SHORT COMMUNICATION
A NEW MICROCOMPUTER-BASED METHOD FOR
MEASURING WALKING PHONOTAXIS IN FIELD
CRICKETS (GRYLLIDAE)
BY JOHN A. DOHERTY* AND ANTHONY PIRES
Section of Neurobiology and Behavior, Seeley G. Mudd Hall, Cornell University, Ithaca, NY 14853, USA
Accepted 13 March 1987
Calling songs of male crickets attract sexually mature, conspecific females for mating (for recent reviews see Eisner & Popov, 1978; Huber & Thorson, 1985; Doherty & Hoy, 1985). This communication system has been the subject of many behavioural studies on the relevant properties of the male's calling song for recognition by females (e.g. Walker, 1957; Popov & Shuvalov, 1977; Pollack & Hoy, 1979, 1981; Thorson, Weber & Huber, 1982; Stout, DeHaan & McGhee, 1983; Doherty, 19856; Doolan & Pollack, 1985). The behavioural investigations have required some means of measuring the female's phonotaxis (her locomotion or turning towards a sound source). These include 'closed-loop' methods in which the animal moves freely in acoustic space and 'open-loop' methods in which the animal is tethered and is not allowed to experience changes in sound intensities as it runs in the direction of the sound source. Closed-loop methods include free walking in arenas (e.g. Murphey & Zaretsky, 1972; Paul, 1976; Hoy, Hahn & Paul, 1977; Pollack & Hoy, 1979; Stout et al. 1983); open-loop methods include tethered flight (Moiseff, Pollack & Hoy, 1978; Pollack & Hoy, 1979, 1981; Pollack, Huber & Weber, 1984), tethered walking on Y-maze globes (Hoy & Paul, 1973) and free walking on a spherical locomotion compensator (Wendler, Dambach, Schmitz & Scharstein, 1980; Thorson & Huber, 1981; Thorson et al. 1982; Pollack et al. 1984; Doherty,
I985a,b,c; Schmitz, 1985).
Here we describe a new open-loop method for quantifying cricket phonotaxis. This method uses inexpensive microcomputer technology, is completely automated and therefore rapid and objective, and could be adapted for studying locomotory movements of other animals. The data obtained are in such a form that they are easily compared to data generated by the spherical locomotion compensator (Kramer treadmill, see Kramer, 1976; Wendler^ al. 1980; Weber et al. 1981), which has been used for quantifying phonotaxis in the field cricket, Gryllus bimaculatus (Doherty, 1985a, b,c).
The spherical locomotion compensator or Kramer treadmill was first developed by scientists in West Germany (Kramer, 1976; Wendlere* al. 1980; Weber et al. 1981; Thorson et al. 1982). It uses an infrared detection system to monitor movements of an untethered cricket on top of a sphere. This positional information feeds back to servomotors that move the sphere in the opposite direction. These compensatory movements 'fix' the cricket at the top of the sphere as it performs walking phonotaxis. The colloquial name for this locomotion compensator is 'Kugel' (German translation of the word 'sphere'). Because our device for measuring walking phonotaxis utilizes the Apple Macintosh computer and is similar in appearance to the German Kugel, we call our new device the 'MacKugeP. The main mechanical difference between these two devices is that in the MacKugel system the cricket is tethered and its own walking movements provide the power to rotate the sphere.
The MacKugel is a complete stimulus control and data acquisition system for on-line studies of cricket phonotaxis. Data on cricket movements during phonotaxis experiments are passed directly to an Apple Macintosh microcomputer, which can also synthesize acoustic stimuli or send control signals to an external acoustic synthesizer. In designing this system, we took advantage of ROM routines in the Macintosh for quantifying movements of the Apple 'mouse'.
The mouse is a mechanical, optoelectronic device that converts rotational movements of a rubber ball to changes in the coordinates of a cursor displayed on the computer's CRT monitor. The ball bears against the operator's desk surface, and against three rollers inside the ball's housing. One roller is for mechanical support only. The other two, representing orthogonal x- and y-axes, are connected to rotating vanes which interrupt light beams between LED—phototransistor pairs. The phototransistors produce electronic signals which are sent to the computer.
R T U
K
F
**-£ F
^
S
V X
r
1t
t
TTTjll
©
[image:3.451.32.407.42.319.2]@
Fig. 1. Side (A) and top (B) view of MacKugel device. TM, tether mount; F, frame; RT, rotating tether; PR, passive roller; T , , x-axis tachometer (composed of photo-transistor module, extension axle and roller); Ty, y-axis tachometer; FB, frictionless
bearing (composed of a 1 cm Teflon ball riding on a film of air); AS, air supply (air routed through a threaded aluminium cylinder used to adjust the height of the Nerf ball); S, sphere.
To establish the usefulness and validity of our method, we repeated some locomotion compensator experiments run previously on the field cricket, G.
bimacu-latus (Doherty, 1985a,b). In both methods, walking phonotaxis in crickets can be
quantified by measuring time profiles of the cricket's walking velocity and direction. Fig. 2 shows these time profiles for one MacKugel experiment in which a synthetic calling song was presented to a female G. bimaculatus from one of two loudspeakers separated by 180°. This calling song was played from the left speaker for 2min and then from the right speaker for another 2min. A 30-s silent period preceded and followed this playback. The female tracked the stimulus, as shown by her narrow meandering about the direction of the active speaker. Her walking velocity also increased when the stimulus was presented, compared to the preceding silent period.
The direction component of cricket walking phonotaxis is a sensitive indicator of the attractiveness of an acoustic stimulus. To investigate this sensitivity further, we ran phonotaxis experiments in which acoustic stimuli with different pulse periods were presented in a sequential, to-and-fro paradigm. As in earlier experiments of cricket phonotaxis on a locomotion compensator (Doherty, 1985c), pulse periods ranging from 30 to 50 ms yielded the best tracking on the MacKugel (i.e. best orientation to active speaker direction, see Fig. 3).
CP
PD
1313 76%
"a.
.^•T 4
io
n
1
Angula
r d
•2- 0
0'
RS
LS
360'
r
[image:4.451.116.336.182.425.2]1 min
In these same phonotaxis experiments on the MacKugel we found slight effects of pulse period on the walking velocity profiles. Females walked for a greater percentage of the stimulus presentation time when pulse periods were optimal (i.e. 30-50ms)
•c
O
0-6 -i
0 4 0 2
-0 -0
- 0 - 2
40
39
10 20 30 40 50 60 Pulse period (ms)
[image:5.451.122.330.132.288.2]70 80
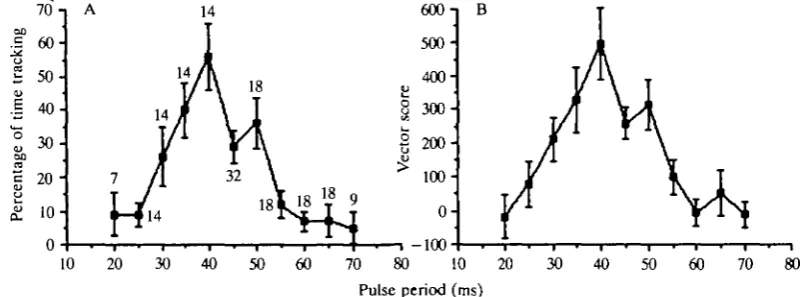
Fig. 3. Effects of synthetic calling songs with different pulse periods on the accuracy of orientation to the speaker. Orientation was quantified as follows: for each 1-s interval of a trial in which the animal was moving, the cosine of the vector angle was calculated. A mean cos (angle) was then obtained for all such intervals within a trial. An average cos (angle) was computed for all trials of each pulse period and plotted as orientation to speaker. If the cricket ran directly towards the loudspeaker (defined as 0°), the cos (angle) was 1; this value was — 1 when it ran directly away from the speaker and 0 when it ran perpendicular to the sound source. The points are means for each pulse period and the vertical lines are standard errors. Data were based on to-and-fro scans of pulse period for 21 female Gryllus bimaculatus. Sample sizes are shown.
70-i 600-1
20 30 40 50 60
i -100
B
70 80 10 20 Pulse period (ms)
30 40 50 60 70
Fig. 4. Comparison of two different methods of quantifying stimulus efficacy in sequential, to-and-fro scans of pulse period (see text and Doherty, 1985a,c). The two methods were (A) manual measurements of the percentage of stimulus presentation time that females tracked the stimulus and (B) microcomputer measurements of vector score. The criteria for manual scoring of stimulus tracking are found in Doherty (1985a,c). The points are mean values and the vertical lines are standard errors. Nine Gryllus
bimaculatus females were tested. Sample sizes are the same in A and B.
[image:5.451.23.426.415.563.2]and walked less in response to stimuli with pulse periods outside this range. Furthermore, mean walking velocity was highest in response to pulse periods of 35 and 40 ms, whether or not pauses in walking were included in the calculation.
Because both walking velocity and direction were affected by acoustic stimuli, we devised a numerical 'vector score' which incorporates both of these components and serves as a quick measure of the phonotactic efficacy of different sound stimuli. This score is defined as:
Vector score = 2[cos(vector angle) X vector length].
In calculating the vector score, the angular direction of the active speaker is always defined as 0° and the summation is over all the sample intervals within a trial. When the animal runs towards the loudspeaker the vector score increases, when it runs away from the loudspeaker the score decreases. By definition, movements perpen-dicular to the speaker axis (cos 90° or cos 270°) and no movements at all (vector length = 0) result in vector scores approaching zero. For example, in Fig. 2, the vector score in response to calling song from the left speaker was lower than that for the right speaker. This difference was due to less accurate tracking, wider meanderings about the speaker direction and more pauses in walking.
The vector score is comparable to other measures of stimulus efficacy in cricket phonotaxis. In earlier studies of G. bimaculatus phonotaxis on a locomotion compensator, the attractiveness or efficacy of an auditory stimulus was quantified by measuring the percentage of the stimulus presentation time that the female clearly 'tracked' the stimulus. Tracking has been defined as meandering within a certain angle 'window' (±60°) about the angular direction of the active speaker (see Thorson
et al. 1982; Doherty, 1985a,b,c). These same criteria were used in our study for
measuring the percentage of time spent tracking in a group of nine G. bimaculatus females. In to-and-fro sequential experiments, tracking scores and vector scores in response to calling songs with different pulse periods were comparable (Fig. 4). Both scores showed that songs with pulse periods between 35 and 50 ms were most effective in eliciting positive phonotaxis.
The MacKugel system is also useful for quantifying cricket phonotaxis in two-stimulus (choice) playback experiments. The total vector score was a sensitive, objective measure of the relative attractiveness of alternative acoustic stimuli. When given a choice between alternating chirps with different pulse periods (PP), female
G. bimaculatus clearly preferred the standard chirp stimulus (40 ms PP) to the
alternative chirp stimuli with pulse periods of 20, 30, 50 and 60 ms. The strength of the female's preference for the standard over the alternative pulse period was also reflected in the mean vector score. These results were consistent with those of previous choice experiments on a locomotion compensator (Doherty, 1985c).
used as a rough basis of comparison between individuals and between experimental treatments. Data analysis can be done 'on-line' as the animal is behaving. Other strengths of the MacKugel are its low price, simplicity of design, and efficient implementation of existing microcomputer technology.
This research was supported by grants from NSF and NIH to JAD, AP and Ronald R. Hoy. We thank Alan Cohen and Rick Nicoletti for technical assistance and Wendy Sussdorf for preparing Fig. 1. Ronald Hoy, Peter Brodfuehrer and Jud Crawford read earlier drafts of this manuscript.
REFERENCES
DoHERTY, J. A. (1985a). Temperature coupling and 'trade-off phenomena in the acoustic communication system of the cricket, Gryllus bimaculatus De Geer (Gryllidae). J. exp. Biol. 114, 17-35.
DOHERTY, J. A. (19856). Trade-off phenomena in calling song recognition and phonotaxis in the cricket, Gryllus bimaculatus (Orthoptera, Gryllidae). J . comp. Physiol. 156, 787-801.
DOHERTY, J. A. (1985c). Phonotaxis in the cricket, Gryllus bimaculatus De Geer: Comparisons of choice and no-choice paradigms, jf. comp. Physiol. 157, 279-289.
DOHERTY, J. A. & HOY, R. R. (1985). Communication in insects. I I I . The auditory behavior of crickets: Some views of genetic coupling, song recognition, and predator detection. Q. Rev. Biol. 60, 457-472.
DOOLAN, J. M. & POLLACK, G. S. (1985). Phonotactic specificity of the cricket Teleogryllus
oceanicus: intensity-dependent selectivity for temporal parameters of the stimulus. J. comp. Physiol. 157, 223-234.
ELSNER, N. & POPOV, A. V. (1978). Neuroethology of acoustic communication. Adv. Insect
Physiol. 13,229-335.
HOY, R. R., HAHN, J. & PAUL, R. C. (1977). Hybrid cricket auditory behavior: Evidence for genetic coupling in animal communication. Science 195, 82-84.
HOY, R. R. & PAUL, R. C. (1973). Genetic control of song specificity in crickets. Science 180, 82-83.
HUBER, F. & THORSON, J. (1985). Cricket auditor)' communication. Scient. Am. 253, 60-68. KRAMER, E. (1976). The orientation of walking honeybees in odour fields with small concentration
gradients. Physiol. Entomol. 1, 27-37.
MOISEFF, A., POLLACK, G. S. & HOY, R. R. (1978). Steering responses of flying crickets to sound and ultrasound: Mate attraction and predator avoidance. Proc. natn. Acad. Set. (J.SA. 75, 4052-4056.
MURPHEY, R. K. & ZARETSKY, M. D. (1972). Orientation to calling song by female crickets,
Scapsipedus marginatus (Gryllidae). J. exp. Biol. 56, 335-352,
PAUL, R. C. (1976). Species specificity in the phonotaxis of female ground crickets (Orthoptera: Gryllidae: Nemobiinae). Ann. ent. Soc.Am. 69, 1007-1010.
POLLACK, G. S. & HOY, R. R. (1979). Temporal pattern as a cue for species-specific calling song recognition in crickets. Science 204, 429-432.
POLLACK, G. S. & HOY, R. R. (1981). Phonotaxis to individual rhythmic components of a complex cricket calling song. J. comp. Physiol. 144, 367—373.
POLLACK, G. S., HUBER, F. & WEBER, T . (1984). Frequency and temporal pattern-dependent phonotaxis of crickets {Teleogryllus oceanicus) during tethered flight and compensated walking.
J. comp. Physiol. 154, 13-26.
POPOV, A. V. & SHUVALOV, V. F. (1977). Phonotactic behavior of crickets. J. comp. Phvsiol. 119, 111-126.
STOUT, J. F., DEHAAN, C. H. & MCGHEE, R. W. (1983). Attractiveness of the male Acheta
domesticus calling song to females. I. Dependence on each of the calling song features..7. comp. Physiol. 153, 509-521.
THORSON, J., WEBER, T . & HUBER, F. (1982). Auditory behaviour of the cricket. I I . Simplicity of calling-song recognition in Gryllus, and anomalous phonotaxis at abnormal carrier frequencies.
J. comp. Physiol. 146, 361-378.
WALKER, T . J. (1957). Specificity in the response of female tree crickets (Orthoptera, Gryllidae, Oecanthinae) to calling songs of the males. Ann. ent. Soc. Am. 50, 626—636.
WEBER, T . , THORSON, J. & HUBER, F. (1981). Auditory behavior of the cricket. I. Dynamics of compensated walking and discrimination paradigms on the Kramer treadmill..7. comp. Physiol. 141, 215-232.
WENDLER, G., DAMBACH, M., SCHMITZ, B. & SCHARSTEIN, H. (1980). Analysis of the acoustic