REVIEW
Behavioural and neuronal basis of olfactory imprinting and kin
recognition in larval fish
Gabriele Gerlach1,2,3,*, Kristin Tietje1, Daniela Biechl4, Iori Namekawa5, Gregor Schalm1and Astrid Sulmann1
ABSTRACT
Imprinting is a specific form of long-term memory of a cue acquired during a sensitive phase of development. To ensure that organisms memorize the right cue, the learning process must happen during a specific short time period, mostly soon after hatching, which should end before irrelevant or misleading signals are encountered. A well-known case of olfactory imprinting in the aquatic environment is that of the anadromous Atlantic and Pacific salmon, which prefer the olfactory cues of natal rivers to which they return after migrating several years in the open ocean. Recent research has shown that olfactory imprinting and olfactory guided navigation in the marine realm are far more common than previously assumed. Here, we present evidence for the involvement of olfactory imprinting in the navigation behaviour of coral reef fish, which prefer their home reef odour over that of other reefs. Two main olfactory imprinting processes can be differentiated: (1) imprinting on environmental cues and (2) imprinting on chemical compounds released by kin, which is based on genetic relatedness among conspecifics. While the first process allows for plasticity, so that organisms can imprint on a variety of chemical signals, the latter seems to be restricted to specific genetically determined kin signals. We focus on the second, elucidating the behavioural and neuronal basis of the imprinting process on kin cues using larval zebrafish (Danio rerio) as a model. Our data suggest that the process of imprinting is not confined to the central nervous system but also triggers some changes in the olfactory epithelium.
KEY WORDS: Coral reef fish, Larval dispersal, Orientation, MHC peptides, Olfaction, Crypt cell
Introduction
Olfactory guided navigation in coral reef fish
Like many aquatic organisms, coral reef fish show a dual life stage, where settled adults produce dispersing larvae. At hatching, planktonic larvae drift away from the reef to spend a species-dependent time (larval dispersal duration) in the open ocean – probably to avoid high predation in the reef. These millimetre-sized larvae quickly (within a few days of dispersal) turn into relatively powerful juvenile swimmers, reaching swimming speeds of several centimetres per second (Fisher et al., 2000; Fisher and Wilson,
2004; Stobutzki and Bellwood, 1994, 1997). Until recently, the distribution and settlement of coral reef fish were assumed to be purely driven by currents and stochastic storm events. However, the persistence of marine populations at small isolated oceanic islands requires that a significant number of juveniles return to the natal habitat after their pelagic dispersal phase (Robertson, 2001). Here, and even in less isolated habitats, self-recruitment and natal homing has been assumed to be (far) greater than purely planktonic dispersal would predict from modelling approaches (Armsworth, 2000; Cowen et al., 2000; Staaterman and Paris, 2014; Wolanski et al., 1997). We used population genetic analysis to demonstrate that up to 60% of juvenile cardinalfish,Ostorhinchus doederleini, could be assigned to the adult reef population where they were about to settle (Gerlach et al., 2007a, 2016). Natal homing of other species, e.g. clownfish juveniles (Amphiprion spp.) was also confirmed by mark–recapture studies, otolith tagging and microchemistry studies (Jones et al., 1999; Swearer et al., 1999; Thorrold et al., 2006).
As it is not possible to track larvae in the ocean, dispersal distances are based on catching larvae in plankton tows and by modelling approaches using the pelagic larval duration as a proxy (but see Weersing and Toonen, 2009). Dispersal distances of coral reef fish larvae are assumed to be shorter than 150 km depending on the species (Burgess et al., 2007; Paris and Cowen, 2004). Despite a potentially wide distribution, numbers of homing coral reef fish juveniles are far higher than expected by random movement. Orientation capabilities could help them to find their way back to their natal reefs.
Finding the way back to a natal reef, river or beach after week- or year-long dispersal might require a learning and memory process involving time-dependent specific parameters of this natal place. For orientation-guided homing to natal reefs, coral reef fish larvae must remember sensory parameters that they experienced directly after hatching, which is the only time in which they can obtain reliable‘home cues’, as larvae–with fully developed olfactory and visual sensory systems–start dispersing into the open ocean on the night of hatching.
To explain navigation over long distances, we provided evidence that the juvenile cardinalfish (O. doederleini) can use time-compensated sun compass orientation during the day (Mouritsen et al., 2013) and a magnetic compass at night (Bottesch et al., 2016). For orientation at closer distances, there is evidence that reef fish larvae can also respond to acoustic cues for orientation (Radford et al., 2010, 2011). Additionally, we have documented a pronounced importance of olfaction in the homing of coral reef fish larvae (Atema et al., 2002; Gerlach et al., 2007a; Paris et al., 2013). Coral reef fishes such as O. doederleini, Pomacentrus coelestis and several other species of apogonid and pomacentrid juveniles were shown to prefer the smell of water collected from a reef to that of open ocean water (Atema et al., 2002). Both species were capable of distinguishing between chemical cues from reefs
1Institute of Biology and Environmental Sciences, Carl von Ossietzky University
Oldenburg, 26129 Oldenburg, Germany.2Helmholtz Institute for Functional Marine
Biodiversity Oldenburg (HIFMB), 26129 Oldenburg, Germany.3Centre of
Excellence for Coral Reef Studies and School of Marine and Tropical Biology,
James Cook University, QLD 4811, Australia.4Graduate School of Systemic
Neurosciences & Department Biology II, Ludwig-Maximilians-Universität Munich,
82152 Planegg-Martinsried, Germany.5Friedrich Miescher Institute for Biomedical
Research, 4058 Basel, Switzerland.
*Author for correspondence ([email protected])
G.G., 0000-0001-5246-944X; I.N., 0000-0002-1012-9823
Journal
of
Experimental
within the Capricorn/Bunker reef group and preferred the smell of water from the reef where they were caught over that of other nearby reefs (Gerlach et al., 2007a). We further demonstrated that the preference for home reef odour did not switch even when larvae were exposed to other reef odours for several days (Miller-Sims et al., 2011). Therefore, we concluded that coral reef fish larvae had imprinted on olfactory cues of their home reefs, similar to the well-known olfactory imprinting of salmon on chemical cues of natal rivers (Hasler et al., 1978; Scholz et al., 1973, 1976). However, the robust experimental manipulations that demonstrate an ability of reef fish to imprint upon artificial odours in a similar way to salmonids have yet to be performed.
Olfactory imprinting and olfactory guided navigation in teleosts Imprinting is a specific form of long-term memory enabling organisms to recognize specific cues from the environment and conspecifics. It might enable offspring to find their mothers among many other females or help animals to remember and recognize environmental conditions that they experienced shortly after hatching or birth. To ensure that organisms memorize the right cue, the learning process must happen during a specific short time period, which should end before the organism encounters irrelevant signals from unrelated conspecifics or environments. One well-known example of a visual imprinting process is Konrad Lorenz’s geese experiment (Lorenz, 1935) in which he showed that goslings imprint on the first moving object that they experience after hatching, which they take to be their mother. Many imprinting processes are involving olfactory cues, probably because in social contexts chemical cues can be taken as reliable‘honest’signals and manipulating one’s own body smell is very difficult.
The timing of imprinting has been studied extensively in salmonids, and there is strong evidence that salmon imprint on environmental cues of their natal water during the parr–smolt transformation (PST) when they adapt from living in freshwater to living in seawater (for review, see Dittman and Quinn, 1996). There is more recent evidence that some species of Pacific salmon, e.g. juvenile pink salmon (Oncorhynchus gorbuscha) and others, can imprint prior to the PST at a very young age (Bett et al., 2016; Dittman et al., 2015). Bett et al. (2016) published an excellent review on olfactory imprinting and olfactory guided navigation during spawning migrations in different salmon species and lampreys, elucidating imprinting cues and underlying mechanisms. However, there is also evidence that salmon imprint magnetically on their home area and this most likely brings them close to the home stream, with olfactory cues used mainly in the final part of the spawning migration (Putman et al., 2014, 2013).
In addition to natal water, anadromous fish also respond to water that is conditioned by conspecifics, which they remember and recognize years later, when they search for their natal river (for review, see Keefer and Caudill, 2014). There is evidence that salmonids are even capable of population-level discrimination (Courtenay et al., 1997; Doving et al., 1974; Groot et al., 1986; McBride et al., 1964; Nordeng, 1971; Nordeng and Bratland, 2006; Olsén, 1989; Quinn and Tolson, 1986; Selset and Doving, 1980), and even discrimination of siblings from non-siblings (Quinn and Busack, 1985; Quinn and Hara, 1986; Winberg and Ols, 1992). But such kin-structured associations are not confined to salmonids. Genetic analyses have shown that in numerous vertebrate and invertebrate species, populations do not consist of a random mix of individuals but of groups of related individuals of full- or half-siblings; kin associations have been demonstrated in many teleost species (for review, see Gerlach and Hinz, 2012; Selwyn et al.,
2016). To identify even unfamiliar kin, organisms must imprint on specific cues of kin, which they match with those expressed by unknown conspecifics ( phenotype matching) (Hepper, 1986; Tang-Martinez, 2001). Therefore, our findings (Gerlach et al., 2008; Gerlach and Lysiak, 2006; Hinz et al., 2013a,b) on the mechanisms underpinning olfactory imprinting in zebrafish (Danio rerio) might apply to a whole variety of species.
Behavioural basis of olfactory imprinting
Olfactory imprinting on conspecific and kin cues in zebrafish While selective advantages of kin recognition were studied in numerous species, the behavioural and neurobiological basis of the imprinting process are much less understood. To elucidate the behavioural and neuronal basis of the imprinting process, we took advantage of the model organism zebrafish (D. rerio), which imprint on an olfactory and visual template of kin early in development, leading to a long-term memory expressed as an olfactory-based differentiation between kin and non-kin (Hinz et al., 2013a).
Zebrafish raised in kin groups (siblings) identify even unfamiliar kin later in life, a process called‘phenotype matching’(Hepper, 1986). We found that larval and juvenile zebrafish preferred olfactory cues of kin when they were given the choice between water in which kin (siblings) or non-kin were kept; larvae were tested in a two-channel‘Atema’choice flume (Mann et al., 2003). The olfactory preference for kin changed with sexual maturity: adult females preferred the odour blend of unfamiliar non-related males over unfamiliar brothers, while adult males were attracted to all females, related and unrelated (Gerlach and Lysiak, 2006).
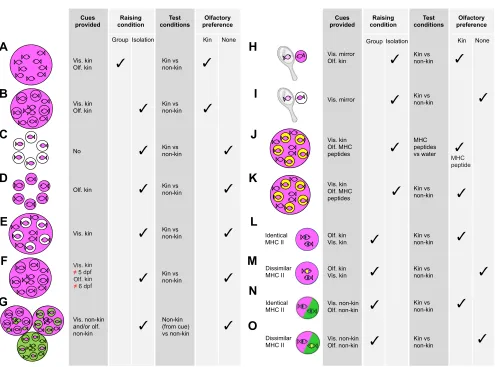
We raised larvae under different conditions to identify the time window of imprinting (see Fig. 1F). As experimentally isolated larvae (kept in glass beakers) did not express any olfactory preference in the Atema choice flume test, we concluded that they must be exposed to siblings during development. The sensitive period for olfactory templates of kin is limited to 24 h at 6 days post-fertilization (dpf ) (Gerlach et al., 2008) (Fig. 1A). When larvae were exposed to kin at other days for 24 h, but not at 6 dpf, they did not express any kin preference later in life (Fig. 1F). We also raised single larvae (hatching occurs at 4–5 dpf at a temperature of 25°C) in separate beakers and could imprint them when adding kin water at 6 dpf (Fig. 1B). Without such olfactory cues, they did not express any preference for kin (Fig. 1C,E). Interestingly, the application of non-kin water at 6 dpf did not result in a preference for non-kin odour (Fig. 1G). Therefore, zebrafish larvae must experience kin odour at 6 dpf for imprinting, and they cannot be imprinted on olfactory cues of non-kin.
Placing the glass beakers containing single larvae directly next to each other was very important, because it turned out that a visual template was essential for the imprinting and later olfactory-based kin recognition process. Without visual exposure to other kin, there was no kin preference and recognition later in life (Fig. 1D). Interestingly, visual cues of same-aged non-kin did not evoke imprinting (Fig. 1G). Again, we identified the time period for visual imprinting, which turned out to happen at 5 dpf (Hinz et al., 2013a). Both olfactory and visual cues were essential and larvae did not express any kin recognition when the cues were given at any other time than 5 dpf (visual cue) and 6 dpf (olfactory cue) or when originating from non-kin. These findings indicate that a genetic predisposition exists that results in imprinting when larvae experience the right cues.
Our results about the necessity of visual kin cues were intriguing. How can a larva know what its siblings look like when it has never seen any larvae before? What makes them know that the other larvae
Journal
of
Experimental
outside its own beaker are relatives? To understand this phenomenon, we raised single fertilized eggs/larvae in glass beakers surrounded by mirrors–we added holding water of kin at 6 dpf and obtained strong kin preference (Fig. 1H). Without adding kin odour, we did not observe kin recognition (Fig. 1I). We concluded that self-matching on chemical cues is not possible, but self-matching on visual cues is possible.
We wondered whether fish did not imprint on their own body odour because the concentration of urine from a single larva is too low to trigger imprinting. To test this possibility, we kept two full siblings in twice as much water as a single larva, to obtain the same concentration, and found that concentration had no effect: imprinting was observed when two siblings were kept together but not when a larva was kept individually, although a view of additional siblings outside the beaker was possible in both conditions.
To understand the visual part of imprinting, we took photos from the lateral side of larvae and compared the pigment pattern within and between sibling groups; this showed that related larvae share a similar pigmentation pattern (Hinz et al., 2012). The highest similarity among related larvae was found in the pigmentation pattern of the eye, indicating that the process is based on genetic disposition rather than direct comparison of own-body pigmentation with that of others. Larvae in a natural environment cannot see their
own eye pigmentation in contrast to larvae in the above-mentioned mirror experiments.
Advantage of kin groups
What is the selective force for olfactory imprinting and resulting kin recognition in zebrafish larvae? Larvae raised in groups of full-siblings showed an increased growth rate of 33% compared with larvae raised with non-kin (Gerlach et al., 2007b); however, we could not observe any differences in aggression or foraging. Therefore, association with kin and faster growth rates might lead to enhanced fitness by earlier reproduction. In Atlantic salmon and rainbow trout, kin group members had more foraging opportunities and experienced significantly greater weight gain than non-kin group members (Brown and Brown, 1996). In three-spined sticklebacks (Gasterosteus aculeatus), Frommen and Bakker (2004) suggested that shoaling with kin reduces competition, allows for more stable dominance hierarchies and increases fitness of relatives.
MHC class II peptide ligands as chemical signals evoking olfactory imprinting and kin recognition
In vertebrates, much effort has been made to reveal which cues are used to identify kin. The kin signal must fulfil two attributes: it must be sufficiently unique to avoid mistakes and it has to be stable in
Isolation
Group Kin None Vis. kin
Olf. kin
Vis. kin Olf. kin
No
Olf. kin
Vis. kin
Vis. kin ≠ 5 dpf Olf. kin ≠ 6 dpf
Vis. non-kin and/or olf. non-kin
Kin vs non-kin
Kin vs non-kin
Kin vs non-kin
Kin vs non-kin
Kin vs non-kin
Kin vs non-kin
Non-kin (from cue) vs non-kin
A
B
C
D
E
F
G
Cues provided
Raising condition
Olfactory preference Test
conditions
Isolation
Group Kin None
Cues provided
Raising condition
Olfactory preference Test
conditions
Vis. mirror Olf. kin
Vis. mirror
Vis. kin Olf. MHC peptides
Olf. kin Vis. kin
Olf. kin Vis. kin
Vis. non-kin Olf. non-kin
Vis. non-kin Olf. non-kin
Kin vs non-kin
Kin vs non-kin
MHC peptides vs water
Kin vs non-kin
Kin vs non-kin
Kin vs non-kin
Kin vs non-kin
MHC peptide
Vis. kin Olf. MHC peptides
Kin vs non-kin
Identical MHC II
Dissimilar MHC II
Dissimilar MHC II Identical MHC II
H
I
J
K
L
M
N
[image:3.612.60.555.50.417.2]O
Fig. 1. Summary of behavioural olfactory kin preference tests. A–O are modified from Hinz et al., 2012, 2013a,b. The diagrams on the left illustrate the raising conditions, which are described in the next two columns (vis., visual cues; olf., olfactory cues).‘Test conditions’describe chemical cues that larvae experienced during the test.‘Olfactory preference’indicates whether larvae expressed a statistically significant preference for kin or no preference; see‘Olfactory imprinting on conspecific and kin cues in zebrafish’for details. MHC II, major histocompatibility complex (class II).
Journal
of
Experimental
order to be recognized over space and time. In the aquatic environment, these are non-volatile olfactory cues. The expression of genes of the major histocompatibility complex (MHC) provides individuals with a unique body odour that is more similar among individuals with increasing genetic relatedness (Apanius et al., 1997; Boehm and Zufall, 2006; Penn, 2002; Singh, 2001). Imprinting on MHC and other relatedness cues released via urine, mucus and gills plays a major role in kin recognition in fish and other aquatic species (for review, see Gerlach and Hinz, 2012).
The olfactory preference of individual zebrafish larvae appears to be defined at least in part by their MHC class II genotype. We used a combination of artificial MHC peptides to study their potential role in imprinting (Hinz et al., 2013b). First, we searched for sibling groups, which developed a preference for a specific set of five randomly chosen synthetic MHC peptides when they had been exposed to these peptides during the critical period for imprinting. We found one zebrafish line (OL1, one pair and their offspring) that showed olfactory preference after exposure to these peptides at 6 dpf (Hinz et al., 2013b) (Fig. 1J). This exposure resulted not only in a subsequent preference for the MHC peptide mix but also in a preference for kin water (Fig. 1K), suggesting that the chosen MHC peptides are identical or similar to components of the kin-specific odour cues that are relevant for imprinting in this line (Fig. 1).
We further examined whether zebrafish can imprint on olfactory cues of other unrelated zebrafish that bear the same MHC genotype. The similarity of MHC class II but not MHC class I alleles between two zebrafish larvae corresponded to imprinting. Larvae did not discriminate between water derived from fish with identical MHC class II alleles even though they differed in genetic relatedness (Hinz et al., 2013b) (Fig. 1L–O). Hence, the MHC class II genotype appears to code for important olfactory cues for kin recognition and imprinting in zebrafish.
Interestingly, MHC class II allele sharing led to similarity not only in olfactory cues but also in pigmentation pattern. The degree of allele sharing of MHC class II genes defined body, rump, tail and iris pigmentation (Hinz et al., 2012).
We concluded that the MHC class II genotype could represent the underlying mechanism carrying the information about kin and non-kin and, thus, be the basis of the genetic predisposition (Hinz et al., 2013b).
Neuronal basis of olfactory imprinting Olfactory system and its signal transduction in fish
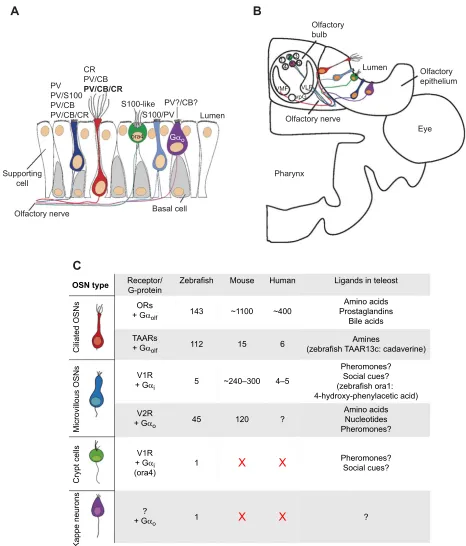
Unlike that in mammals, the olfactory system of the zebrafish as a teleost consists of only one main olfactory organ (Hansen et al., 2004; Hansen and Zeiske, 1993, 1998). Olfactory chemoreception is dependent on the binding of an odorant to its corresponding receptors, located on microvilli or cilia of olfactory sensory neurons (OSNs), with subsequent signal transmission into the central nervous system. Olfactory receptors (ORs) are located on the dendrites of OSNs embedded in the olfactory epithelium (OE) (see Fig. 2). Two major populations of OSNs are present in fish – microvillous OSNs (mOSNs) and ciliated OSNs (cOSNs) – resembling OSNs present in tetrapod vomeronasal and main olfactory systems (Eisthen, 1997; Korsching, 2016). Zebrafish feature two more OSN types: crypt cells (Ahuja et al., 2013; Hansen and Finger, 2000; Kress et al., 2015) and kappe cells (Ahuja et al., 2014)–both are named after their peculiar morphology and are so far only known to be present in fish (see Fig. 2). Odorant information is transduced into electrical signals inside the OSNs. OSNs in the nose express one olfactory receptor out of a large repertoire and project directly to the first olfactory processing centre
in the brain, the olfactory bulb (OB), where many of them converge into fewer glomeruli ( primary olfactory pathway) (Yoshihara, 2009). Bulbar projections further transmit odorant information into different areas of the telencephalon and diencephalon (secondary olfactory pathway) (Yoshihara, 2009). For example, in carp, mOSNs are mainly tuned towards food-related odours, whereas cOSNs play a role in mediating the alarm reaction, and crypt cells are suggested to be involved in reproduction (Hamdani et al., 2001, 2008; Hamdani and Døving, 2002, 2006). Electrophysiological data from Hansen et al. (2003) in channel catfish suggest that all OSN types respond to amino acids, with cOSNs additionally responding to bile salts, whereas mOSNs also respond to nucleotides. In goldfish, mOSNs are assumed to respond preferentially to amino acids (Speca et al., 1999). In transgenic zebrafish, blocking of synaptic transmission in distinct populations of mOSNs abolished attractive behavioural responses to amino acids (Koide et al., 2009), which is in line with the findings of other studies (Lipschitz and Michel, 2002). Amino acids and nucleotides are typically indicative of food whereas bile salts are considered as social odorants as bile salt profiles within a teleost family and order show high similarity (Hagey et al., 2010). However, a general conclusion on teleost OSN tuning is hard to make as the response profiles of OSNs differ, as do bulbar projection patterns across species (Bazáes et al., 2013).
Experience of olfactory cues can induce changes in the olfactory neuronal system
The olfactory system of teleosts enables the recognition and discrimination of a vast number of environmental and social odours. In the aquatic realm, olfactory-based behavioural context includes migration (Yamamoto et al., 2010), association with kin (Hinz et al., 2013b; Olsen et al., 2002), feeding (Miklavc and Valentincic, 2012), predator avoidance (Idler et al., 1956), response to alarm signals (Schreckstoff) (Speedie and Gerlai, 2008) and social interactions such as mating and reproduction (Veyrac et al., 2011). The olfactory system of fish detects and discriminates between structurally different classes of odorants: mainly charged molecules including amino acids, bile acids, nucleotides, steroids and prostaglandins, but also uncharged volatile molecules (Korsching, 2016). Responses to amino acids, bile acids and nucleotides (Carr, 1988) emerge early in development, shortly before hatching (Li et al., 2005). In zebrafish and other teleosts, defined prostaglandins and steroids serve as pheromonal signals prior to mating (Kobayashi et al., 2002; Moore and Waring, 1996; Sorensen et al., 1988; Stacey et al., 1989; Yabuki et al., 2016), and substances released from the skin evoke an alarm response (Jesuthasan and Mathuru, 2008). Most of the behavioural responses to these stimuli arise late in development, but the emergence of physiological and neurobiological responses to these stimuli has not been studied in detail. However, Braubach et al. (2013) provided exciting evidence of an activity-dependent developmental mechanism in the zebrafish olfactory system, which might be important to understand olfactory imprinting. They suggested that imprinting could trigger neuronal changes in the OB and perhaps in the expression of specific olfactory receptor genes dependent on contact with olfactory cues at a specific time during development. Ochs et al. (2017) described the morphology of glomerular patterning in wild Chinook salmon during a temporal window previously shown to be significant for early olfactory imprinting.
Sockeye salmon (Oncorhynchus nerka) that successfully imprinted onL-arginin, as evidenced by adult behaviour, demonstrated increased expression (relative to arginine-naive fish) of the putative arginine receptor mRNA in the OE during key life stages (Dittman et al., 2010). In brains of hatchery-reared juvenile chum salmon (Oncorhynchus
Journal
of
Experimental
C
OSN type Receptor/ Zebrafish G-protein
ORs
+ Gαolf 143
Microvillous OSNs
Ciliated OSNs
Crypt cells
Kappe neurons
112 15 6
5 ~240–300 4–5
45
1
1
120 ?
?
~1100 ~400
TAARs
+ Gαolf
V1R
+ Gαi
V2R
+ Gαo
V1R
+ Gαi
(ora4)
?
+ Gαo
Mouse Human
Amino acids Prostaglandins
Bile acids
Amines
(zebrafish TAAR13c: cadaverine)
Pheromones? Social cues? (zebrafish ora1: 4-hydroxy-phenylacetic acid)
Amino acids Nucleotides Pheromones?
Pheromones? Social cues? Ligands in teleost
X
X
X
X
C
OSN type Receptor/ Zebrafish G-protein
ORs
+ Gαolf 143
M
icrovillous
OS
N
s
C
iliated
OS
Ns
C
rypt cells
KK
a
pp
e neurons
112 15 6
5 ~240–300 4–5
45
1
1
120 ?
?
~1100 ~400
TAARs
+ Gαolf
V1R
+ Gαi
V2R
+ Gαo
V1R
+ Gαi
(ora4)
?
+ Gαo
Mouse Human
Amino acids Prostaglandins
Bile acids
Amines
(zebrafish TAAR13c: cadaverine)
Pheromones? Social cues? (zebrafish ora1: 4-hydroxy-phenylacetic acid)
Amino acids Nucleotides Pheromones?
Pheromones? Social cues? Ligands in teleost
X
X
X
X
CR PV/CB
PV/CB/CR
PV PV/S100 PV/CB PV/CB/CR
S100-like
Supporting cell
Olfactory nerve Basal cell
Pharynx
A
B
Eye Lumen
VLF
1 2 3
4 5 6
vpG VMF
Olfactory epithelium
Olfactory nerve Olfactory bulb
Lumen S100/PV
ora4 Gα
o
[image:5.612.72.538.63.609.2]PV?/CB?
Fig. 2. Zebrafish odorant receptors and olfactory sensory neurons (OSNs).(A) Olfactory epithelial positions of zebrafish OSNs. Combinatorial analysis of calcium-binding protein (CBP) expression reveals at least eight subpopulations of OSNs. Bold font indicates the largest of three subpopulations of ciliated OSNs ( parvalbumin/calbindin/calretinin, PV/CB/CR; red). Four subpopulations of microvillous OSNs (mOSNs, dark blue) express PV. CBP S100 is specifically present in all crypt cells (green) and a minor subpopulation of mOSNs, which also express PV (shown again on the right of crypt cells in light blue). Kappe neurons ( purple) probably express PV and CB and probably correspond to the PV/CB-mOSN population. (B) Zebrafish olfactory sensory cell types defined by morphology and CBPs. Schematic drawing of a cross-section through a larval zebrafish olfactory epithelium and olfactory bulb of one body side. The mediodorsal glomerular field (mdG) is indicated by six glomeruli (1–6). A hypothetical representation of the crypt cells (green) projecting to mdG2 is shown. Additional axonal input to this glomerulus comes from S100/PV-expressing microvillous OSNs (light blue). A second glomerulus (out of the 6) within the mediodorsal domain, mdG5 (5), receives input from PV/CB-expressing OSNs, probably representing kappe neurons. Also shown are the ventroposterior glomerulus (vpG) and the ventrolateral (VLF) and ventromedial (VLM) glomerular fields subserved by mOSN and cOSN input, respectively. See text and Kress et al. (2015) for more details. (C) Comparison of zebrafish olfactory receptors and OSNs with those in mouse and human (for sources of data, see‘Olfactory system and its signal
transduction in fish’).
Journal
of
Experimental
keta), thyrotropin-releasing hormone gene expression increased immediately after release from a hatchery into the natal stream, and the expression of the essential NR1 subunit of the N-methyl-D -aspartate receptor (NMDAR) increased during downstream migration (Ueda et al., 2016). The NMDAR is a glutamate receptor channel subtype and mediates most of the fast-excitatory synaptic transmission in the central nervous system. It plays important roles in memory formation and retrieval in fish (Gómez et al., 2006; Kinoshita et al., 2004). Gene expression of salmon gonadotropin-releasing hormone (sGnRH) and NR1 increased in the adult chum salmon brain during homing from the Bering Sea to the natal hatchery (Ueda et al., 2016). However, in contrast to salmonids, in zebrafish, we could not evoke any olfactory preference for a variety of amino acids after larvae were exposed to different concentrations of these amino acids during development (K.T. and G.G., unpublished data). However, zebrafish as a non-migrating fish might imprint on other cues.
Tuning of teleost OSNs and processing of kin odour in the zebrafish OE
To investigate which type of OSN detects kin odour, we mapped neuronal activity following olfactory stimulation visualized by pERK upregulation in OSNs in the OE of 9 dpf zebrafish larvae (Biechl et al., 2016). pERK is widely used to mark neuronal activity in mammals; for example, in the olfactory system following odour stimulation in mice (Mirich et al., 2004). Upon phosphorylation, pERK is translocated into the nucleus of the activated cell to modulate the expression of transcription factors, which in turn regulate the expression of genes involved in neuronal and synaptic plasticity underlying learning and memory. pERK is preferable to other markers, such as immediate early genes (e.g.c-fosoregr1), because of its more rapid activation and its cellular distribution (soma and cell protrusions) (Gao and Ji, 2009; Randlett et al., 2015). Olfactory stimulation with different odours (food, conspecific odour or E3-medium) showed a differential pattern of activated OSNs within the larval OE (Biechl et al., 2016). High numbers of cOSNs – and a smaller number of mOSNs – were activated in response to food odour exposure, whereas mOSNs almost exclusively showed pERK upregulation following exposure to a conspecific odour (Biechl et al., 2016). Interestingly, crypt cells showed neuronal activation in response to neither food nor conspecific (non-kin) odour. These response patterns were generally consistent with data from other studies, despite great interspecific variability regarding the tuning of OSNs within teleosts (Bazáes et al., 2013).
By rearing larvae isolated in small glass beakers in a larger tank, we either prevented olfactory imprinting on day 6 or allowed the larvae to imprint on their kin by adding kin odour containing water to the beakers (Biechl et al., 2016). The rearing conditions for all larvae were otherwise identical (e.g. changing water, feeding and isolation), precluding other influences on the results. Importantly, despite raising larvae in glass beakers, visual imprinting at 5 dpf was possible because isolated larvae could see their siblings swimming outside their own beakers in the larger tank (see also Hinz et al., 2013a). Additionally, in the subsequent histological assay for pERK we used the calcium-binding protein (CBP) S100 to specifically identify crypt cells. In the crucian carp (Carassius carassius), crypt cells project into the ventral OB from where projection neurons terminate in the lateral part of the medial olfactory tract, which is known to mediate reproductive behaviours (Weltzien et al., 2003). Furthermore, in sexually mature carp, crypt cells were shown to vary in their density as well as location within the OE depending on the season (Hamdani and Døving, 2006; Hamdani et al., 2008). Carp
exhibited only a few such cells during winter whereas in the summer spawning season, carp crypt cells were clearly detectable and positioned at the surface of the OE, suggesting that they were involved in carp reproductive behaviour (Hamdani et al., 2008). Sandulescu et al. (2011) demonstrated an interesting time course of zebrafish crypt cell quantity during OE development. Upon first appearance in the zebrafish OE at 4 dpf, the number of crypt cells increased steadily until reaching a peak at 7 dpf. Comparing this growth of crypt cells with the sensitive phase of olfactory imprinting (day 6), it appears as though crypt cell numbers might adapt to the upcoming imprinting event.
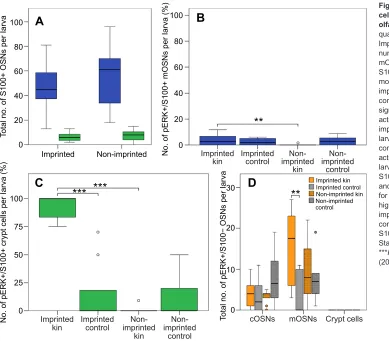
Based on these studies, we exposed imprinted and non-imprinted zebrafish larvae at 9 dpf to kin odour; only crypt cells of imprinted larvae showed activation in response to kin odour (Fig. 3C; Biechl et al., 2016). Importantly, this was not due to an absence of this cell type, as crypt cell numbers did not differ between non-imprinted and imprinted larvae (Biechl et al., 2016) (Fig. 3A). Furthermore, microarray data provided no evidence for a down-regulation of ORA4 receptor expression in crypt cells in non-imprinted compared with imprinted zebrafish larvae (K.T. and G.G., unpublished data). Thus, the critical cue, which is obviously contained in kin odour, apparently changed the responsiveness of crypt cells in an unknown manner during the imprinting process. Crypt cells express only a single V1R-like receptor, encoded by theora4gene (Oka et al., 2012). The ligand that binds to ORA4 is still unknown; however, based on our data, the ligand of ORA4 is contained in kin odour but it is unlikely that ORA4 is the specific receptor for MHC peptides in zebrafish (Behrens et al., 2014; Boschat et al., 2002; Isogai et al., 2011).
In addition to crypt cells, a small subpopulation of (S100-negative) mOSNs was shown to respond to kin odour (Fig. 3D). Considering that MHC peptides are a component of kin odour, these mOSNs might express a V2R receptor and bind to such peptides. Regardless, kin odour is composed of a mixture of numerous odorants that signal –besides genetic relatedness–additional social information such as sex and physiological status, and possibly kin recognition is mediated by more than one chemical compound. Thus, a receptor code, generated by the interaction of multiple receptors and activated by more than one ligand, might signal information about relatedness in larval zebrafish. In sum, our studies support the hypothesis that crypt cells as well as a small subpopulation of mOSNs are involved in detecting a kin odour-related signal.
Potential neuronal changes due to olfactory imprinting
Nevitt et al. (1994) hypothesized a model for olfactory imprinting in salmon. They suggested that during sensitive imprinting periods, thyroid hormones stimulate the proliferation of non-specific olfactory sensory neurons. Subsequently, the receptors that are most responsive to the odorants survive and the non-responsive receptors die. This model was based on previous studies that demonstrated odorant-specific sensitization of the peripheral olfactory system in coho salmon (Oncorhynchus kisutch) (Dittman et al., 1997). Their results suggested that exposure of coho salmon to an odorant for 10 days during smolting could result in significant changes in the peripheral sensitivity to that odorant in the spawning adult coho salmon (Dittman et al., 2010, 1997).
We tested whether the expression of odorant receptors and other genes in zebrafish larvae changed during the sensitive phase of olfactory imprinting through a genome-wide transcriptional analysis using microarray chip that included gene-level probe sets for 129 OR, 82 TAAR, 46 OlfC and 6 ORA odorant receptor genes, covering all knownoragenes, but missing probe sets for some odorant receptor genes from the TAAR, OR and OlfC group (K.T. and G.G.,
Journal
of
Experimental
unpublished results). Microarray analysis showed no differences in gene expression for 263 odorant receptor genes in imprinted versus non-imprinted larvae (K.T. and G.G., unpublished results); however, we might have missed specific odorant receptors especially some TAARs, which are suggested to be associated with the detection of social cues in mice (Ziegler et al., 2002).
Zebrafish larvae exposed to the artificial odorant β-phenethyl alcohol (βPEA) showed an increase in expression of the transcription factor Otx2 inside the OE (Calfún et al., 2016; Harden et al., 2006). The authors suggested that βPEA triggers olfactory imprinting in juvenile zebrafish and further showed that three OR genes (OR111-1, OR115-5, OR125-1) are affected by Otx2 expression.
Our qPCR analysis in 5, 6 and 7 dpf zebrafish larvae (before, during and after olfactory imprinting) revealed no differences in Otx2 expression; the same finding was obtained for imprinted and non-imprinted larvae (K.T., G.S and A.S., unpublished results). In situ hybridization experiments confirmed our findings: no differences in Otx2 expression pattern were observed in 5, 6 and 7 dpf imprinted and non-imprinted larvae.
Spatial distribution of odour responses in the OB
To explore how olfactory cues relevant for imprinting are represented in the zebrafish brain, we measured odour-evoked activity in the OB and adjacent areas by multiphoton calcium imaging (Hinz et al., 2013b). Patterns of input activity across the array of glomeruli are processed by neuronal circuits in the OB that are composed of principal neurons, the mitral cells and multiple types of interneurons. General odorants such as amino acids, bile acids and nucleotides stimulate multiple glomeruli, and stimuli of the same class often evoke
responses in overlapping, yet stimulus-specific, subpopulations of OB neurons (Friedrich, 2013).
The spatial distribution of odour responses is usually broad but not entirely random because responses to odours of the same class are biased towards broad sub-regions. For example, responses to amino acids and bile acids are found predominantly in the ventrolateral and the dorsomedial OB, respectively (Doving et al., 1980; Friedrich and Korsching, 1997, 1998a; Yaksi et al., 2009). This coarse functional topography of odour responses is established at early stages of development and maintained throughout adulthood (Li et al., 2005). Pheromones, in contrast, have been reported to activate few glomeruli with high specificity (Friedrich and Korsching, 1998b; Yabuki et al., 2016). Moreover, pheromones and general odorants activate different brain areas downstream of the OB (Yabuki et al., 2016), consistent with the hypothesis that pheromonal signals are detected by specific receptors and control distinct behavioural or endocrine responses.
Neuronal representation of olfactory kin cues in the OB
In mammals, MHC peptides convey information about individuality and stimulate sensory neurons of the main and accessory olfactory systems (Leinders-Zufall et al., 2004; Spehr et al., 2006). Individual sensory neurons respond to multiple peptides, indicating that stimulus identity is encoded by a combination of activity patterns across multiple odorant receptors and glomerular channels. However, the spatial distribution of responses to MHC peptides in the OB has not been analysed in detail. To address this, we mapped odour responses to a mixture of the five MHC peptides that evoked olfactory imprinting and kin recognition in a specific kin group (Hinz et al., 0
0 25 50 75 100
0 10 20 30
Imprinted Non-imprinted Imprinted
kin
Imprinted control
Non-imprinted
kin
Imprinted kin Imprinted control Non-imprinted kin Non-imprinted control
Non-imprinted
control
Imprinted kin
Imprinted control
Non-imprinted
kin
Non-imprinted
control
cOSNs mOSNs Crypt cells 20
T
o
tal no. of S100+ OSNs per Iarva
No. of pERK+/S100+ crypt cells per Iarva (%) To
tal no. of pERK+/S100− OSNs per Iarva
No. of pERK+/S100+ mOSNs per Iarva (%)
40 60 80
100
A
B
C
D
0 20 40
**
***
**
***
[image:7.612.49.438.56.397.2]60 80 100
Fig. 3. Differential activation of crypt cells with kin odour–effects of olfactory imprinting.(A) Total cell quantity of S100+ mOSNs and crypt cells. Imprinting has no effect on total cell number. (B) Number of S100+/pERK+ mOSNs shown as a percentage of all S100+ mOSNs per larva. Significantly more S100+ mOSNs are activated in imprinted larvae versus non-imprinted control larvae exposed to kin odour. (C) A significantly higher number of crypt cells is activated after kin odour stimulation in imprinted compared with non-imprinted larvae and compared with imprinted control larvae stimulation. No difference in activation was found within non-imprinted larvae. (D) The total number of pERK+, but S100−, ciliated OSNs (cOSNs), mOSNs and crypt cells. Cell activation was similar for all treatments in cOSNs. A significantly higher number of mOSNs was found in imprinted larvae stimulated with kin compared with control stimulation. No S100−crypt cells were observed. Statistical significance: **P<0.01, ***P<0.001. Modified from Biechl et al. (2016).
Journal
of
Experimental
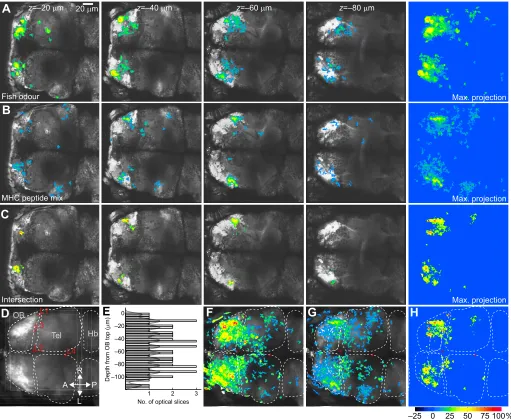
2013b) in the OB by multiphoton calcium imaging in transgenic fish expressing the genetically encoded calcium indicator GCaMP2 under the control of the pan-neuronal elavl3 promoter (Ahrens et al., 2012) (Fig. 4). Because zebrafish larvae are small and transparent, activity could be imaged in multiple focal planes, separated by 10 µm, that covered the entire OB and parts of the telencephalon and habenulain vivo. Results from multiple fish were combined by registration of image data to a common reference brain (Fig. 4D). We found that responses to MHC peptides were distributed and not confined to a specific location, although responses appeared to be particularly dense in a sub-volume of the ventrolateral, amino acid-responsive part of the OB (Fig. 4) (Hinz et al., 2013b).
Unlike responses to amino acid odours, responses to MHC peptides did not substantially broaden with increasing stimulus concentration (Hinz et al., 2013b), consistent with observations in mammalian sensory neurons (Leinders-Zufall et al., 2004). The best response was observed at very low concentrations of MHC peptide mix (1.25×10−12mol l−1) (Hinz et al., 2013b). Activity patterns evoked by MHC peptides showed significant overlap with activity evoked by the odour of conspecifics (Fig. 4), but less overlap with activity evoked by food extract (Hinz et al., 2013b), even though food extract is a strong and complex odour. These observations support the hypothesis that MHC peptides are released into the water and detected by the olfactory system of zebrafish. Moreover,
Depth from OB top (
μ
m)
A
B
C
D
OBOBBE
F
Tel
T Hb
L1
L2 L3
L4
G
H
R
L
A P
Max. projection
Max. projection z=–20 μm z=–40 μm z=–60 μm z=–80 μm
Max. projection
–25 0 25 50 75 100%
–60 –80 –100 –40 –20 0
1 2 3 No. of optical slices
Fish odour
MHC peptide mix
Intersection
[image:8.612.50.561.206.625.2]20 μm
Fig. 4. Spatial distribution of responses to MHC peptides and fish odour.(A,B) Expression of GCaMP2 (greyscale) and odour-evoked relative change in fluorescence (colour; thresholded) in four optical sections of transgenic zebrafish. A depth ofz=0 corresponds to the dorsal pole of the olfactory bulb; negative values indicate more ventral locations. Right: maximum projection of responses in 10 optical planes spaced at 10 µm. Stimuli were water conditioned by conspecifics (fish odour; A) and a mix of five MHC peptides (B). All fish were raised in the presence of siblings. In trial- and time-averaged response maps, pixels were thresholded at 2.5 s.d. and contiguous areas of less than 20 pixels (∼2.8×2.8 µm2) were deleted. (C) Intersection of activity patterns evoked by fish
odour and the MHC peptide mix. In each optical section, response intensities were summed when relative changes in fluorescence exceeded 2.5 s.d. in response to both stimuli. Right: maximum projection of responses in 10 optical sections spaced at 10 µm. (D) Alignment of images from multiple individuals. The four landmarks L1–L4 were aligned by translation, scaling and rotation of image data. OB, olfactory bulb; Tel, telencephalon; Hb, habenula; A, anterior; P, posterior; R, right; L, left. (E) Depth histogram of optical planes imaged in six fish (10 µm spacing). (F–H) Thresholded and averaged responses to fish odour (F) or to MHC peptide mix (G); (H) intersection. Maximum projections through 10–11 optical planes in each of six fish were aligned and combined by taking maximum values. Optical sections covered the entire olfactory bulb but only parts of the telencephalon and habenula.
Journal
of
Experimental
unlike pheromones, MHC peptides do not appear to be detected by a small number of narrowly tuned sensory channels. Rather, MHC peptides appear to be represented in a combinatorial fashion by activity patterns across neuronal populations, consistent with observations in the peripheral olfactory system of mammals (Leinders-Zufall et al., 2004; Spehr et al., 2006).
As the observed higher neuronal activity in specific OSNs (crypt cells and mOSNs) in imprinted larvae is not explained by an increased number of these OSNs or a change in odorant receptor gene expression, olfactory imprinting might be explained by a change in sensitivity of the odorant receptor itself. A ligand–MHC protein– odorant receptor interaction might lead to a stronger neuronal activation compared with that in larvae that are exposed to peptide ligands of non-kin odour. Odorant-binding proteins can enhance the solubility of hydrophobic odours and facilitate the transport of odours to receptor sites (Pelosi, 1994). Even if this does not represent a modulation of receptor affinity, it still enhances the possibility of a ligand–receptor interaction and thereby might represent a station of neuronal activity modulation in general.
Our approach to study processes that underlie olfactory imprinting and kin recognition in a non-migratory but much more tractable fish should shed light on the mechanistic underpinnings of imprinting in parental populations in migratory fishes. But olfactory imprinting on kin versus environmental cues seems to differ, as shown by differences in olfactory receptor gene expression.
Acknowledgements
Some parts of the results and discussion are taken from the PhD thesis of Daniela Biechl (Ludwig Maximilian University, Munich, 2018). We thank Sigrun Korsching for fruitful discussions. We also thank B. Grothe for additional support. We thank Pascal Fieth for bioinformatics support for the microarray gene expression analysis. We are grateful to R. Friedrich and Mario Wullimann for their contributions to the discussion.
Competing interests
The authors declare no competing or financial interests.
Funding
Funding was provided by the Deutsche Forschungsgemeinschaft DFG (SPP 1392 Integrative Analysis of Olfaction; GE 842/5-1, 5-2, FR 1667/2-1, 2-2 and WU211/2-1, 2-2).
References
Ahrens, M. B., Li, J. M., Orger, M. B., Robson, D. N., Schier, A. F., Engert, F. and
Portugues, R.(2012). Brain-wide neuronal dynamics during motor adaptation in
zebrafish.Nature485, 471-477.
Ahuja, G., Ivandic, I., Salturk, M., Oka, Y., Nadler, W. and Korsching, S. I.(2013).
Zebrafish crypt neurons project to a single, identified mediodorsal glomerulus.Sci. Rep.3, 2063.
Ahuja, G., Nia, S. B., Zapilko, V., Shiriagin, V., Kowatschew, D., Oka, Y. and
Korsching, S. I.(2014). Kappe neurons, a novel population of olfactory sensory
neurons.Sci. Rep.4, 4037.
Apanius, V., Penn, D., Slev, P. R., Ruff, L. R. and Potts, W. K.(1997). The nature of
selection on the major histocompatibility complex. Crit. Revi. Immunol. 17, 179-224.
Armsworth, P. R.(2000). Modelling the swimming response of late stage larval reef
fish to different stimuli.Mar. Ecol. Progr. Ser.195, 231-247.
Atema, J., Kingsford, M. J. and Gerlach, G.(2002). Larval reef fish could use
odour for detection, retention and orientation to reefs.Mar. Ecol. Prog. Ser.241, 151-160.
Bazáes, A., Olivares, J. and Schmachtenberg, O.(2013). Properties, projections,
and tuning of teleost olfactory receptor neurons.J. Chem. Ecol.39, 451-464.
Behrens, M., Frank, O., Rawel, H., Ahuja, G., Potting, C., Hofmann, T., Meyerhof,
W. and Korsching, S.(2014). ORA1, a zebrafish olfactory receptor ancestral to all
mammalian V1R genes, recognizes 4-Hydroxyphenylacetic acid, a putative reproductive pheromone.J. Biol. Chem.289, 19778-19788.
Bett, N. N., Hinch, S. G., Dittman, A. H. and Yun, S. S.(2016). Evidence of
olfactory imprinting at an early life stage in pink salmon (Oncorhynchus gorbuscha).Sci. Rep.6, 36393.
Biechl, D., Tietje, K., Gerlach, G. and Wullimann, M. F.(2016). Crypt cells are
involved in kin recognition in larval zebrafish.Sci. Rep.6, 24590.
Boehm, T. and Zufall, F.(2006). MHC peptides and the sensory evaluation of
genotype.Trends Neurosci.29, 100-107.
Boschat, C., Pelofi, C., Randin, O., Roppolo, D., Luscher, C., Broillet, M. C. and
Rodriguez, I.(2002). Pheromone detection mediated by a V1r vomeronasal
receptor.Nature Neurosci.5, 1261-1262.
Bottesch, M., Gerlach, G., Halbach, M., Bally, A., Kingsford, M. J. and
Mouritsen, H.(2016). A magnetic compass that might help coral reef fish
larvae return to their natal reef.Curr. Biol.26, R1266-R1267.
Braubach, O. R., Miyasaka, N., Koide, T., Yoshihara, Y., Croll, R. P. and Fine, A.
(2013). Experience-dependent versus experience-independent postembryonic development of distinct groups of zebrafish olfactory glomeruli.J. Neurosci.33, 6905-6916.
Brown, G. E. and Brown, J. A.(1996). Does kin-biased territorial behavior increase
kin-based foraging in juvenile salmonids.Behav. Ecol.7, 24-29.
Burgess, S. C., Kingsford, M. J. and Black, K. P.(2007). Influence of tidal eddies
and wind on the distribution of presettlement fishes around One Tree Island, Great Barrier Reef.Mar. Ecol. Prog. Ser.341, 233-242.
Calfún, C., Domı́nguez, C., Pérez-Acle, T. and Whitlock, K. E.(2016). Changes in
olfactory receptor expression are correlated with odor exposure during early development in the zebrafish (Danio rerio).Chem. Senses41, 301-312.
Carr, W. E. S.(1988). The molecular nature of chemical stimuli in the aquatic
environment. InSensory biology of aquatic animals(ed. J. Atema, R. R. Fay, A. N. Popper and W. N. Tavolga), pp. 3-27. New York: Springer.
Courtenay, S. C., Quinn, T. P., Dupuis, H. M. C., Groot, C. and Larkin, P. A.
(1997). Factors affecting the recognition of population-specific odours by juvenile coho salmon.J. Fish Biol.50, 1042-1060.
Cowen, R. K., Lwiza, K. M. M., Sponaugle, S., Paris, C. B. and Olson, D. B.
(2000). Connectivity of marine populations: open or closed? Science 287, 857-859.
Dittman, A. H. and Quinn, T. P.(1996). Homing in pacific salmon: mechanisms and
ecological basis.J. Exp. Biol.199, 83-91.
Dittman, A. H., Quinn, T. P., Nevitt, G. A., Hacker, B. and Storm, D. R.(1997).
Sensitization of olfactory guanylyl cyclase to a specific imprinted odorant in coho salmon.Neuron19, 381-389.
Dittman, A. H., May, D. and Michelle, A.(2010). Odorant-Induced changes in
olfactory receptor mRNA expression in Sockeye salmon (Oncorhynchus nerka) after imprinting.Integ. Comp. Biol.50, E44-E44.
Dittman, A. H., Pearsons, T. N., May, D., Couture, R. B. and Noakes, D. L. G.
(2015). imprinting of hatchery-reared salmon to targeted spawning locations: a new embryonic imprinting paradigm for hatchery programs. Fisheries 40, 114-123.
Doving, K. B., Nordeng, H. and Oakley, B.(1974). Single unit discrimination of fish
odours released by char (Salmo alpinusL.) populations.Comp. Biochem. Physiol. 47A, 1051-1063.
Doving, K. B., Selset, R. and Thommesen, G.(1980). Olfactory sensitivity to bile
acids in salmonid fishes.Acta Physiol. Scand.108, 123-131.
Eisthen, H. L.(1997). Evolution of vertebrate olfactory systems.Brain Behav. Evol
50, 222-233.
Fisher, R. and Wilson, S. K.(2004). Maximum sustainable swimming speeds of
late-stage larvae of nine species of reef fishes.J. Exp. Mar. Biol. Ecol.312, 171-186.
Fisher, R., Bellwood, D. R. and Job, S. D.(2000). Development of swimming
abilities in reef fish larvae.Mar. Ecol. Prog. Ser.202, 163-173.
Friedrich, R. W.(2013). Information processing in the olfactory system of zebrafish.
Ann. Rev. Neurosci.36, 383-402.
Friedrich, R. W. and Korsching, S. I.(1997). Combinatorial and chemotopic
odorant coding in the zebrafish olfactory bulb visualized by optical imaging.
Neuron18, 737-752.
Friedrich, R. W. and Korsching, S. I.(1998a). Chemotopic, combinatorial, and
noncombinatorial odorant representations in the olfactory bulb revealed using a voltage-sensitive axon tracer.J. Neurosci.18, 9977-9988.
Friedrich, R. W. and Korsching, S. I.(1998b). Odorant representation in the
zebrafish olfactory bulb visualized by optical recording techniques: imaging odor images.Eur. J. Neurosci.10, 435-435.
Frommen, J. G. and Bakker, T. C. M.(2004). Adult three-spined sticklebacks prefer
to shoal with familiar kin.Behaviour141, 1401-1409.
Gao, Y.-J. and Ji, R.-R.(2009). c-Fos and pERK, which is a better marker for
neuronal activation and central sensitization after noxious stimulation and tissue injury?Open Pain J.2, 11-17.
Gerlach, G. and Hinz, C.(2012). Chemical signals and kin biased behaviour. In
Chemical Ecology in Aquatic Systems(ed. L. A. Hansson and C. Brönmark), pp. 58-71. Oxford: University Press.
Gerlach, G. and Lysiak, N.(2006). Kin recognition and inbreeding avoidance in
zebrafish,Danio rerio, is based on phenotype matching. Anim. Behav. 71, 1371-1377.
Gerlach, G., Atema, J., Kingsford, M. J., Black, K. P. and Miller-Sims, V.(2007a).
Smelling home can prevent dispersal of reef fish larvae.Proc. Natl. Acad. Sci.
USA104, 858-863.
Journal
of
Experimental
Gerlach, G., Hodgins-Davis, A., MacDonald, B. and Hannah, R. C.(2007b). Benefits of kin association: related and familiar zebrafish larvae (Danio rerio) show improved growth.Behav. Ecol. Sociobiol.61, 1765-1770.
Gerlach, G., Hodgins-Davis, A., Avolio, C. and Schunter, C. (2008). Kin
recognition in zebrafish: a 24-h window for olfactory imprinting.Proc. Roy. Soc. London B275, 2165-2170.
Gerlach, G., Atema, J., Raupach, M. J., Deister, F., Müller, A. and Kingsford,
M. J. (2016). Cryptic species of cardinalfish with evidence for old and new
divergence.Coral Reefs35, 437-450.
Gómez, Y., Vargas, J. P., Portavella, M. and López, J. C.(2006). Spatial learning
and goldfish telencephalon NMDA receptors. Neurobiol. Learn. Memory 85, 252-262.
Groot, C., Quinn, T. P. and Hara, T. J.(1986). Responses of migrating adult
sockeye salmon (Oncorhynchus nerka) to population specific odors.Can. J. Zool. 64, 926-932.
Hagey, L. R., Møller, P. R., Hofmann, A. F. and Krasowski, M. D.(2010). Diversity
of bile salts in fish and amphibians: evolution of a complex biochemical pathway.
Physiol. Biochem. Zool.83, 308-321.
Hamdani, E. H. and Døving, K. B.(2002). The alarm reaction in crucian carp is
mediated by olfactory neurons with long dendrites.Chem. Senses27, 395-398.
Hamdani, E. H. and Døving, K. B.(2006). Specific projection of the sensory crypt
cells in the olfactory system in crucian carp,Carassius carassius.Chem. Senses 31, 63-67.
Hamdani, E. H., Alexander, G. and Døving, K. B.(2001). Projection of sensory
neurons with microvilli to the lateral olfactory tract indicates their participation in feeding behaviour in crucian carp.Chem. Senses26, 1139-1144.
Hamdani, E. H., Lastein, S., Gregersen, F. and Doving, K. B.(2008). Seasonal
variations in olfactory sensory neurons - Fish sensitivity to sex pheromones explained?Chem. Senses33, 119-123.
Hansen, A. and Finger, T. E.(2000). Phyletic distribution of crypt-type olfactory
receptor neurons in fishes.Brain Behav. Evol.55, 100-110.
Hansen, A. and Zeiske, E.(1993). Development of the olfactory organ in the
zebrafish,Brachydanio rerio.J. Comp. Neurol.333, 289-300.
Hansen, A. and Zeiske, E.(1998). The peripheral olfactory organ of the zebrafish,
Danio rerio: an ultrastructural study.Chem. Senses23, 39-48.
Hansen, A., Rolen, S. H., Anderson, K., Morita, Y., Caprio, J. and Finger, T. E.
(2003). Correlation between olfactory receptor cell type and function in the channel catfish.J. Neurosci.23, 9328-9339.
Hansen, A., Anderson, K. T. and Finger, T. E.(2004). Differential distribution of
olfactory receptor neurons in goldfish: Structural and molecular correlates.
J. Comp. Neurol.477, 347-359.
Harden, M. V., Newton, L. A., Lloyd, R. C. and Whitlock, K. E.(2006). Olfactory
imprinting is correlated with changes in gene expression in the olfactory epithelia of the zebrafish.J. Neurobiol.66, 1452-1466.
Hasler, A. D., Scholz, A. T. and Horrall, R. M.(1978). Olfactory imprinting and
homing in salmon.Am. Sci.66, 347-355.
Hepper, P. G.(1986). Kin recognition-functions and mechanisms a review.Biol.
Rev. Cambridge Phil. Soc.61, 63-93.
Hinz, C., Gebhardt, K., Hartmann, A. K., Sigman, L. and Gerlach, G.(2012).
Influence of kinship and MHC class II genotype on visual traits in zebrafish larvae (Danio rerio).PLoS ONE7, e5118210.137.
Hinz, C., Müller, A., Kobbenbring, S., Sigman, L. and Gerlach, G.(2013a). Kin
recognition in zebrafish,Danio rerio, is based on imprinting on olfactory and visual stimuli.Anim. Behav.85, 925-930.
Hinz, C., Namekawa, I., Behrmann-Godel, J., Oppelt, C., Jaeschke, A., Müller,
A., Friedrich, R. W. and Gerlach, G.(2013b). Olfactory imprinting is triggered by
MHC peptide ligands.Sci. Rep.3, 2800.
Idler, D. R., Fagerlund, U. H. M. and Mayoh, H.(1956). Olfactory perception in
migrating salmon. I. L-serine, a salmon repellent in mammalian skin.J. Gen. Physiol.39, 889-892.
Isogai, Y., Si, S., Pont-Lezica, L., Tan, T., Kapoor, V., Murthy, V. N. and Dulac, C.
(2011). Molecular organization of vomeronasal chemoreception. Nature478, U241-U130.
Jesuthasan, S. J. and Mathuru, A. S.(2008). The alarm response in zebrafish:
innate fear in a vertebrate genetic model.J. Neurogen.22, 211-228.
Jones, G. P., Milicich, M. J., Emslie, M. J. and Lunow, C.(1999). Self-recruitment
in a coral reef fish population.Nature402, 802-804.
Keefer, M. L. and Caudill, C. C.(2014). Homing and straying by anadromous
salmonids: a review of mechanisms and rates.Rev. Fish Biol. Fisheries 24, 333-368.
Kinoshita, M., Hosokawa, T., Urano, A. and Ito, E.(2004). Long-term potentiation
in the optic tectum of rainbow trout.Neurosci. Lett.370, 146-150.
Kobayashi, M., Sorensen, P. W. and Stacey, N. E. (2002). Hormonal and
pheromonal control of spawning behavior in the goldfish.Fish Physiol. Biochem. 26, 71-84.
Koide, T., Miyasaka, N., Morimoto, K., Asakawa, K., Urasaki, A., Kawakami, K.
and Yoshihara, Y.(2009). Olfactory neural circuitry for attraction to amino acids
revealed by transposon-mediated gene trap approach in zebrafish.Proc. Natl. Acad. Sci. USA106, 9884-9889.
Korsching, S.(2016). Aquatic olfaction. InChemosensory Transduction(ed. F.
Zufall and S. D. Munger), pp. 81-100. Amsterdam, Oxford, Bew York, Tokyo: Elsevier.
Kress, S., Biechl, D. and Wullimann, M. F.(2015). Combinatorial analysis of
calcium-binding proteins in larval and adult zebrafish primary olfactory system identifies differential olfactory bulb glomerular projection fields.Brain Struc. & Func.220, 1951-1970.
Leinders-Zufall, T., Brennan, P., Widmayer, P., Chandramani, P., Maul-Pavicic,
A., Jager, M., Li, X. H., Breer, H., Zufall, F. and Boehm, T.(2004). MHC class I
peptides as chemosensory signals in the vomeronasal organ.Science 306, 1033-1037.
Li, J., Mack, J. A., Souren, M., Yaksi, E., Higashijima, S., Mione, M., Fetcho, J. R.
and Friedrich, R. W.(2005). Early development of functional spatial maps in the
zebrafish olfactory bulb.J. Neurosci.25, 5784-5795.
Lipschitz, D. L. and Michel, W. C.(2002). Amino acid odorants stimulate microvillar
sensory neurons.Chem. Senses27, 277-286.
Lorenz, K.(1935). Der Kumpan in der Umwelt des Vogels.J. Ornithol.83, 137-213
and 289-413.
Mann, K. D., Turnell, E. R., Atema, J. and Gerlach, G.(2003). Kin recognition in
juvenile zebrafish (Danio rerio) based on olfactory cues.Biol. Bull.205, 224-225.
McBride, J. R., Fagerlund, U. H. M., Smith, M. and Tomlinson, N.(1964).
Olfactory perception in juvenile salmon: II. Conditioned respose of juvenile sockeye salmon (Oncorhynchus nerka) to lake waters.Can. J. Zool.42, 245-248.
Miklavc, P. and Valentincic, T.(2012). Chemotopy of amino acids on the olfactory
bulb predicts olfactory discrimination capabilities of zebrafishDanio rerio.Chem. Senses37, 65-75.
Miller-Sims, V. C., Atema, J., Gerlach, G. and Kingsford, M. J.(2011). How stable
are the reef odor preferences of settling reef fish larvae?Mar. Freshw. Behav. Physiol.44, 133-141.
Mirich, J. M., Illig, K. R. and Brunjes, P. C. (2004). Experience-dependent
activation of extracellular signal-related kinase (ERK) in the olfactory bulb.
J. Comp. Neurol.479, 234-241.
Moore, A. and Waring, C. P.(1996). Electrophysiological and endocrinological
evidence that F- series prostaglandins function as priming pheromones in mature male Atlantic salmon (Salmo salar) Parr.J. Exp. Biol.199, 2307-2316.
Mouritsen, H., Atema, J., Kingsford, M. J. and Gerlach, G.(2013). Sun compass
orientation helps coral reef fish larvae return to their natal reef.PLoS ONE8, e66039.
Nevitt, G. A., Dittman, A. H., Quinn, T. P. and Moody, W. J.(1994). Evidence for a
peripheral olfactory memory in imprinted salmon.Proc. Natl. Acad. Sci. USA91, 4288-4292.
Nordeng, H. (1971). Is local orientation of anadromous fishes determined by
pheromones?Nature233, 411-413.
Nordeng, H. and Bratland, P.(2006). Homing experiments with parr, smolt and
residents of anadromous Arctic charSalvelinus alpinusand brown troutSalmo trutta: transplantation between neighbouring river systems.Ecol. Freshw. Fish15, 488-499.
Ochs, C. L., Suntres, T., Zygowska, A., Pitcher, T. and Zielinski, B. S.(2017).
Organization of glomerular territories in the olfactory bulb of post-embryonic wild chinook salmonOncorhynchus tshawytscha.J. Morphol.278, 464-474.
Oka, Y., Saraiva, L. R. and Korsching, S. I.(2012). Crypt neurons express a single
V1R-related ora gene.Chem. Senses37, 219-227.
Olsén, K. H.(1989). Sibling recognition in juvenile Arctic charr,Salvelinus alpinus
(L.).J. Fish Biol.34, 571-581.
Olsen, J. B., Lewis, C. J., Kretschmer, E. J., Wilson, S. L. and Seeb, J. E.(2002).
Characterization of 14 tetranucleotide microsatellite loci derived from Pacific herring.Mol. Ecol. Notes2, 101-103.
Paris, C. B. and Cowen, R. K.(2004). Direct evidence of a biophysical retention
mechanism for coral reef fish larvae.Limnol. Ocean.49, 1964-1979.
Paris, C. B., Atema, J., Irisson, J.-O., Kingsford, M., Gerlach, G. and Guigand,
C. M.(2013). Reef odor: a wake up call for navigation in reef fish larvae.PLoS
ONE8, e72808.
Pelosi, P.(1994). Odorant-binding proteins.Critic. Rev. Biochem. Mol. Biol.29,
199-228.
Penn, D. J.(2002). The scent of genetic compatibility: sexual selection and the
major histocompatibility complex.Ethology108, 1-21.
Putman, N. F., Lohmann, K. J., Putman, E. M., Quinn, T. P., Klimley, A. P. and
Noakes, D. L. G.(2013). Evidence for geomagnetic imprinting as a homing
mechanism in Pacific salmon.Curr. Biol.23, 312-316.
Putman, N. F., Jenkins, E. S., Michielsens, C. G. J. and Noakes, D. L. G.(2014).
Geomagnetic imprinting predicts spatio-temporal variation in homing migration of pink and sockeye salmon.J. Roy. Soc. Interface11, 20140542.
Quinn, T. P. and Busack, C. A.(1985). Chemosensory recognition of siblings in
juvenile Coho salmon (Onchorinchus kisutch).Anim. Behav.33, 51-56.
Quinn, T. P. and Hara, T. J.(1986). Sibling recognition and olfactory sensitivity in
juvenile coho salmon (Oncorhynchus kisutch).Can. J. Zool.64, 921-925.
Quinn, T. P. and Tolson, G. M.(1986). Evidence of chemically mediated population
recognition in coho salmon (Oncorhynchus kisutch).Can. J. Zool.64, 84-87.
Journal
of
Experimental
Radford, C. A., Stanley, J. A., Tindle, C. T., Montgomery, J. C. and Jeffs, A. G.
(2010). Localised coastal habitats have distinct underwater sound signatures.
Mar. Ecol. Prog. Ser.401, 21-29.
Radford, C. A., Tindle, C. T., Montgomery, J. C. and Jeffs, A. G.(2011). Modelling
a reef as an extended sound source increases the predicted range at which reef noise may be heard by fish larvae.Mar. Ecol. Prog. Ser.438, 167-174.
Randlett, O., Wee, C. L., Naumann, E. A., Nnaemeka, O., Schoppik, D., Fitzgerald, J. E., Portugues, R., Lacoste, A. M. B., Riegler, C., Engert, F. et al.
(2015). Whole-brain activity mapping onto a zebrafish brain atlas.Nat. Methods 12, 1039-1046.
Robertson, D. R. (2001). Population maintenance among tropical reef fishes:
inferences from small-island endemics.Proc. Nat. Acad. Sci. USA98, 5667-5670.
Sandulescu, C. M., Teow, R. Y., Hale, M. E. and Zhang, C. B.(2011). Onset and
dynamic expression of S100 proteins in the olfactory organ and the lateral line system in zebrafish development.Brain Res.1383, 120-127.
Scholz, A. T., Cooper, J. C., Madison, D. M., Horrall, R. M., Hasler, A. D., Dizon,
A. E. and Poff, R. J.(1973). Olfactory imprinting in Coho salmon: behavioral and
electrophysiological evidence.Proc. 16th Conf. Great Lakes Res.1973, 143-153.
Scholz, A. T., Horrall, R. M., Cooper, J. C. and Hasler, A. D.(1976). Imprinting to
chemical cues: the basis for home stream selection in salmon.Science192, 1247-1249.
Selset, R. and Doving, K. B.(1980). Behaviour of mature anadromous char (Salmo
alpinusL.) towards odorants produced by smolts of their own population.Acta Physiol. Scand.108, 113-122.
Selwyn, J. D., Hogan, J. D., Downey-Wall, A. M., Gurski, L. M., Portnoy, D. S. and
Heath, D. D.(2016). Kin-aggregations explain chaotic genetic patchiness, a
commonly observed genetic pattern in a marine fish.PLoS ONE11, e0153381.
Singh, P. B.(2001). Chemosensation and genetic individuality.Reproduction121,
529-539.
Sorensen, P. W., Hara, T. J., Stacey, N. E. and Goetz, F. M. (1988). F
prostaglandins function as potent olfactory stimulants that comprise the postovulatory female sex pheromone in goldfish.Biol. Repro.39, 1039-1050.
Speca, D. J., Lin, D. M., Sorensen, P. W., Isacoff, E. Y., Ngai, J. and Dittman,
A. H.(1999). Functional identification of a goldfish odorant receptor.Neuron23,
487-498.
Speedie, N. and Gerlai, R.(2008). Alarm substance induced behavioral responses
in zebrafish (Danio rerio).Behav. Brain Res.188, 168-177.
Spehr, M., Kelliher, K. R., Li, X. H., Boehm, T., Leinders-Zufall, T. and Zufall, F.
(2006). Essential role of the main olfactory system in social recognition of major histocompatibility complex peptide ligands.J. Neurosci.26, 1961-1970.
Staaterman, E. and Paris, C. B.(2014). Modelling larval fish navigation: the way
forward.Ices J. Mar. Sci.71, 918-924.
Stacey, N. E., Sorensen, P. W., Van Der Kraak, G. J. and Dulka, J. G.(1989).
Direct evidence that 17 alpha ,20 beta -dihydroxy-4-pregnen-3-one functions as a
goldfish primer pheromone: Preovulatory release is closely associated with male endocrine responses.Gen. Comp. Endocrin.75, 62-70.
Stobutzki, I. and Bellwood, D. R.(1994). An analysis of sustained swimming
abilities of pre- and postsettlement coral reef fishes.J. Exp. Biol. Ecol.175, 275-286.
Stobutzki, I. and Bellwood, D. R.(1997). Sustained swimming abilities of the late
pelagic stages of coral reef fishes.Mar. Ecol. Prog. Ser.149, 35-41.
Swearer, S. E., Caselle, J. E., Lea, D. W. and Warner, R. R.(1999). Larval retention
and recruitment in an island population of a coral-reef fish.Nature402, 799-802.
Tang-Martinez, Z.(2001). The mechanisms of kin discrimination and the evolution
of kin recognition in vertebrates: A critical re-evaluation.Behav. Proc.53, 21-40.
Thorrold, S. R., Jones, G. P., Planes, S. and Hare, J. A.(2006). Transgenerational
marking of embryonic otoliths in marine fishes using barium stable isotopes.
Can. J. Fish. Aquat. Sci.63, 1193-1197.
Ueda, H., Nakamura, S., Nakamura, T., Inada, K., Okubo, T., Furukawa, N.,
Murakami, R., Tsuchida, S., Zohar, Y., Konno, K. et al.(2016). Involvement of
hormones in olfactory imprinting and homing in chum salmon.Sci. Rep.6, 21102.
Veyrac, A., Wang, G., Baum, M. J. and Bakker, J.(2011). The main and accessory
olfactory systems of female mice are activated differentially by dominant versus subordinate male urinary odors.Brain Res.1402, 20-29.
Weersing, K. and Toonen, R. J.(2009). Population genetics, larval dispersal, and
connectivity in marine systems.Mar. Ecol. Prog. Ser.393, 1-12.
Weltzien, F. A., Hoglund, E., Hamdani, E. H. and Doving, K. B.(2003). Does the
lateral bundle of the medial olfactory tract mediate reproductive behavior in male crucian carp?Chem. Senses28, 293-300.
Winberg, S. and Olsén, K. H.(1992). The influence of rearing conditions on the
sibling odor preference of juvenile arctic charr,Salvelinus alpinus L.Anim. Behav. 44, 157-164.
Wolanski, E., Doherty, P. and Carleton, J.(1997). Directional swimming of fish
larvae determines connectivity of fish populations on the Great Barrier Reef.
Naturwissenschaften84, 262-268.
Yabuki, Y., Koide, T., Miyasaka, N., Wakisaka, N., Masuda, M., Ohkura, M.,
Nakai, J., Tsuge, K., Tsuchiya, S., Sugimoto, Y. et al.(2016). Olfactory receptor
for prostaglandin F2alpha mediates male fish courtship behavior. Nature Neurosci.19, 897-904.
Yaksi, E., von Saint Paul, F., Niessing, J., Bundschuh, S. T. and Friedrich, R. W.
(2009). Transformation of odor representations in target areas of the olfactory bulb.Nature Neurosci.12, 474-482.
Yamamoto, Y., Hino, H. and Ueda, H.(2010). Olfactory imprinting of amino acids in
lacustrine sockeye salmon.PLoS ONE5, e8633.
Yoshihara, Y. (2009). Molecular genetic dissection of the zebrafish olfactory
system.Res. Probl. Cell Diff.47, 97-120.
Ziegler, A., Dohr, G. and Uchanska-Ziegler, B.(2002). Possible roles for products
of polymorphic MHC and linked olfactory receptor genes during selection processes in reproduction.Am. J. Repro. Immunol.48, 34-42.