (2010). Ethylene signaling regulates accumulation of the FLS2 receptor and is required for the oxidative burst contributing to plant immunity. Plant physiology 154, 391-400.
Miya, A., Albert, P., Shinya, T., Desaki, Y., Ichimura, K., Shirasu, K., Narusaka, Y.,
Kawakami, N., Kaku, H., and Shibuya, N. (2007). CERK1, a LysM receptor kinase, is essential for chitin elicitor signaling in Arabidopsis. Proceedings of the National Academy of Sciences of the United States of America 104, 19613-19618.
Monaghan, J., and Zipfel, C. (2012). Plant pattern recognition receptor complexes at the plasma membrane. Current opinion in plant biology 15, 349-357.
Morris, C.E., Kinkel, L.L., Xiao, K., Prior, P., and Sands, D.C. (2007). Surprising niche for the plant pathogen Pseudomonas syringae. Infection, genetics and evolution : journal of molecular epidemiology and evolutionary genetics in infectious diseases 7, 84-92. Morris, C.E., Sands, D.C., Vanneste, J.L., Montarry, J., Oakley, B., Guilbaud, C., and Glaux,
C. (2010). Inferring the evolutionary history of the plant pathogen Pseudomonas syringae from its biogeography in headwaters of rivers in North America, Europe, and New Zealand. mBio 1.
Morris, C.E., Sands, D.C., Vinatzer, B.A., Glaux, C., Guilbaud, C., Buffiere, A., Yan, S., Dominguez, H., and Thompson, B.M. (2008). The life history of the plant pathogen Pseudomonas syringae is linked to the water cycle. The ISME journal 2, 321-334. Mukhtar, M.S., Carvunis, A.R., Dreze, M., Epple, P., Steinbrenner, J., Moore, J., Tasan, M.,
Galli, M., Hao, T., Nishimura, M.T., et al. (2011). Independently evolved virulence effectors converge onto hubs in a plant immune system network. Science 333, 596-601.
Nicaise, V., Joe, A., Jeong, B.R., Korneli, C., Boutrot, F., Westedt, I., Staiger, D., Alfano, J.R., and Zipfel, C. (2013). Pseudomonas HopU1 modulates plant immune receptor levels by blocking the interaction of their mRNAs with GRP7. The EMBO journal 32, 701-712.
Nimchuk, Z., Marois, E., Kjemtrup, S., Leister, R.T., Katagiri, F., and Dangl, J.L. (2000). Eukaryotic fatty acylation drives plasma membrane targeting and enhances function of several type III effector proteins from Pseudomonas syringae. Cell 101, 353-363. Nimchuk, Z.L., Fisher, E.J., Desveaux, D., Chang, J.H., and Dangl, J.L. (2007). The HopX
(AvrPphE) family of Pseudomonas syringae type III effectors require a catalytic triad and a novel N-terminal domain for function. Molecular plant-microbe interactions : MPMI 20, 346-357.
Nomura, K., Debroy, S., Lee, Y.H., Pumplin, N., Jones, J., and He, S.Y. (2006). A bacterial virulence protein suppresses host innate immunity to cause plant disease. Science 313, 220-223.
Nomura, K., Mecey, C., Lee, Y.N., Imboden, L.A., Chang, J.H., and He, S.Y. (2011).
Effector-triggered immunity blocks pathogen degradation of an immunity-associated vesicle traffic regulator in Arabidopsis. Proceedings of the National Academy of Sciences of the United States of America 108, 10774-10779.