Copyright 0 1994 by the Genetics Society of America
DNA Sequence Effects
on
Single Base Deletions Arising During DNA
Polymerization i n
Vitro
by
Escherichia coli
Klenow Fragment Polymerase
Fei
Jun Wang and
Lynn
S .
Ripley’
Department of Microbiology and Molecular Biology, UMD-New Jersey Medical School, Newark, New Jersey 07103
Manuscript received September 28, 1993 Accepted for publication November 29, 1993
ABSTRACT
Most single base deletions detected after DNA polymerization in vitro directed by either Escherichia coli DNA polymerase I or its Klenow fragment are opposite Pu in the template. The most frequent mutations were previously found to be associated with the consensus template context 5’-PyTPu-3’. In this study, the predictive power of the consensus sequence on single base deletion frequencies was directly tested by parallel comparison of mutations arising in four related DNAs differing by a single base. G, a deletion hotspot within the template context 5’-TTGA-3’, was substituted by each of the 3 other bases. Previous studies had shown that deletions opposite the G were frequent but that deletions opposite its neighboring A were never detected. Based on the predictions of the consensus, the substitution of T for G should produce frequent deletions opposite the neighboring A due to its new 5’-l”ITA-3’ template context. This prediction was fulfilled; no deletions of this A were detected in the other templates. The consensus further predicted that deletions opposite template C would be lower than those opposite either A or G at the same site and this prediction was also fulfilled. The C substitution also produced a new hotspot for 1 bp deletions 14 bp away. The new hotspot depends on quasi-palindromic misalignment of the newly synthesized DNA strand during polymerization; accurate, but ectopically templated synthesis is responsible for this mutagenesis. Mutations templated by quasi-palindromic misalignments have previously been rec- ognized when they produced complex sequence changes; here we show that this mechanism can produce frequent single base deletions. The unique stimulation of misalignment mutagenesis by the C substitution in the template is consistent with the singular ability of C at that site to contribute to extended comple- mentary pairing during the DNA misalignment that precedes mutagenesis.
I
N vitro studies of DNA polymerization fidelity pro- vide one approach to identifylng a subset of mecha- nisms that may contribute to spontaneous mutagenesis. The specificity of polymerization errors and their de- pendence upon experimentally manipulable factors such as DNA sequence and DNA polymerase provide important clues to their underlying causes. Moreover, demonstration of dependence (or independence) of mutagenesis i n v i v o and in vitro on the same manipu- lable factors may provide a basis for deciding which spontaneous mutations are caused directly by i n v i v opolymerization errors (RIPLEY 1990).
In vitro studies using a wide variety of DNA polym- erase enzymes and several different DNA templates have been used to generate mutants which have now been sequenced in substantial numbers (KUNKEL 1985a,b; de Boer and RIPLEY 1988; KUNKEL et al. 1989; Mo et al. 1991). The studies reveal that multiple mechanisms con- tribute to polymerization dependent frameshift muta- genesis in vitro and that the relative importance of these mechanisms to the spectra of mutants produced by dif- ferent polymerases varies widely.
’
To whom correspondence should be addressedGenetics 136: 709-719 (March, 1994)
Studies in this laboratory have focused on the frame- shifts produced by Escherichia coli DNA polymerase I
and its Klenow fragment while copying single-stranded templates designed to permit the specific recovery of
frameshifts in either the +1 or -1 directions (DE BOER and RIPLEY 1988; PAPANICOLAOU and RIPLEY 1989, 1991;
REVICH and RIPLEY 1990). In vitro studies of
polymerizationdependent frameshifts in the -1 direc- tion, led to the description of three distinguishable frameshift mechanisms. Two of the three mechanisms rely on misaligned DNA pairing. One misalignment mechanism implicates misalignment of directly re- peated DNA sequences. In vitro the sequence changes characteristic of this mechanism are multibase deletions deleting one copy of the repeat and bases between the repeats. This mechanism is like that proposed by STREIS-
INGER and ceworkers (1966) and has been implicated
repeatedly as a major contributor to spontaneous frame- shift mutagenesis i n v i v o (STEWART and SHERMAN 19974;
FARABAUGH et al. 1978; PRIBNOW et al. 1981; RIPLEY et al.
complex frameshifts (substitution/deletion/insertion changes) which are precisely predicted by templated, but misaligned DNA synthesis, followed by realignment of the DNA to the template. Similar frameshift se- quences attributable to these misalignment classes have been found in vivo and are implicated in spontaneous mutagenesis (RIPLEY 1982; DE BOER and RIPLEY 1984; HAMPSEY et al. 1988). Misalignment mutagenesis in its simplest form might be imagined to be primarily a func- tion of DNA sequence and to be largely insensitive to DNA polymerase. Experimentally, this appears not to be the case. Both in vitro and in vivo, misalignment mu- tagenesis specificity and frequency is strongly influenced by the DNA polymerase (RIPLEY and SHOEMAKER 1983; RIPLEY et al. 1983; DE BOER and RIPLEY 1984; KUNKEL 1985a,b; FIX et al. 1987; PAPANICOLAOU and RIPLEY 1989, 1991; BEBEBNEK et al. 1990; MASKER and CRISSEY 1993; SCHAAPER 1993).
In addition to misalignment-based frameshifts, E . coli
PolI and Klenow polymerase frequently produce 1-bp deletions at specific DNA contexts whose characteristics are inconsistent with frameshift mechanisms depending on misaligned pairing stabilized by either direct repeats
or quasi-palindromes (DE BOER and RIPLEY 1988). The deletions are most frequently seen opposite template purines, but not all template purines are prone to de- letions. Only template purines adjacent to 5’ Ts were frequently mutated. These mutations do not depend on repetition of the purine. The average deletion frequen- cies opposite template Pu’s which are part of a run of identical bases are not higher than frequencies opposite
Pu’s that are not part of runs (DE BOER and RIPLEY 1988). This shows that primer template misalignments, stabi- lized by the kind of slipped mispairing hypothesized by STREISINCER et al. (1966) and regularly invoked to ex- plain the loss of a base pair in a run of identical bases does not regularly stimulate this mutagenesis.
We initially proposed that the context might be error prone because the template sequence could assume a DNA configuration which would promote the ability of the polymerase to bypass the purine and move on to the adjacent
5‘
T template site. Evidence generally consis tent with this idea was suggested by studies of the speci- ficity of acridine-induced frameshift mutagenesis due to the presence of acridine during in vitro DNA polymer- ization and molecular modeling of sequence-specific, crystalline nucleic acid structures associated with acri- dines into the context of a replicating DNA molecule (REVICH and RIPLEY 1990; BERMAN et al. 1992). However, an alternative model has been proposed to explain 5’context effects on the basis of initiation of mutagenesis through misincorporation (KUNKEL 1990). In this model, the first step of mutagenesis is the incorporation of a nucleotide opposite the deletion site that is non- complementary to that site, but is instead complemen- tary to the 5’ nucleotide. Subsequent realignment of this
misincorporated base and its extension produce the frameshift.
Despite their substantial differences, the two models are difficult to distinguish experimentally. For example, it appears that in some cases slippage is likely to occur first. Slippage allowing incorporation of a base comple- mentary to the 5’ adjacent template base which is pos- tulated to subsequently “slip back” to the normal align- ment thereby accounting for higher frequencies of base substitutions complementary to the 5’ base at the pre- ceding site (KUNKEL and SONI 1988). The observation that slippage may precede apparent mispairing qualifies the generality of any experiment employing forced misincorporations to produce predicted frameshifts in support of the model that requires misincorporation first (BEBENEK and KUNKEL 1990). Experimentally forced misinsertion experiments clearly demonstrate that re- alignments between primer and template occur but cannot address the issue of whether misinsertion pre- cedes misalignment or whether instead misalignment precedes complementary incorporation. It is presently unclear whether at a particular site or group of sites, misincorporation or misalignment is the first step of mutagenesis.
Our previous studies derived a preferred DNA context for E . coli DNA PolI-induced 1-bp deletions by devel- oping a consensus sequence from the overall spectrum of frameshifts arising in approximately 120 bp of DNA. In this study, the direct manipulation of a single DNA base was used to test the predictive power of the con- sensus sequence on mutagenesis at one site. A specific 1-bp deletion context, 5‘ TTGA 3’ was selected for study. In this sequence deletions opposite G are frequent. This report describes the result of substituting the G with each of the other bases (A, T or C ) on the frequency and specificity of frameshifts arising during in vitro polymer- ization. The substitutions are variously predicted to dis- rupt an existing hotspot and/or create novel hotspots. This measurement examines polymerization errors o p posite four different bases embedded within an other- wise constant DNA context.
MATERIALS AND METHODS
Enzymes and chemicals: In vitro DNA synthesis was cata- lyzed by either E . coli DNA polymerase I-large fragment (Klenow) or its entire form (PoZI) , both purchased from Boeh- ringer Mannheim. T4 phage polynucleotide kinase was from Stratagene. [?’*P]ATP was from Amersham.
DNA constructions: The M74 phage is a derivative of M13- mp8, previously described by DE BOER and RIPLEY (1988). De- rivatives of M74 with base substitutions at position G,,, used in this study were constructed using oligonucleotidedirected mutagenesis (Amersham kit). The constructions are desig- nated as M74A,,,, M74C1,, and M74T1,,. The original M74 is analogously designated M74G,,. The four DNAs differ only at their 108 site.
Oligonucleotides: M13, S20 universal sequencing primer
DNA Contexts of Polymerase Errors 71 1
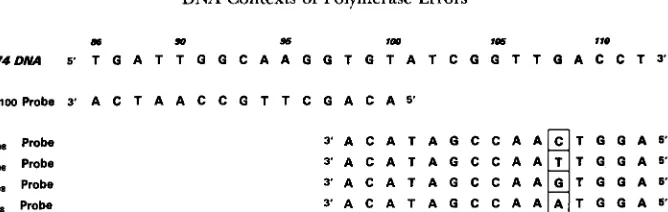
m 90 % IO0 106 I IO
M74DNA S T G A T T G G C A A G G T G T A T C G G T ~ Q A C C T ~
86-tWhobe 3' A C T A A C C G T T C G A C A 5'
G,,, Probe A,, Probe C,,, Probe TI,, Probe
FIGURE 1.-DNA sequence of the template in the vicinity of G,,, and oligonucleotide probes used in construction of new templates and analysis of mutants. The A,,,, C,,, and TI,, probes were used to construct templates containing these substitutions. These and the remaining probes illustrated were also used to identify the positions of mutants in this region in the DNA among blue plaques recovered after DNA polymerization and transfection into E . coli. The mutant genomes fail to bind the labeled oligo as tightly as the fully complementary wild type genome during stringent washing of nitrocellulose filters prepared by dot-blot.
for in vitro to prime DNA synthesis for mutagenesis experi- ments. The T at the 3' end of this primer is complementary to the A at position 128 in M74. DNA sequencing was primed by either S20 primer or S40 universal primer (5'-GTTT- TCCCAGTCACGAGS') from New England Biolabs. Five oli- gonucleotides for sequence specific construction and probing, were synthesized using a Applied Biosystems 380 Synthesizer. The oligos used for constructions and probing were each complementary to the 108 site, covering the 98-112 region. Their sequences are shown in the Figure 1.
In vitro DNA synthesis: The S20 primer and single- stranded M74 DNAs were annealed at a template concentra- tion of 50 nM in 50 mM of TrisHCl (pH 7.6) and 0.1 M NaC1. Primer was added in approximately 2-fold molar excess to tem- plate; the total volume of the annealing mixture was 60 pl. Annealing was accomplished by incubating the mixture at 65" for 5 min, followed by 15 min at 37". The annealed mixture was held on ice until use. Primer extensions were carried out by adjusting the annealed template-primer mixture to 10 nM in 50 mM TrisHC1 (pH 7.6), 10 mM MgCl,, 1 mM dithiothreitol and 250 p~ of each of the four dNTPs. A sample of 0.1-0.2 unit/pl DNA polymerase was added to start the synthesis; the total reaction volume was 300 pl. The extension reactions were incubated for 30 min at 37".
The extent of DNA synthesis was measured as a function of time. The newly synthesized DNA strand was labeled by in- corporation of ~'~P-labeled dATP. Samples from the reaction were stopped by the addition of EDTA, and were spotted onto DE81 Whatman disks. These disks were washed in 0.3 M am- monium formate and 10 mM of sodium pyrophosphate, dried and counted. Klenowdirected DNA polymerization reached the stationary phase after 30 min of extension, corresponding roughly to one round of extension, if all templates are used. In parallel, the extent of synthesis was examined on a 0.8% agarose gel. Samples taken at 10, 20 and 30 min were exam- ined. The agarose gel electrophoresis showed that, after 10 min of reaction, single-strand DNA disappeared and that double-strand nicked DNA appeared (data not shown).
Assay for in v i t r o mutagenesis: Competent cells were pre- pared using the CaC1, method (MANIATIS et al. 1982). 0.6 ml of E . coli JMlOl competent cells was transfected by adding an amount of DNAfrom the polymerization reaction correspond- ing to 0.04 pmol of template. The transfected cells, together with 0.2 ml log phase E . coli JMlOl cells, 20 pl of isopropyl Pmthiogalactopyranoside (IPTG) (100 mg/ml) 40 pl of X-gal (20 mg/ml in dimethylformamide) and 2.5 ml of soft agar (8 g of Bacto agar, 9 g of NaCl/liter) on one minimal medium plate (containing 30 ml of 0.2 g MgS04-7H,0, 2 g citric acid anhydride, 10 g K,HPO,, 3.5 g Na(NH4)HP0,.4H,0, 4 g glu- cose, 5 mg thiamine and 16 g Bacto agar/liter) . Mutants were collected after overnight incubation at 37", followed by an- other 24 hr of standing at room temperature to allow full blue
color to develop. There were approximately 0.5-1 X lo4
plaques/plate. Mutant DNA was isolated for sequencing from blue plaques purified by restreaking.
Mutant mapping by oligonucleotide probing A dot-blot as- say was used to quickly identify the mutants in the vicinity of the GI,, site, using a protocol similar to that used for T4 DNA
(RIPLEY et a l . 1988). Mutants between 86 and 112 in the DNA sequence were identified by probingwith two oligonucleotides one extending from 86 to 100 and another extending from 97 to 112. The oligo extending from 97 to 112 was appropriately matched to the identity of the base at 108 in the M74 template for the experiment (Figure 1). A 60-pl sample of phage stock (0.5-1 X 10") was deposited on a nitrocellulose filter using a dot-blot apparatus. DNA was released from phage by addition of 60 pl of 0.6 M NaOH. Filters were prehybridized and washed as previously described (RIPLEY et a l . 1988). Hybridization was at 37" overnight followed by a 10-min stringent wash at 48". DNA sequencing: All of the mutants identified by the proh ing assay as lying in the vicinity of the 108 site were sequenced. DNA preparation and sequencing were carried out as de- scribed previously (PAPANICOLAOU and RIPLEY 1989). The se- quencing primer was 5' labeled with "P using T4 polynucleo- tide kinase.
Polymerase pausing: In vitro pausing experiments were carried out as in DNA polymerization experiments used to gather mutants except that the primer was first 5'-''P-labeled using T4 polynucleotide kinase. Polymerization was carried out in a total reaction volume of 60 p1 from which aliquots were removed in a time course and the reaction stopped by directly pipetting into the gel loading solution containing formamide and dye. Samples were loaded on a denaturing polyacrylamide gel and run in parallel with a DNA sequencing reaction using the same primer. Pausing pattern was visually estimated from the radioautograms.
RESULTS AND DISCUSSION
M74, a derivative of M13-mp8, specifically designed for examining 1-bp deletions (DE BOER and RIPLEY 1988) was modified for these studies at the 108 site. The G at the 108 site [G,,,] was a site of frequent deletions in previous studies and is surrounded by the following con- text:
5'
TTGAC 3'. The G,,, base was substituted withA,,,, C,,, or T,,, using oligo-directed mutagenesis. Each
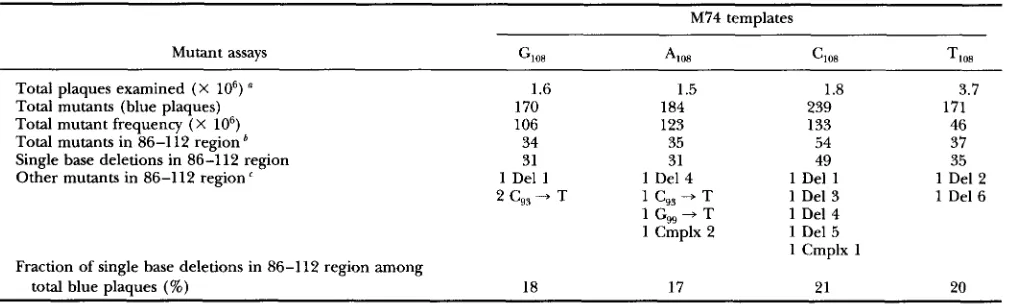
TABLE 1
Frameshifts produced by in vitro polymerization by E . coli Klenow DNA polymerase
M74 templates
Mutant assays GI,, 4 0 , C,,, TI,,
Total plaques examined ( X lo6) a 1.6 1.5 1.8 3.7
Total mutants (blue plaques) 170 184 239 171
Total mutant frequency ( X IO6) 106 123 133 46
Total mutants in 86-1 12 region 34 35 54 37
Single base deletions in 86-112 region 31 31 49 35
Other mutants in 86-112 region ' 1 Del 1 1 Del 4 1 Del 1 1 Del 2 2
c,,
+ T 1 C,, - j T 1 Del 3 1 Del 61 C,, + T 1 Del 4
1 Cmplx 2 1 Del 5 1 Cmplx 1
Fraction of single base deletions in 86-112 region among
total blue plaques (%) 18 17 21 20
a Average plaques per plate were 0.5-1 X lo4.
*
Determined by oligonucleotide probing.T h e sequences of Deletions (Del) and Complex frameshifts (Cmplx) are shown in Figure 3. Base pair changes are indicated by position number and sequence change in the template strand.
presence of IPTG and X-gal. This report describes the results of analysis of 764 blue plaques.
Mutations mapping to the vicinity of the 108 site were identified by the oligonucleotide probing assay de- scribed in the MATERIALS AND METHODS and Figure l. The mapping identified the mutants with altered DNA se- quences between bases 86 and 112 of "74. The validity of the mapping was initially established using approxi- mately 120 sequenced frameshifts (-30 mutants from each of the 4 templates) selected for their blue plaque phenotype. Application of the mapping protocol to the sequenced mutants produced neither false negatives nor false positives.
Each mutant identified by probing to lie between 86 and 112 in the sequence was sequenced. A summary of the data is presented in Table 1. Most of the sequenced frameshifts were single base deletions. The fraction of mutants mapping to the DNA segment 86-112 ranged from
17
to21%
in the various templates and is similar to the 16% found in our previous studies of the un- modified [G,,,] template (PAPANICOLAOU and RIPLEY 1989). The single base deletion spectrum for each tem- plate is presented in Figure2.
The sequences of addi- tional mutant types found are shown in Figure 3.Single base deletions: Single base deletion spectra i n
GI,, and A,,, templates: All spectra have a hotspot for 1-bp deletions opposite Gin the TTGG (89-92) context, consistent with its 5' TPu context. The consistency of mutations at this site in all spectra suggests that the spec- tra can be directly compared. Although the ratio of G,,, deletions to G,, deletions is somewhat smaller in all of these experiments than in previously reported measure- ments (PAPANICOLAOU and RIPLEY 1989; DE BOER and RIP- LEY 1988) the consistency in the independently meas- ured templates reported here argues that the reason does not lie in day-today variation in the protocol. E. coli DNA polymerase I produced a similar ratio of G,,,:G,,
deletions as does E. coli Klenow polymerase (data not shown). Comparison of the G,,, and A,,, spectra reveals no major differences (Figure
2).
The G,,, + A,,, sub- stitution has little, if any, influence on mutations at the 108 site or its immediate neighbors. This result supports the designation of TPu as a site of frequent deletion, and does not favor a specific purine.Single base deletion spectra i n T,,, and C,,, templates: In contrast to the purine substitution at the 108 site, pyrimidine substitutions produced spectra readily dis- tinguishable from the mutant spectrum in the G,,, tem- plate (Figure 2). In the T,,, spectrum, a novel TPu junc- tion is created at A,,,. Consistent with the association of frameshift hotspots with this sequence, a new hotspot is found at A,,. The A,,, deletion was not detected in any other template tested. The deletions at A,,, represents 20% of the mutants in the 86-112 region (or 4% of all mutants) sequenced in the M74T1,, template. This fre- quency of deletions at A,,, is comparable to the frequen- cies of 1-bp deletions at Glos or A,,, in the G,,, and
Alas
templates, respectively. This observation suggests that TPu context is sufficient to define a high frequency deletion site; the identity of the purine appears to be unimportant in this region. In addition, the similar frequency of A,,, or A,,, deletions in the A,,, or T,,, templates, respectively, demonstrates that repeats of two purines do not detectably stimulate deletions at this site.E. coli DNA polymerase I also produced A,,, deletions on the M74Tlo, template. After polymerization by PolI,
A,,, was found twice among 25 blue plaques sampled from M74T1,,, but was not found at all among the 64
DNA Contexts of Polymerase Errors 713
86 91 94 1 IM 111
M74-GlO, C T G A T T G G C A A G G T G T A T C G
...
G T T G A C C T G.
.
. .
.
86 91 94 108 I12
M 7 4 - A 1 0 B C T G A T T G G C A A G G T G T A T C G G T T A A C C T G
86 91 94 I08 I I ?
M 7 4 - T l O , C T G A T T G G C A A G G T G T A T C G G T T T A C C T G
66 91 94 108 112
M 7 4 - C l W C T G A T T G G C A A G G
..
T G T.
A T C G G T T C A C C T GFIGURE 2.Spectra of 1-bp deletions in templates G,,, A,,,, T,, and CIo8. The template strand is illustrated. Each of the four templates differs by only a single base located at position 108. Mutants were mapped to this region by oligonucleotide hybridization. Each mutant mapping to the region was sequenced. The position of each deletion mutant is indicated by a W. When a deleted base occurs in the midst of identical bases, DNA sequence does not identify which base has actually been deleted. In this figure the is consistently placed at the 5’ most position in the template where the deletion could have occurred.
sites over the more general PyPu designation, consistent with our earlier studies.
Deletions at C,,, are more rare than are A,,, or G,,,
deletions, consistent with the absence of a purine at the deletion site. The run of three consecutive Ts in TI,,, precludes assignment of a l-bp deletion frequency spe- cifically to T,,,. However, 1-bp deletions at T,, are prob- ably not higher than at C,,, because the sum of mutants at sites between 106 and 108 in both the ClO8 and T,,, templates is the same and the calculated frequency is
2-fold lower in the TI,, template (8/35 X 0.46 X
than in the C,,, template (8/49 X 1.33 X (see Table 1). This example further supports the conclusion of our earlier studies (DE BOER and RIPLEY 1988) that modest monotonic base run are not hotspots for l-bp deletions during in vitro DNA polymerization by Kle- now fragment DNA polymerase.
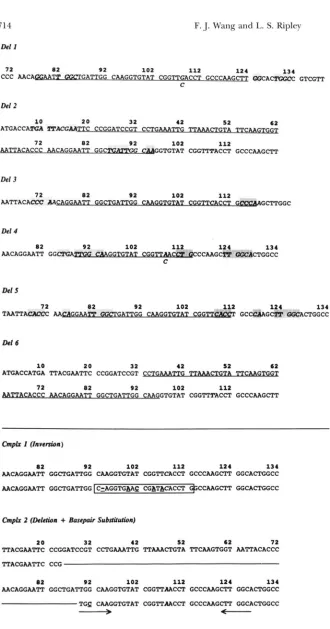
Del I
Del 2
1 0 3 2 2 0 4 2
5 2 6 2
ATGACCAlCA ll’ACGMm CCGGATCCGT C C T G A M T K m T A TTC-
72 1 1 2 82 1 0 2 9 2
ccc m c m m G G f l ( r ; a n u ; c CMGGTGTAT CGGTl’TACCT GCCCMGCIT
Del 3
7 2
AATTACACCC AAT-T G G mGCTTGGC
8 2 9 2
1 0 2
1 1 2 L.~.,
Del 4
8 2
AACAGGAATT G G C ! l W i P
9 2
1 0 2 L12”v 1 2 4 1 3 4
C
C C A A G C G C C
Del6
10 3 2 2 0
4 2 so 6 2
ATGACCATGA TTACGAATTC CCGGATCCGT
7 2 8 2
9 2 102 112
GG C a G T G T A T CGGTTTACCT GCCCAAGCTT
8 2 4 9 2 1 2 1 1 2 1 0 2 1 3 4
M C A G G M T T GGCTGATTGG CMGGTGTAT CCGTTCACCT GCCCAAGCTT GGCACTGGCC
FIGURE 3.-DNA sequences of multi- base deletions and complex frameshifts. The DNA bases deleted are indicated by underlining. The identity of the base at site 108 is indicated; in instances where the mutation was recovered in multiple templates, alternative bases are indi- cated at the 108 site. Directly repeated sequences in the vicinity of the deletion end points are indicated by shaded boxes. As described in the text, the re- peats can only explain the exact se- quences in the case of deletion (Del) 1 and 2. Complex mutant sequences are illustrated directly below the starting se- quence. Complex frameshift 1 (Cmplx 1) is an inversion of the sequence shown in the block. The result is the deletion of
A,,,
the 4 base substitutions T,, --j A, TIn2+ C, GI,, + A, and TIn7 + A. A mo- lecular explanation for this sequence change is presented in Figure 6. Com- plex mutation 2 is a base substitution Gq2
+ C (underlined) and bases lost indi-
cated by a line. The deletion is not ex- plained by misaligned complementary pairing. The base change could be pro- duced by a fold-back misalignment me- diated by the quasi-palindrome indi- cated by arrows.
2 0 3 2 42 s2 62 72
TTACGMTTC CCGGATCCGT C C T G A M T K TTMACTGTA TTCAAGTGGT AATTACACCC
TTACGAATTC CCG
8 2 9 2
1 0 2 1 1 2
1 2 4
1 3 4
AACAGGAATT GGCTGATTGG CAAGGTGTAT CGGTTAACCT GCCCAAGCTT GGCACPCCCC
-
TGc
CMGGTGTAT CGGTTMCCT GCCCAAGCTl’ GGCACTGGCC-
Despite the distance, the hotspot clearly depends on the C,,, context because the deletion is infrequent in each of the spectra from other templates. The new hotspot is accounted for by a quasi-palindromic DNA misalign- ment whose frequency is influenced by the ClOs site. Fig- ure 4 demonstrates how a quasi-palindromic fold-back misalignment explains this specific single base deletion. The explanation accounts for both the precise sequence
of the mutant and accounts for the specific association of the hotspot with the
M74-C,o,
template. Only the C,,, template increases the complementary pairing partici- pating in the DNA misalignment that precedes muta- genesis.DNA Contexts of Polymerase Errors 715
A
W % 1 w 108 I 14
S ' T G G C A A G G T G T A T C G G T T C A C C T G C C C A 3 G G G T 5'
mer 4+ o*
axtension
B
mo % 1 w 108
S ' T G G C A A G G T G T A T C G G T T C A C C T G C C C A 3 T C C A C A T A G C C A A G T G G A C G G G T "
1 I4
Iw
FoM-back
C
lo % 1 w 1 0 8
S ' T G G C A A G G T G T A T C G G T T K A C C T G C C C A 3
114
D
mo ss 100 1 0 8
W T G G C A A G G T G T A T C G G T T C A C C T G C C C A 3 , C A A G T G G A C G G G T s
I14
FIGURE 4.-A quasi-palindromic fold- back mechanism accounts for the
h4
deletion hotspot in the C,o, template.A shows the DNA template being cop- ied during a primer extension reaction mediated by E . coli Klenow DNA po- lymerase. In B, extension continues to position 95 where misalignment is ini- tiated. C shows the misaligned interme- diate. The boxed base pair can form only in the C,,, template; in all other templates this site is a mismatch. The misaligned DNA serves as a substrate for further DNA polymerization in D. Realignment of the ectopically synthe- sized DNA to the original template in E
creates the mutational heterozygote with a copy of the
h4
deletion in the newly copied strand. An additional round of replication creates the full mutation.E
W ns 100 108 114
S ' T G G C A A G G T G T A T C G G T T C A C C T G C C C A 3 , C C G - T C C A C A T A G C C A A G T G G A C G G G T S
D&thw~ a t A,
reactions. We had predicted that this mechanism occa- sionally produced 1-bp deletions as well, based on the observation that a single specific 1-bp deletion site [G15]
in M74 occurred at different frequencies during poly- merization by E . coli DNA polymerase I or its Klenow fragment. This hypothesis was based on the synthesis of two observations: (1) the mutant sequence was consis tent with a local quasi-palindromic misalignment mechanism and
(2)
mutations depending on quasi- palindromic misalignment often display different fre- quencies in comparisons of the two enzymes while single base deletions usually do not (PAPANICOLAOU and RIPLEY 1989,1991). The result reported here shows clearly that quasi-palindromic misalignments produce context spe- cific 1-bp deletions at substantial frequencies during in vitro polymerization reactions.We previously showed that the site-specific frequency of frameshift mutagenesis mediated by palindromic
DNA misalignments is correlated to the frequency of
DNA polymerase pausing (PAPANICOLAOU and RIPLEY
1991). Thus, we examined the influence of the template sequence changes at the 108 site on DNA polymerase pausing.
As
can be seen in Figure 5, the DNA sequence changes at 108 produced no detectible change in paus- ing under the conditions of the mutagenesis experi- ments. Thus, the mechanism by which C,,, promotes misalignment does not appear to include an influence of C,,, on DNA polymerase processivity and is thus prob- ably due to the additional complementary pairing dur- ing misalignment provided by the C,,, base change.Other mutant sequences in the 86-112 region: A
small number of DNA sequence changes were found in the 86-1 12 region that were not 1-bp deletions. The full repertoire of changes previously associated with in vitro DNA polymerization errors were found; namely, base pair substitutions, multibase deletions and complex frameshifts.
M74-C108 M74-Cl08
T C G A lV2CW451'1'30'2'5'
b 3
"
30
M74-U 08 M74-Gl08 M74-Al08 M74-Tl08
FIGURE 5.-DNA polymerase
pausing on GI,, A,,, C,,, and TI,, templates. Transient pausing of DNA polymerase during extension reactions similar to those used to
create mutations was measured. No influence of the identity of the base at the 108 site is seen on polymerase pausing in the DNA regions in which mutagenesis is measured. Panel A shows the C,, template. Panel B shows pausing on the re- maining templates. Dideoxy DNA sequencing patterns were used to
mark the positions of pausing in both panels. The M74C108 tem- plate was used in both cases and sy"-
thesis was directed by the same la- beled primer as used in the pausing experiment.
GIOA template and once in the AI,, template. The s u b
stitution creates an in-frame termination codon, and we believe that out-of-frame translational reinitiation ac- counts for the blue plaque phenotype (PAPANICOLAOU and RIPLEY 1989). A G% + T transversion was also found
to produce a light blue phenotype in the A,, template. This change does not produce a termination codon, and the molecular basis of its phenotype is unknown. Simi- larly, an adjacent transversion Tg8 + A was found to produce a light blue phenotype in the G,,, template (PAPANICOLAOU and RIPLEY 1989).
Deletions: Deletions between direct repeats are com- mon after E. coli DNA polymerase I extensions, but have been found more rarely after Klenow extensions. Six deletion sequences that include at least part of the 86-
112
region were sequenced in this study and are shown in Figure 3. Some deletions have end points associated with direct repeats and can thus be explained by primer misalignments; for example, deletion 1, found after r e p lication on both the G and the C templates. Deletion 2 can also be explained by this model, but the deleted sequence suggests that the misalignment from the u pstream sequence to the downstream position, the direc- tion opposite that predicted by primer misalignment during in vitro polymerization, but fully consistent with misalignments arising i n v i v o . Deletions 3-5 are impre- cisely associated with imperfect direct repeats and de- letion 6 is imprecisely connected to only a 2-bp repeat. Deletions with these characteristics have been quite fre- quently seen in studies of deletion mutagenesis in mam- malian cells, but have been rarely seen in procaryotes (MEUTH 1990). The low numbers of these mutations still provides no real insight into how they arise. Their re-
covery in these experiments was completely dependent on Klenow fragment rather than PolI polymerase syn-
thesis. Deletions recovered after PolI synthesis on these templates (data not shown) had endpoints associated with direct repeats that could account for their se- quences and were like those we have previously reported
(PAPANICOLAOU and RIPLEY 1989).
Complex frameshifts: Two complex mutations were found in the probed region. The sequences are shown in Figure
3.
Mutant 1, arose in theM74-ClO,
template and is an interesting variation on theb4
hotspot dele- tions associated with theM74-C,,,
template. In the com- plex 1 (Cmplx 1) mutant theb4
deletion is accompa- nied by four non-adjacent base pair substitutions, covering a distance of 8 bp. These substitutions reflect an inversion of the entire local sequence in which they lie. Theb4
deletion and the inversion are explained asa single mutational event involving templated DNA syn-
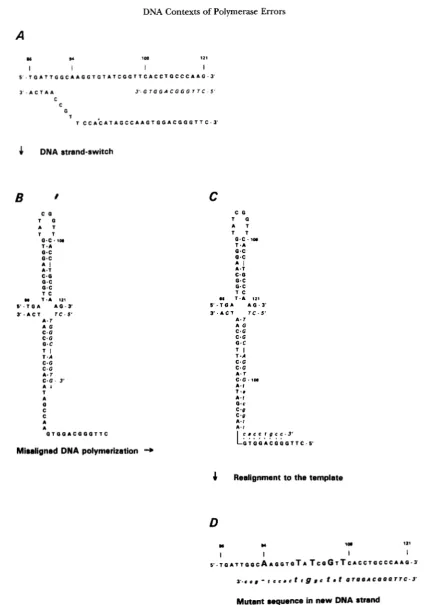
thesis initiated by a quasi-palindromic DNA misalign- ment. The sequence change cannot be accounted for by the fold-back misalignment described in Figure 4, but is instead explained by a strand-switch misalignment shown in Figure 6.
DNA Contexts of Polymerase Errors
A
717
8s )I 108 121
I I I I
~ ' - T 0 A T T G 0 C A A G G T G T A l C G G l l C A C C ~ 0 C C C A ~ G ~ ~ '
3 ' - A C T A A
3 ' . G T G G A C G G G T T C 5 '
C C
G
B
I.. .
A G C . G C . G
O . C
:
.!A b T A 0 C C A A
0 T 0 0 A C 0 0 0 T T C
Misaligned DNA polymerization -+
4
Realignment to the templatem u 1- 111
I I I I
S . T ~ A T T G ~ C A A Q G T G T A T c o G ~ T c ~ c c ~ ~ c c c ~ ~ o - 3
~ . c c r - t c c ~ c t t g s c 1 . r G Z Q Q A C Q G G T T C - 3
Mutant sequence in new DNA strand
FIGURE 6.-Inversion mutagenesis mediated by polymerase strand-switching. A shows the starting template sequence containing C , , which has been copied around the entire genome of " 7 4 and is now polymerization continues via strand displacement. The displaced strand is shown in bold, the newest synthesis is shown in italics. B describes a strand-switch misalignment that brings DNA initially extended to the 108 position by correctly aligned polymerization intojwtaposition with the template that will account for the mutant sequence. C shows the ectopic DNA polymerization step that produces the mutant DNA sequence. Ectopically templated DNA is indicated in lower case. Realignment of this DNA to the template in D places the mutant sequence in its final context. A further round of replication of this mutational heterozygote (not shown) produces a complete mutant. The deletion
CONCLUSIONS
The consensus DNA context for frequent 1-bp dele- tions produced during in vitro polymerization by E . coli
PolI Klenow polymerase successfully predicted the ef- fects of changing a base at a single deletion hotspot site to each of the other three bases. Most impressively, the frequency of deletions opposite template A in the con- texts, 5’-T1TA-3’ and 5’-TTAA-5’ was similar to that ob- served at G in the original 5’-TTGA-3’ context, but de- letions opposite template C in the 5’-TTCA-3’ context were reduced. Thus, the TPujunction sequence was con- sistently associated with high 1-bp deletion frequencies. Neither the identity of the purine (A or G) nor the theo- retical opportunity for slippage in the 5’-TTAA-3’ con- text influenced the deletion frequency. The important contribution of the 5’ T on mutant frequency in the TPu context was strongly confirmed by the absence of AIo9 deletions in the 5’-TTCA-3’ and 5’-TTGA-3’ contexts.
The large increase in single base deletions occurring at
b4
associated with the C,,, template is well explained by misaligned synthesis mediated by a quasi-palindromic DNA misalignment. The increased mutation frequency may be ascribed to extended complementarity to me- diate misaligned pairing in the C,,, template, a feature expected to increase the concentration of misaligned DNA intermediates and/or to increase the ability of the polymerase to extend them. It is reasonable to propose that the low frequencies ofb4
deletions in the other templates are due to less efficient formation or process- ing of the same misalignment. Indeed, quasi- palindromic misalignments should be more seriously considered as potential explanations for 1-bp deletions in contexts that are not TPu. For example, the loss of one base, opposite G96-9, [5‘-AGGTG3‘] might depend on a fold-back misalignment to the complementary DNA sequence opposite positions 127-130 [5’-CACT- 3’1. Such distant relationships should be readily uncov- ered using templates bearing base substitutions in the suspected template regions.Palindromic misalignments have been previously demonstrated to produce complicated DNA sequence changes during polymerization reactions mediated by E.
coli Klenow fragment (PAPANICOLAOU and RIPLEY 1989) and Yeast PolI (KUNKEL et al. 1989). Here the same class of misalignments is shown to produce 1-bp deletions. It should be further noted that quasi-palindromic mis- alignments predict context-specific base pair substitu- tions (RIPLEY 1982) and these substitutions may also con- tribute to mutational spectra.
It has been recently noticed, that during polymeriza- tion on damaged DNA templates, the Klenow polymer- ase produces specific double mutations whose frequen- cies are far higher than would be predicted on the basis of the occurrence of independent single mutations
(D. FEIC and L. LOEB, personal communication). In some cases, the sequences of these double mutants are consistent with the idea that an initial error during po-
lymerization produces a new strand bearing a change which would have an improved ability to form a fold- back misalignment that would template the second change. The substantial increase in single base deletions due to a single base change at C,,, observed in this study suggests that the frequency of secondary misalignment could be substantially enhanced by a single base change. Moreover, potentially large contributions by misalign- ment-mediated mutagenesis mechanisms may be ex- pected during in vitro polymerization on damaged tem- plates in view of the fact that DNA damage is expected to interfere with processive DNA polymerization and the demonstration that DNA polymerase pause sites corre- late strongly to the most frequent misalignment muta- tions (PAPANICOLAOU and RIPLEY 1991).
Altogether this study has refined our view of the mechanisms contributing to 1-bp deletions during po- lymerization by the E . coli Klenow polymerase. One mechanism is clearly associated with TPu template con- texts and fails to operate at a detectable level in a CPu context at the same site. Although these studies cannot
p e r se address whether this context preference reflects misincorporation and/or misalignment properties of Klenow DNA polymerase at this site, it provides the con- textual rules that must be explained by context-specific polymerase behavior. A broad exploration of DNA se- quence effects on Klenow fragment-mediated misincor- poration and mismatch extension reactions by JOYCE
et al. (1992) demonstrated large effects of context on polymerization behavior. For example, Klenow polym- erase directed highly variable, site-specific patterns of misinsertions at template G sites, but usually directed the rapid misinsertion of A at template A sites. Perhaps parallel measurements of the influence of context on both the biochemical and mutational characteristics of polymerase activity could be used to define the site- specific polymerization events associated with
polymerase-mediated single base deletions.
Some of the templates used in the work reported were constructed
in our laboratory by ZUWEI Q~AN and WEI LI. We thank CATHERINE PA-
PANICOLAOU for her advice. Support for this work is provided by grant
CN-50 from the American Cancer Society and by grant MCB-9305475
from the National Science Foundation.
LITERATURE CITED
BEBEBNEK, R, and T. A. KUNKEL, 1990 Frameshift errors initiated by
nucleotide misincorporation. Proc. Natl. Acad. Sci. USA 87:
BEBEBNEK, R , C. M. JOYCE, M. P. FITZCERALD and T. A. KUNKEL,
1990 The fidelity of DNA synthesis catalyzed by derivatives of
Escherichia coli DNA polymerase I. J. Biol. Chem. 265: 13878- 13887.
BERMAN, H. M., J. L. SUSSMAN, L. JOSHUA-TOR, G. G. REWCH and L.
s.
RIPLEY, 1992 A structural model for sequencespecific proflavin-
DNA interactions during in vitro frameshift mutagenesis.J. B i e
mol. Struct. Dyn. 10: 317-331.
DE BOER, J. G., and L. S. RIPLEY, 1984 Demonstration of the produc-
dromic DNA sequences. Proc. Natl. Acad. Sci. USA 81: 5528- tion of frameshift and basesubstitution mutations by quasipdin-
5531.
DNA Contexts of Polymerase Errors 719
mutations: hotspots for deletions of 1 bp by Klenow-fragment po- lymerase share a consensus sequence. Genetics 118: 181-191. FARABAUGH, P. J., U. SCHMEISSNER, M. HOFER and J. H. MILLER, 1978 Ge-
netic studies of the lac repressor. VII. On the molecular nature of spontaneous hotspots in the lacI gene of Escherichia coli. J. Mol. Biol. 126 847-863.
FIX, D. F., P. A. BURNS and B. W. GLICKMAN, 1987 DNA sequence analysis of spontaneous mutation in a p o l A l strain of Escherichia coli indicates sequence-specific effects. Mol. Gen. Genet. 202:
HAMPSEY, D. M., J. F. ERNST, J. W. STEWA~T and F. SHERMAN, 1988 Mul- tiple basepair mutations in yeast. J. Mol. Biol. 201: 471-486. JOYCE, C. M., X. C. SUN and N. D. F. GRINDLEY, 1992 Reactions at the
polymerase active site that contribute to the fidelity ofEscherichia coli DNA polymerase I (Klenow fragment). J. Biol. Chem. 267:
14485-24500.
KUNKEL, T. A,, 1985a The mutational specificity of DNApolymerase-P during in vitro DNA synthesis. Production of frameshift, base substitution and deletion mutations. J. Biol. Chem. 2 6 0 5787- 5796.
KUNKEL, T. A., 1985b The mutational specificity of DNA polymer- ases-a and -y during in vitro DNA synthesis. J. Biol. Chem. 260:
12866-12874.
KUNKEL, T. A,, 1990 Misalignment-mediated DNA synthesis errors. Biochemistry 2 9 8003- 8011.
KUNKEL, T. A., and A. SONI, 1988 Mutagenesis by transient misalign- ment. J. Biol. Chem. 263: 4450-4459.
KUNKEL, T. A,, R. K. tIAMATAKE, J. Mono-Fox, M. P. FITZGERALD and A. SUGINO, 1989 Fidelity of DNA polymerase I and DNA polymerase I-DNAprimase complex from Saccharomyces cerevisiae. Mol. Cell. Biol. 9: 4447-4458.
MANIATIS, T., E. F. FRITXH and J. SAMBROOK, 1982 Molecular Cloning: A Laboratory Manual. Cold Spring Harbor Laboratory, Cold Spring Harbor, N.Y.
MASKER, W., and M. A. CRISSEY, 1993 The effect of the 3’+ 5’ exo- nuclease of T7 DNA polymerase on frameshifts and deletions. Mutat. Res. 301: 235-41.
MEUTH, M., 1990 The structure of mutation in mammalian cells. Bio- chim. Biophys. Acta 1032: 1-17.
Mo, J. Y., H. MAKl and M. SEKIGUCHI, 1991 Mutational specificity of the dnaE173 mutator associated with a defect in the catalytic s u b
unit of DNA polymerase 111 of Escherichia coli. J. Mol. Biol. 222:
PAPANICOLAOU, C., and L. S. RIPLEY, 1989 Polymerase-specific differ- ences in the DNA intermediates of frameshift mutagenesis. In 267-272.
925-936.
vitro synthesis errors if Escherichia coli DNA polymerase I and its large fragment derivative. J. Mol. Biol. 207: 335-353. PAPANICOLAOU, C., and L. S. RIPLEY, 1991 An in vitro approach to
identifymg specificity determinants of mutagenesis mediated by DNA misalignments. J. Mol. Biol. 221: 805-821.
PRIBNOW, D., D. C. SIGURDSON, L. GOLD, B. S. SINGER, C. NAPOLI et al., 1981 rIIcistrons of bacteriophage T 4 DNA sequence surround the intercistronic divide and positions of genetic landmarks. J. Mol. Biol. 149: 337-376.
REVICH, G. G., and L. S. RIPLEY, 1990 Effects of proflavin and pho- toactivated proflavin on the template function of single-stranded DNA. J. Mol. Biol. 211: 63-74.
RIPLEY, L. S., 1982 Model for the participation of quasipalindromic DNAsequences in frameshift mutation. Proc. Natl. Acad. Sci. USA
RIPLEY, L. S., 1990 Frameshift mutation: determinants of specificity. Annu. Rev. Genet. 2 4 189-213.
RIPWY, L. S., and N. B. SHOEMAKER, 1983 A major role for bacterio- phage T4 DNA polymerase in frameshift mutagenesis. Genetics
1 0 3 353-366.
RIPWY, L. S., B. W. GLICKMAN and N. B. SHOEMAKER, 1983 Mutator
versus antimutator activity of a T4 DNA polymerase mutant dis- tinguishes two different frameshifting mechanisms. Mol. Gen.
Genet. 189: 113-117.
RIPLEY, L. S., A. CLARK and J. G. DEBOER, 1986 Spectrum of sponta- neous frameshift mutations. Sequences of bacteriophage T4 rII
gene frameshifts. J. Mol. Biol. 191: 601-613.
RIPLEY, L. S., J. S. DUBINS, J. G. DEBOER, D. M. DEMARINI, A. M. BOCERLI
et al., 1988 Hotspot sites for acridine-induced frameshift mu- tations in bacteriophage T4 correspond to sites of action of the T4 type I1 topoisomerase. J. Mol. Biol. 2 0 0 665-680. SCHAAPER, R., 1993 The mutational specificity of two Escherichia coli
dnaE antimutator alleles as determined from laczmutation spec- tra. Genetics 134: 1031-1038.
STEWART, J. W., and F. SHERMAN, 1974 Yeast frameshift mutations iden- tified by sequence changes in iso-lcytochrome c, pp. 102-127 in
Molecular and Environmental Aspects of Mutagenesis, edited by L. PRAKASH, F. SHERMAN, M. W. MILLER, C. W. LAWRENCE and H. W. TABER. Charles C Thomas, Springfteld, Ill.
STREISINGER, G., Y. OWA, J. EMRICH, J. NEWTON, A. TSUGITA et al., 1966 Frameshift mutations and the genetic code. Cold Spring Harbor Symp. Quant. Biol. 31: 77-84.
79: 4128-4132.