rspb.royalsocietypublishing.org
Research
Cite this article:
Martin CH, Crawford JE,
Turner BJ, Simons LH. 2016 Diabolical survival
in Death Valley: recent pupfish colonization,
gene flow and genetic assimilation in the
smallest species range on earth.
Proc. R. Soc. B
283
: 20152334.
http://dx.doi.org/10.1098/rspb.2015.2334
Received: 29 September 2015
Accepted: 5 January 2016
Subject Areas:
evolution, genetics, genomics
Keywords:
demographic inference, population genomics,
speciation, conservation genetics, genetic
accommodation, introgression
Author for correspondence:
Christopher H. Martin
e-mail: [email protected]
Electronic supplementary material is available
at http://dx.doi.org/10.1098/rspb.2015.2334 or
via http://rspb.royalsocietypublishing.org.
Diabolical survival in Death Valley: recent
pupfish colonization, gene flow and
genetic assimilation in the smallest
species range on earth
Christopher H. Martin
1, Jacob E. Crawford
2,3, Bruce J. Turner
4and Lee H. Simons
51Department of Biology, University of North Carolina, Chapel Hill, NC, USA
2Department of Integrative Biology, and3Center for Theoretical Evolutionary Genomics, University of California,
Berkeley, CA, USA
4Department of Biological Sciences, Virginia Tech, VA, USA 5US Fish and and Wildlife Service, Las Vegas, NV, USA
One of the most endangered vertebrates, the Devils Hole pupfishCyprinodon diabolis, survives in a nearly impossible environment: a narrow subterranean fissure in the hottest desert on earth, Death Valley. This species became a conservation icon after a landmark 1976 US Supreme Court case affirming federal groundwater rights to its unique habitat. However, one outstanding question about this species remains unresolved: how long hasdiabolis per-sisted in this hellish environment? We used next-generation sequencing of over 13 000 loci to infer the demographic history of pupfishes in Death Valley. Instead of relicts isolated 2– 3 Myr ago throughout repeated flooding of the entire region by inland seas as currently believed, we present evidence for frequent gene flow among Death Valley pupfish species and divergence after the most recent flooding 13 kyr ago. We estimate that Devils Hole was colonized by pupfish between 105 and 830 years ago, followed by genetic assimilation of pelvic fin loss and recent gene flow into neighbouring spring systems. Our results provide a new perspective on an iconic endan-gered species using the latest population genomic methods and support an emerging consensus that timescales for speciation are overestimated in many groups of rapidly evolving species.
1. Introduction
Simple environments, isolated habitats with minimal biotic interactions, serve as natural laboratories for examining the extremes of ecological and evolutionary processes [1,2]. From adaptive radiation on remote depauperate islands [3–5] to laboratory microcosms [6–8], simple ecosystems may be isolated from com-plex community-level processes and often provide unique opportunities for testing theory. For example, what are the simplest environments in which specia-tion can occur without physical barriers [9–12]? How does effective populaspecia-tion size affect the risk of extinction or the potential for adaptation [13–16]? Small inbred populations in simple natural environments may also provide a window into the future of managed endangered populations [17–19].
Devils Hole is the smallest natural range of any vertebrate species, comprising a 3.522 m opening into a flooded fault cavern greater than 130 m deep (figure 1d; [21,22]). The census population size of Cyprinodon diabolis within Devils Hole has varied from only 35–548 individuals since semi-annual monitor-ing began in 1972 [23], reachmonitor-ing its record low in 2013 and second lowest point (38) in 2006–2007. In addition to limited spawning habitat (a single 2.66.1 m shelf [22]: figure 1d), the population is limited by the productivity of Devils Hole, which receives no direct sunlight for two months each year and produces 200 times less algal biomass than neighbouring springs [24]. The population
size typically decreases in response to reduced winter food availability and rebounds in the spring. Year-round water temperatures in Devils Hole remain near 328C [21]; however, broad thermal tolerances in Cyprinodon from 21.98C to 45.58C [25–27], such asCyprinodon pachycephaluswhich com-pletes its entire life cycle at 448C [28], suggest that diabolis
should tolerate higher water temperatures (contra [29]). The improbability of a vertebrate species surviving at such low population sizes in a suboptimal habitat, even for a short time, would seem to defy basic principles of conservation biology. For example, the probability of diabolis extinction within 50 years exceeds 80% based on historical census population sizes [23]. Initial time-calibrated mitochondrial phylogenies suggested that Death Valley pupfishes were relicts isolated for 2–3 Myr [30–32] and estimated the age of the
diabolismitochondrial lineage to be 500 000 years [30,31]. How-ever, early studies did not detect any unique mitochondrial haplotypes indiabolisrelative to other Death Valley pupfishes [33], consistent with more recent colonization. Geological evi-dence strongly indicates that Devils Hole opened to the surface only 60 000 years ago based on the abrupt cessation
of mammillary calcite deposition [34], which places an upper bound on the isolation ofdiaboliswithin this extreme habitat. Previous studies attributed overestimation of species diver-gence times to incomplete lineage sorting [30,31]; however, this cannot account for such a large discrepancy [35,36]. Instead, this conflict appears to be caused by 11 Myr priors placed on the root age ofCyprinodonþMegupsilonþCualac
[30,31] based on the assumption that pupfish populations were isolated by the formation of ancient land barriers [32]. Thus, the discrepancy between geological and mtDNA evi-dence for the age of diabolis is not due to any underlying conflict in the data (e.g. time-calibration is extremely sensitive to priors [37]), but rather the assumption that Cyprinodon
cannot disperse over land. Devils Hole was never connected to neighbouring springs by surface flow, thus the current con-sensus is thatdiaboliscolonized approximately 10–20 000 years ago when water levels were higher [22,31].
A recent study estimated the age ofdiabolisfrom the coalesc-ence of 12 microsatellite loci using approximate Bayesian computation without assumptions about overland dispersal and placed the age between 90 and 14 400 years (95% credible
(a) (b)
(c)
(d)
Figure 1.
(a) Sample sites within Death Valley National Park, California and Ash Meadows National Wildlife Refuge, Nevada overlaid on the surrounding topography
(Google Maps), extant desert springs (dark blue lines) and approximate Pleistocene waterways (light blue shading) present in the region 10 – 35 kyr ago [20].
Colours of each desert spring correspond to population colours in figure 2. (b) Aerial view of Ash Meadows (Google Earth) with the position of Devils Hole indicated
in red. (c,d) Close-up views of Devils Hole in November 2013. The white pipe (for daily supplemental feeding which began in 2007) marks the edge of the spawning
shelf. (Online version in colour.)
rspb.r
oy
alsocietypublishing.org
Pr
oc.
R.
Soc.
B
283
:
20152334
intervals in table 1: [38]). However, additional uncertainty in microsatellite mutation rates (1023–1026) was not accounted
for by use of a point estimate from mammals (1023), nor was
the known failure of a molecular clock for microsatellite mutation rates [39]. Thus, despite substantial attention,diabolis
age estimates still range from 90 to 20 000 years [38,40]. Further-more, additional outstanding questions about the survival of this species within its habitat remain: was Devils Hole colo-nized only once? Did phenotypic plasticity facilitate adaptation to this harsh environment [41]? How did so many small Death Valley pupfish populations survive inbreeding depression if they were isolated for millions of years [42]?
We examined population structure and gene flow among Death Valley populations, estimated the pupfish mutation rate from a new time-calibrated phylogeny containing all majorCyprinodonlineages, and then used this rate to calibrate a demographic model for the divergence between two closely related pupfish populations in Death Valley and approximate an age for diabolis. We sampled all extant Death Valley and Ash Meadows species and subspecies as well as outgroup species from across theCyprinodonclade (electronic supplemen-tary material, appendix S1). We conducted all analyses at the population level (i.e. each desert spring) and for clarity we refer to these populations by only their subspecies designation and desert spring name. In some cases, the same subspecies was sampled from multiple desert springs and we treated these as different populations. We used double-digest restric-tion-site-associated DNA sequencing (ddRADseq [43]) to genotype over 13 000 loci in 56 individuals. This provided 1.3 million sites for demographic inference, 100 000 times more data than previous studies. Our analyses reshape our understanding of population isolation and speciation in this highly endangered group of extremophile fishes and suggest a surprisingly rapid timescale for speciation, gene-tic assimilation and the evolution of intrinsic reproductive incompatibilities in this group.
2. Material and Methods
(a) Sample collection and bioinformatics
Cyprinodon diabolisis one of the most endangered fish on earth and thus collecting tissue from live animals was impossible. Highly degraded samples putrefied by the 328C water were obtained from dead specimens collected by National Park staff. Other Death Valley samples used archived specimens from a previous study [44]. DNA was extracted with Qiagen Blood & Tissue kits and ddRADseq libraries [43] were prepared as described previously [45]. One hundred and ninety-eight million Illumina HiSeq 2000 single-end 100 bp reads were aligned to theCyprinodon variegatusreference genome (v. 1.0) and called using the Stacks pipeline [46]. Loci genotyped in at least six of the 56 high-coverage individuals and sequenced to a minimum depth of eight reads were used for analyses. Overall mean coverage depth per locus per individual for aligned reads was 138. Further details are provided in the electronic supplementary material, Supplemental Methods.
(b) Population genetic structure and analyses of
gene flow
Population genetic structure among populations was evaluated by principal components analysis of genetic variance and
Bayesian unsupervised clustering using STRUCTURE[47]. Gene flow among populations was estimated using four complemen-tary methods: formal tests of introgression using ABBA/BABA ratios (D-statistics) [48], ancestry proportions across indepen-dent STRUCTURE runs, non-zero migration parameters in our
dadimodel, and by using TREEMIX[49] to estimate secondary gene flow among branches in a maximum-likelihood tree based on the residual covariance in allele frequencies among populations. Although we sampled small numbers of individ-uals per population, this will not necessarily bias our estimates of genetic differentiation owing to the large number of genetic markers sampled per individual [50].
(c) Phylogenetic analyses and time-calibration
We used BEAST (v. 1.8.1; [51]) for phylogenetic inference from 16 567 concatenated 100 bp loci based on a separate Stacks pipeline sampling the most divergent lineages across Cyprino-don. Estimation of divergence times and mutation rates is highly sensitive to the choice of priors [37] and requires internal calibrations [36,52,53], thus we chose the only well-defined internal calibration event known for Cyprinodon (options discussed in [54]): the 8000+200 year age of Laguna Chichan-canab [55,56], an endorheic basin which contains an endemic species flock of Cyprinodon pupfishes (figure 2d; [57]). It is highly unlikely that the Chichancanab species flock diverged before the basin formed based on microsatellite evidence [58,59] and because these species cannot coexist with fish pre-dators found in all neighbouring surface waters (the flock is now extinct due to invasive fishes [54,60]). We conservatively placed a lower bound on the stem age of this clade using a normal prior for the divergence betweenCyprinodon artifrons
(the most closely related species) and the Chichancanab line-age with a mean of 8000 years and standard deviation of 100 years. This age and associated error (95% confidence inter-val (CI):+200 years) were estimated in geological studies of this lake using multiple core samples and multiple lines of evidence, including stable isotope data and shifts from terres-trial to aquatic invertebrate communities [55,56]. Our analysis is still highly dependent on this choice of prior and would support much older ages for diabolis if we used ancient land barriers or other mutation rates for calibration as in pre-vious studies of this group [30,32]. For example, calibration with the human mutation rate would unrealistically place the age of the Laguna Chichancanab species flock at 4.9 Myr (see the electronic supplementary material, Supplemental Methods).
(d) Estimation of the mutation rate in pupfishes
We estimated a median global substitution rate of 5.371027mutations site21yr21using our original time-calibrated
phylo-geny for Cyprinodontidae (electronic supplementary material, table S2). This is much greater than the human genome-wide mutation rate estimated from pedigrees (0.51029
mutations site21yr21: [61]). Differences between human gener-ation time (20–30 years) and pupfish genergener-ation time (less than 4 months in the wild) only partially account for this difference. It is unknown whether this reflects a bias in our RADseq data or a real biological difference in pupfishes, such as increased metabolism in extreme thermal or hypersaline environments [62]. Alternative phylogenetic models, more stringent filtering, and taxon subsets did not substantially change our estimate of the mutation rate (electronic supplementary material, table S2). This
rspb.r
oy
alsocietypublishing.org
Pr
oc.
R.
Soc.
B
283
:
20152334
estimate was also robust to completely rerunning the pipeline from raw reads trimmed to 53 bp to remove later positions with decreased read qualities and estimation of a new time-calibrated phylogeny (electronic supplementary material, figure S4 and table S2).
(e) Demographic modelling
Our time-calibrated phylogeny probably overestimates the age of young taxa such asdiabolisowing to incomplete line-age sorting of only a single concatenated haplotype [35]. Therefore, we used demographic modelling indadi[63] to esti-mate divergence time from allele frequencies. Rather than using an approximate simulation of the likelihood (e.g. [38]),
dadidirectly calculates the likelihood of the joint-site frequency spectrum under various demographic scenarios using a diffusion approximation [63].
Owing to the extreme degradation ofdiabolissamples, we were not able to obtain a good estimate ofdiabolisallele fre-quencies. Instead, we bounded the age of diabolis from the divergence time between the two main population clusters in Ash Meadows: mionectes and amargosae/shoshone/nevadensis
(figure 2a–c) and the older divergence of thesalinus/milleri out-group. Our phylogenetic analyses place the diabolis split in between these two branches (figure 2) or within themionectes
group in the trimmed dataset (electronic supplementary material, figure S4). In both cases, thediabolisdivergence time is very close to themionectesandamargosae/shoshone/nevadensis
divergence time. We then approximated the diabolis age by correcting for the relative difference in branch lengths. None-theless, this approximation is a weakness of our study and should be interpreted with caution.
3. Results
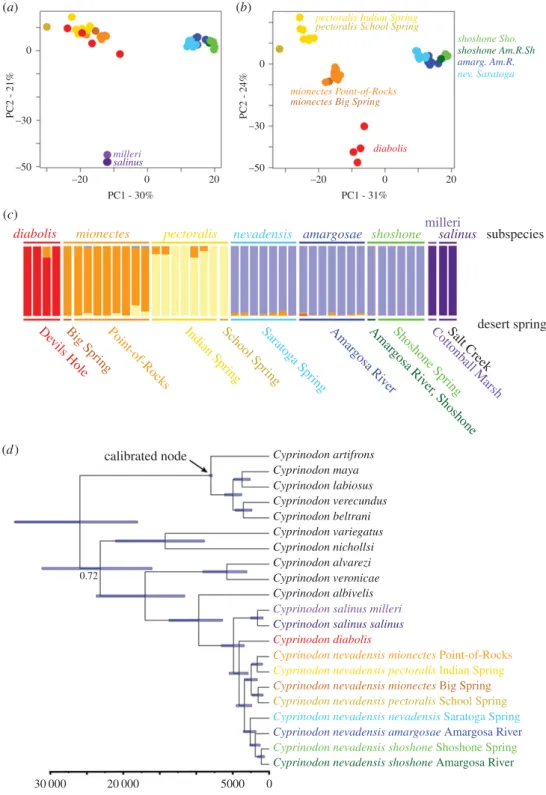
(a) Population structure and genetic diversity
We found evidence of genetic structure among five major groups in Death Valley: diabolis, salinus/milleri, mionectes,
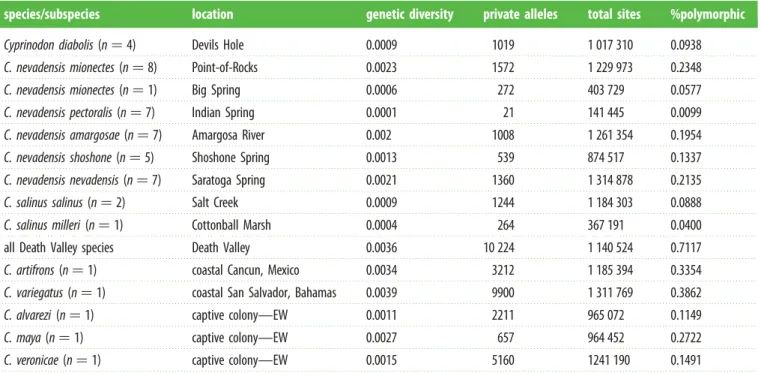
pectoralisandamargosae/shoshone/nevadensis(figure 2a–c; elec-tronic supplementary material, figure S5). This is consistent with the expectation that genetic drift in small populations will accelerate lineage sorting and accumulation of genetic structure [64]. Indeed, Death Valley populations exhibited up to 40-fold lower genetic diversity than coastal Cyprinodon
populations (table 1). The lowest diversity observed inpectoralis
was equal to the lowest genetic diversity ever documented in a wild population ([24]:Lynx lynx) and 10-fold lower than a captive pupfish population extinct in the wild for over 15 years (table 1:Cyprinodon alvarezi[65]). Despite low diversity within each spring, if we pool all Death Valley populations into one large interconnected population, the genetic diversity represented in the entire valley is similar to high diversity coastal populations (table 1).
(b) Time-calibrated phylogeny
Our phylogeny places the stem divergence of Death Valley pupfishes at approximately 10 kyr ago (95% CI: 5–20 kyr ago; figure 2d; electronic supplementary material, figure S4). This interval is centered directly on the most recent flooding of the Death Valley basin from 10 to 35 kyr ago observed in core samples [20]. Death Valley pupfishes shared a common
ancestor with Northeastern Mexican species approximately 17 kyr ago (95% CI: 9–34 kyr ago) and Atlantic coastal popu-lations 25 kyr ago (95% CI: 12–41 kyr ago; figure 2d). These ages are consistent with increased population mixing expected from the formation of large pluvial lakes throughout North America (e.g. Palaeolake Manly: figure 1a) during the most recent glacial period 12–25 kyr ago [66]. It is not apparent how low-lying desert populations could have remained iso-lated within large inland seas because nearly all pupfish species readily interbreed on secondary contact and produce viable offspring (see Discussion).
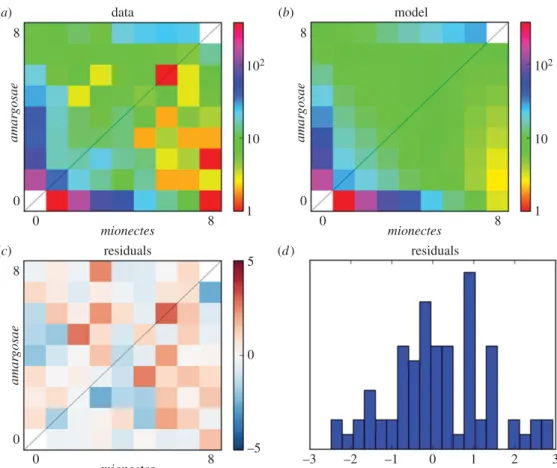
(c) Demographic analysis of divergence time
Our maximum-likelihood estimate of the divergence between
amargosae/shoshone/nevadensis and mionectes was 210 years (95% CI: 60–363; figure 3; electronic supplementary material, table S2). Encouragingly, this interval contains the Great Flood of 1862, the largest recorded rainfall for California and Nevada [67], which probably flooded Ash Meadows and mixed mionectes with amargosae/shoshone/nevadensis popu-lations downstream (figure 1a). We next compared the median divergence time ofdiabolisrelative to the median diver-gence betweenmionectesandamargosae/shoshone/nevadensisin our phylogeny (figure 2d) to rescale an approximate age for
diabolisbased on our demographic model and our mutation rate estimate for pupfishes (electronic supplementary material, table S2: 5.371027site21yr21). We estimate the age ofdiabolis
to be 255 years (electronic supplementary material, table S2: 95% CI: 105–408), suggesting that the current population colo-nized only 159 years before the first historical record of Devils Hole pupfish in 1893 [21]. Alternatively, using the slowest and fastest mutation rates across the range of models and datasets we explored, diabolis ranged in age from 830 years to as young as 105 years (younger than the first historical record), respectively (electronic supplementary material, table S2: max and min 95% confidence boundaries).
(d) Recent gene flow
Unexpectedly, we also found evidence of recent gene flow betweendiabolisand bothamargosaeandpectoralissupported by formal tests for introgression (electronic supplementary material, table S4: amargosae D-statistic¼0.17, p¼0.0007;
pectoralis D-statistic¼0.15, p¼0.002). Cyprinodon diabolis
introgression was also consistent with STRUCTURE analyses (figure 2c) and TREEMIX (electronic supplementary material, figure S1). One amargosae individual contained a median of 1.6% diabolis ancestry overall (figure 2c); however, 95% credible intervals ofdiabolisancestry proportions for this indi-vidual contained zero in seven out of eight independent STRUCTURE runs. Introgression from diabolis into amargosae
was also supported by the residual covariance in allele frequencies between these populations estimated using TREEMIX(electronic supplementary material, figure S1).
The small amount of diabolis ancestry within amargosae
(figure 2c), lack of significant introgression with other Death Valley and Ash Meadows populations besidespectoralis (elec-tronic supplementary material, table S4), and directionality of gene flow inferred by TREEMIX (electronic supplementary material, figure S1) is consistent with gene flow from Devils Hole into Amargosa River sometime after the divergence of amargosae/shoshone/nevadensis and mionectes 210 years ago. Biologists transferred 75diabolisto a spring in southern
rspb.r
oy
alsocietypublishing.org
Pr
oc.
R.
Soc.
B
283
:
20152334
Ash Meadows in 1946 [68] which might explain thepectoralis
signal, but does not appear to explain the amargosae signal since no introgression was observed in the interveningshoshone
population.
Overall, genetic covariance among Death Valley populations indicates additional introgression fromsalinusandpectoralisinto
nevadensis (electronic supplementary material, figure S1) and
suggests overland dispersal on 100-year timescales. Similarly, our demographic model supported significant low levels of migration betweenamargosae/shoshone/nevadensisandmionectes
after divergence (electronic supplementary material, table S3). We also confirmed a previous study [69] which found that a refuge population ofdiaboliswithin a concrete pond hybridized with introducedmionectes(electronic supplementary material, –20
–50 –30 0
0
milleri
milleri
subspecies
desert spring
Salt Creek Cottonball Marsh
Shoshone Spring
Amar
gosa Ri
ve r, Shoshone Amar
gosa Ri
ver Saratoga Spring
School Spring
Indian Spring Point-of-Rocks
Big Spring De
vils Hole diabolis
Cyprinodon artifrons Cyprinodon maya Cyprinodon labiosus Cyprinodon verecundus Cyprinodon beltrani Cyprinodon variegatus Cyprinodon nichollsi Cyprinodon alvarezi Cyprinodon veronicae Cyprinodon albivelis Cyprinodon salinus milleri Cyprinodon salinus salinus Cyprinodon diabolis
Cyprinodon nevadensis mionectes Point-of-Rocks Cyprinodon nevadensis pectoralis Indian Spring Cyprinodon nevadensis mionectes Big Spring Cyprinodon nevadensis pectoralis School Spring Cyprinodon nevadensis nevadensis Saratoga Spring
Cyprinodon nevadensis shoshone Amargosa River
30 000 20 000 5000 0
time (years)
Cyprinodon nevadensis shoshone Shoshone Spring Cyprinodon nevadensis amargosae Amargosa River mionectes pectoralis nevadensis amargosae shoshone salinus
pectoralis Indian Spring pectoralis School Spring
shoshone Sho. shoshone Am.R.Sh amarg. Am.R. nev. Saratoga
diabolis
mionectes Point-of-Rocks mionectes Big Spring
salinus
PC1 - 30%
PC2 - 21%
(a)
(c)
(d) calibrated node
0.72
–50 –30 0
PC2 - 24%
(b)
20 –20 0
PC1 - 31%
20
Figure 2.
Genetic structure (a–c) and time-calibrated phylogenetic tree (d) for Death Valley pupfish populations. (a) First two principal components of genetic
variance for 4679 single nucleotide polymorphisms (SNPs) showing three main clusters of Death Valley populations. (b) Excluding the distant
salinus
populations
reveals four to five distinct genetic clusters. (c) Bayesian clustering analyses using S
TRUCTUREindicating the proportion of shared ancestry among individuals with
k
¼
5 groups for 4679 SNPs, supported by a reduced rate of change of the log-likelihood at this value (electronic supplementary material, table S5). Species and
subspecies designations are indicated on the top row and desert spring systems are indicated on the bottom row, corresponding to sample sites in figure 1a.
(d) Time-calibrated maximum clade credibility tree for the Death Valley populations plus all major lineages across
Cyprinodon. A single haplotype was sampled
from one high-coverage individual per population. All nodes were supported with 100% posterior probability except the one annotated node (0.72). Blue bars
indicate 95% credible intervals for the estimated age of each node. (Online version in colour.)
rspb.r
oy
alsocietypublishing.org
Pr
oc.
R.
Soc.
B
283
:
20152334
figure S2: 85.3%mionectesancestry), despite its isolation from neighbouring spring systems.
4. Discussion
Our population genomic analyses paint a dramatically differ-ent picture of survival in Death Valley pupfishes than previous assumptions. Rather than vicariance-driven specia-tion of isolated relicts surviving millions of years throughout flooding of the entire region lasting thousands of years (figure 1a), we found evidence for frequent overland dispersal and mixing of Death Valley pupfish populations on a scale of hundreds–thousands of years. Although overland dispersal across kilometres of desert by an aquatic organism is difficult to fathom, this view is much more consistent with established theories of population persistence and known hydrological connectivity over historical and geological timescales. For example, our time-calibrated phylogeny reasonably indicates that Death Valley pupfishes were isolated from other desert basins after the drying of the most recent pluvial lake in the region 10 kyr ago [20]. Our demographic analysis of the most recent mixing between two desert spring populations is coinci-dent with the largest recorded rainfall in the area in 1862 [67] which would have flooded the dry riverbed connecting these two habitats (figure 1a). We explored a range of phylogenetic models which suggest thatdiaboliswas isolated aftersalinus/ milleriand only slightly earlier thanmionectesandamargosae/ shoshone/nevadensis, and bound its age between 105 and 830 years (min/max 95% confidence boundaries in the electronic supplementary material, table S2). This very recent origin is consistent with early reports of no unique mitochondrial hap-lotypes indiabolisrelative to other Death Valley pupfishes in the region [33]. However, unbiased sampling of genomic diver-sity by genomic resequencing and divergence time estimation from the distribution of haplotype block lengths [70,71] are needed to conclusively estimate divergence time. Finally,
expectations of rapidly increasing genetic load at the low popu-lation sizes frequently observed in desert springs [42], particularly Devils Hole, suggest that frequent dispersal among these populations may be necessary for their long-term persistence. For example, genetic diversity in Death Valley pupfishes was up to 40-fold lower than coastal popu-lations and included the lowest genetic diversity ever observed in a wild population (table 1:Cyprinodon nevadensis pectoralis was equivalent to L. lynx [24]). Nonetheless, our results suggest that the stem lineage of pupfishes in Death Valley survived at least 5000 years before the appearance of extant populations (figure 2d), possibly by maintaining a stable metapopulation despite frequent local extirpations. This perspective may inform conservation strategies for these fishes. The young age of diabolis is surprising given its large number of unique traits: reduced aggression, sexual dimorph-ism and size; larger eyes; darker coloration; and the absence of pelvic fins [21,22]. For example, although Death Valley pup-fishes do not show exceptional rates of morphological diversification as in the San Salvador and Chichancanab sym-patric radiations (which diversified up to 130 times faster than background rates for certain jaw traits [54]), morphological diversification of the Death Valley clade containing diabolis
was five times faster than background rates in other pupfishes [54], comparable to rates observed for similar traits in coral reef fishes [72] and Malawi cichlids [73] over longer timescales. Some diabolis-like traits were observed in amargosae when reared in the laboratory at 328C on a restricted diet to simulate conditions in Devils Hole, including pelvic fin loss in up to 86% of fish [41]. However, laboratory-rearing experiments indicate that pelvic fin loss indiabolishas a genetic basis (electronic sup-plementary material, figure S3). Combined, these observations suggest that colonists of Devils Hole initially lost pelvic fin expression as a plastic response to higher temperatures and reduced resources, only later evolving a broader norm of reac-tion for pelvic fin loss at lower temperatures. This process,
Table 1.
Genetic diversity (p), private alleles, total nucleotide sites examined and percentage of polymorphic sites in Death Valley pupfishes and additional
pupfish outgroups for comparison (EW
¼
extinct in the wild based on IUCN designation or unpublished observations). (The number of individuals sequenced in
each population is indicated (electronic supplementary material, appendix S1).)
species/subspecies
location
genetic diversity
private alleles
total sites
%polymorphic
Cyprinodon diabolis
(n
¼
4)
Devils Hole
0.0009
1019
1 017 310
0.0938
C. nevadensis mionectes
(n
¼
8)
Point-of-Rocks
0.0023
1572
1 229 973
0.2348
C. nevadensis mionectes
(n
¼
1)
Big Spring
0.0006
272
403 729
0.0577
C. nevadensis pectoralis
(n
¼
7)
Indian Spring
0.0001
21
141 445
0.0099
C. nevadensis amargosae
(n
¼
7)
Amargosa River
0.002
1008
1 261 354
0.1954
C. nevadensis shoshone
(n
¼
5)
Shoshone Spring
0.0013
539
874 517
0.1337
C. nevadensis nevadensis
(n
¼
7)
Saratoga Spring
0.0021
1360
1 314 878
0.2135
C. salinus salinus
(n
¼
2)
Salt Creek
0.0009
1244
1 184 303
0.0888
C. salinus milleri
(n
¼
1)
Cottonball Marsh
0.0004
264
367 191
0.0400
all Death Valley species
Death Valley
0.0036
10 224
1 140 524
0.7117
C. artifrons
(n
¼
1)
coastal Cancun, Mexico
0.0034
3212
1 185 394
0.3354
C. variegatus
(n
¼
1)
coastal San Salvador, Bahamas
0.0039
9900
1 311 769
0.3862
C. alvarezi
(n
¼
1)
captive colony—EW
0.0011
2211
965 072
0.1149
C. maya
(n
¼
1)
captive colony—EW
0.0027
657
964 452
0.2722
C. veronicae
(n
¼
1)
captive colony—EW
0.0015
5160
1241 190
0.1491
rspb.r
oy
alsocietypublishing.org
Pr
oc.
R.
Soc.
B
283
:
20152334
known as genetic assimilation, has rarely been demonstrated in nature [74,75]. Pelvic fin loss may be adaptive in a benthic habi-tat, as suggested for stickleback [37]. However, the haphazard pelvic fin loss observed in several other lineages of Cyprinodon-tiform fishes with no known environmental commonalities (including Empetrichthyids,Megupsilon,Orestias, twoRivulus, one Simpsonichthys and one Aphanius species) is consistent with genetic drift.
Our study adds to an emerging consensus that species ages may frequently be overestimated by time-calibrated mtDNA phylogenies [53,70,76]. There appear to be three main reasons. First, similar demographic modelling studies have consistently found that species divergence times were overestimated in mtDNA phylogenies by millions of years, e.g. in flycatchers [76], due to a systematic bias when sampling a single haplotype per species. Similarly, molecular estimates for the age of polar bears spanned 600 kyr–5 Myr ago using conventional demo-graphic modelling approaches, but were revised to 400 kyr ago, consistent with the fossil evidence, by modelling with
dadi[70]. Second, many studies calibrate with distantly related outgroup fossils or ancient vicariance events, such as the breakup of Gondwanaland, assuming no subsequent dispersal was possible [30,32,77]. For example, the age of the modern bowhead whale was revised from 1.1 Myr ago to 150 kyr ago (1 order of magnitude, similar to this study) when calibrating with radiocarbon dates for ancient DNA samples rather than outgroup fossils [36]. Third, substitution rates appear to be at least an order of magnitude slower in older lineages in both coding and non-coding regions, possibly owing to purifying selection or saturation of mutational hotspots, resulting in the overestimation of species ages when these rates are used to cali-brate more recently diverged taxa [78,79]. Estimates based on
only the single mitochondrial haplotype also contain substan-tially more uncertainty than multi-locus nDNA estimates.
Overestimation of species divergence times challenges existing assumptions about the timescale for speciation. For example, current models assume that the accumulation of postzygotic intrinsic isolation takes millions of years due to substantial work in many groups [80,81], including pupfishes [82,83]. Nearly all studies of rapid speciation in fishes report no intrinsic incompatibilities in hybrid crosses [84–87], most notably in East African cichlid clades diversifying over millions of years [88]. These assumptions have inspired recent models of diversification, such as ephemeral speciation [89] and the million-year-waiting-time for macroevolutionary bursts [90]. However, these long timescales conflict with observations of reproductive incompatibilities segregating in natural populations [91] which predict much shorter times for the evol-ution of intrinsic isolation [92,93]. Indeed, our results suggest that intrinsic genetic incompatibilities documented in two different pupfish species [94,95] evolved within less than 30 000 years. Similarly, recent studies indicate that non-lethal intrinsic genetic incompabitilities abound after secondary con-tact in both swordtail fishes [96] and hominids [93], suggesting that traditional progeny screens are insufficient for detecting subtle incompatibilities. Overall, our study points to the evol-ution of intrinsic incompatibilities, phenotypic change, and genetic assimilation on extremely rapid timescales, challenging existing models of speciation [89,90].
Our results also provide context for the management of highly inbred wild and captive populations [17,97] and the recent decline ofdiabolis. High egg survival rates in captivity ([98]: 55%), rebounding population sizes [23], and lower gen-etic diversity in neighbouring stable populations (table 1) all
0 8
(a) (b)
(c) (d)
–5
–3 –2 –1 0 1 2 3
0
0
8 5
mionectes
0 0 8
8 1
10 102
1 10 102
mionectes
residuals
data model
residuals
amar
gosae
amar
gosae
0 0 8
8
mionectes
amar
gosae
Figure 3.
Demographic model of
amargosae/shoshone/nevadensis
and
mionectes
divergence time inferred from allele frequency data using
dadi. (a) Unfolded joint
site frequency spectrum collapsed to eight chromosomes, based on 578 557 sites. (b) Predicted site frequency spectrum under our demographic model. (c) Residuals
from the demographic model across the site frequency spectrum. (d) Residuals were approximately normally distributed. (Online version in colour.)
rspb.r
oy
alsocietypublishing.org
Pr
oc.
R.
Soc.
B
283
:
20152334
suggest that the decline is not owing to an exceptional genetic load causing mutational meltdown as previously speculated [69]. However, theory predicts that extinction due to increas-ing genetic load is highly likely for this small population over timescales of 100 years or less [13,42]. Combined with the 60 000 year age of Devils Hole, this suggests that pupfish extinction and gene flow may have happened many times before in this unique habitat and that diabolis may not be the first pupfish population to have survived there. Protect-ing the connectivity of this region will be essential for this cycle of rebirth to continue.
Data accessibility.All datasets (SNP matrices) used for this study are
deposited in Dryad: http://dx.doi.org/10.5061/dryad.s30t4.
Authors’ contributions.C.H.M. designed the study, prepared samples for
sequencing, conducted phylogenetic and population genetic ana-lyses, and wrote the manuscript. J.E.C. ran demographic analyses
with dadi and calculated D-statistics. L.H.S. provided logistical support, funding, samples, and motivation for the study. B.J.T. collected most of the samples. All authors commented on the manuscript and gave final approval for publication.
Competing interests.We have no competing interests.
Funding.This study was funded by a research stipend from the Miller
Institute for Basic Research in the Sciences to C.H.M. and a small informal expense fund from the National Park Service.
Acknowledgements. We thank Anthony Echelle for supplying tissue
samples from Indian Spring and the refuge diabolis population; Kevin Wilson, Bailey Gaines, and Jeffrey Goldstein for providing access to preserveddiabolisspecimens; Patrik Nosil, Anthony Echelle, and two anonymous reviewers for insightful comments on the manu-script. We thank the National Park Service and the US Fish and Wildlife Service for permission to conduct this research.
Disclaimer.The findings and conclusions in this article are those of the
author(s) and do not necessarily represent the views of the US Fish and Wildlife Service.
References
1. Tobler M, Schlupp I, Heubel KU, Riesch R, de Leo´n FJG, Giere O, Plath M. 2006 Life on the edge: hydrogen sulfide and the fish communities of a Mexican cave and surrounding waters.Extremophiles 10, 577 – 585. (doi:10.1007/s00792-006-0531-2) 2. Schliewen UK, Tautz D, Pa¨a¨bo S. 1994 Sympatric
speciation suggested by monophyly of crater lake cichlids.Nature6472, 629 – 632. (doi:10.1038/ 368629a0)
3. Wagner CE, Harmon LJ, Seehausen O. 2012 Ecological opportunity and sexual selection together predict adaptive radiation.Nature487, 366 – 369. (doi:10.1038/nature11144)
4. Gavrilets S, Losos JB. 2009 Adaptive radiation: contrasting theory with data.Science323, 732 – 737. (doi:10.1126/science.1157966) 5. Losos J, Mahler D. 2010 Adaptive radiation: the
interaction of ecological opportunity, adaptation, and speciation. InEvolution since Darwin: the first 150 years, pp. 381 – 420. Sunderland, MA: Sinauer Associates. (cited 12 Jan 2015).
6. Lenski RE, Travisano M. 1994 Dynamics of adaptation and diversification: a 10,000-generation experiment with bacterial populations.Proc. Natl Acad. Sci. USA91, 6808 – 6814. (doi:10.1073/pnas. 91.15.6808)
7. Rainey P, Travisano M. 1998 Adaptive radiation in a heterogeneous environment.Nature394, 69 – 72. (doi:10.1038/27900)
8. Bono LM, Gensel CL, Pfennig DW, Burch CL. 2013 Competition and the origins of novelty: experimental evolution of niche-width expansion in a virus.Biol. Lett.9, 20120616. (doi:10.1098/rsbl. 2012.0616)
9. Blount ZD, Borland CZ, Lenski RE. 2008 Historical contingency and the evolution of a key innovation in an experimental population ofEscherichia coli. Proc. Natl Acad. Sci. USA105, 7899 – 7906. (doi:10. 1073/pnas.0803151105)
10. Martin C. 2012 Weak disruptive selection and incomplete pheonytpic divergence in two classic examples of sympatric speciation: Cameroon crater
lake cichlids.Am. Nat.180, E90 – E109. (doi:10. 1086/667586)
11. Martin CH. 2013 Strong assortative mating by diet, color, size, and morphology but limited progress toward sympatric speciation in a classic example: Cameroon crater lake cichlids.Evolution67, 2114 – 2123. (doi:10.1111/evo.12090)
12. Papadopulos ASTet al.2014 Evaluation of genetic isolation within an island flora reveals unusually widespread local adaptation and supports sympatric speciation.Phil. Trans. R. Soc. B369, 20130342. (doi:10.1098/rstb.2013.0342)
13. Lynch M, Conery J, Burger R. 1995 Mutation accumulation and the extinction of small populations.Am. Nat.146, 489 – 518. (doi:10.1086/ 285812)
14. Lanfear R, Kokko H, Eyre-Walker A. 2014 Population size and the rate of evolution.Trends Ecol. Evol.29, 33 – 41. (doi:10.1016/j.tree.2013.09.009) 15. Bijlsma R, Bundgaard J, Boerema AC. 2000 Does
inbreeding affect the extinction risk of small populations? predictions fromDrosophila.J. Evol. Biol.13, 502 – 514. (doi:10.1046/j.1420-9101.2000. 00177.x)
16. Willi Y, Van Buskirk J, Hoffmann AA. 2006 Limits to the adaptive potential of small populations.Annu. Rev. Ecol. Evol. Syst.37, 433 – 458. (doi:10.1146/ annurev.ecolsys.37.091305.110145)
17. Kennedy ES, Grueber CE, Duncan RP, Jamieson IG. 2014 Severe inbreeding depression and no evidence of purging in an extremely inbred wild species: the Chatham Island black robin.Evolution68, 987 – 995. (doi:10.1111/evo.12315)
18. Eldridge MDB, King JM, Loupis AK, Spencer PBS, Taylor AC, Pope LC, Hall GP. 1999 Unprecedented low levels of genetic variation and inbreeding depression in an island population of the black-footed rock-wallabi.Conserv. Biol.13, 531 – 541. (doi:10.1046/j.1523-1739.1999.98115.x)
19. O’Grady JJ, Brook BW, Reed DH, Ballou JD, Tonkyn DW, Frankham R. 2006 Realistic levels of inbreeding depression strongly affect extinction risk in wild
populations.Biol. Conserv.133, 42 – 51. (doi:10. 1016/j.biocon.2006.05.016)
20. Lowenstein T, Li J, Brown C, Roberts S. 1999 200 ky paleoclimate record from Death Valley salt core. Geology27, 3 – 6. (doi:10.1130/0091-7613(1999)027,0003:KYPRFD.2.3.CO;2) 21. Deacon JE, Taylor FR, Pedretti JW, Pedretti W. 1995
Egg viability and ecology of Devils Hole pupfish: insights from captive propagation.Southwest. Nat. 40, 216 – 223.
22. Riggs A, Deacon J. 2002 Connectivity in desert aquatic ecosystems: the Devils Hole story. InProc. Spring-fed Wetl. Important Sci. Cult. Resour. Intermt. Reg., 7 – 9 May 2002, Las Vegas, Nevada (eds DW Sada, SE Sharpe). DHS Publication no. 41210. 23. Beissinger SR. 2014 Digging the pupfish out of its
hole: risk analyses to guide harvest of Devils Hole pupfish for captive breeding.PeerJ2, e549. (doi:10. 7717/peerj.549)
24. Wilson KP, Blinn DW. 2007 Food web structure, energetics, and importance of allochthonous carbon in a desert cavernous limnocrene: Devils Hole, Nevada.West. North Am. Nat.67, 185– 198. (doi:10. 3398/1527-0904(2007)67[185:FWSEAI]2.0.CO;2) 25. Heath AG, Turner BJ, Davis WP. 1993 Temperature
preferences and tolerances of three fish species inhabiting hyperthermal ponds on mangrove islands. Hydrobiologia259, 47–55. (doi:10.1007/BF00005964) 26. Bennett W, Beitinger T. 1997 Temperature tolerance of the sheepshead minnow,Cyprinodon variegatus. Copeia1997, 77 – 87. (doi:10.2307/1447842) 27. Carson EW, De la Maza-Benignos M, de Lourdes
Lozano-Vilano M, Vela-Valladares L, Banda-Villanueva I, Turner TF. 2013 Conservation genetic assessment of the critically endangered Julimes pupfish,Cyprinodon julimes.Conserv. Genet.15, 483 – 488. (doi:10.1007/s10592-013-0548-x) 28. Minckley W, Minckley C. 1986Cyprinodon
pachycephalus, a new species of pupfish (Cyprinodontidae) from the Chihuahuan desert of northern Mexico.Copeia1986, 184 – 192. (doi:10. 2307/1444904)
rspb.r
oy
alsocietypublishing.org
Pr
oc.
R.
Soc.
B
283
:
20152334
29. Hausner M, Wilson K, Gaines D, Suarez F, Scoppettone G, Tyler S. 2014 Life in a fishbowl: prospects for the endangered Devils Hole pupfish (Cyprinodon diabolis) in a changing climate.Water Resour. Res.50, 7020 – 7034. (doi:10.1002/2014WR015511) 30. Echelle AA, Carson EW, Echelle AF, Van Den Bussche
RA, Dowling TE, Meyer A. 2005 Historical biogeography of the New-World pupfish genus Cyprinodon(Teleostei: Cyprinodontidae).Copeia 2005, 320 – 339. (doi:10.1643/CG-03-093R3) 31. Echelle A. 2008 The western North American
pupfish clade (Cyprinodontidae:Cyprinodon): mitochondrial DNA divergence and drainage history. Geol. Soc. Am. Spec. Pap.439, 27 – 38. (doi:10. 1130/2008.2439(02))
32. Hrbek T, Meyer A. 2003 Closing of the Tethys Sea and the phylogeny of Eurasian killifishes (Cyprinodontiformes: Cyprinodontidae).J. Evol. Biol. 16, 17– 36. (doi:10.1046/j.1420-9101.2003.00475.x) 33. Echelle A, Dowling T. 1992 Mitochondrial DNA
variation and evolution of the Death Valley pupfishes (Cyprinodon, Cyprinodontidae).Evolution (N. Y).46, 193 – 206. (doi:10.2307/2409814) 34. Winograd IJ, Coplen TB, Landwehr JM, Riggs AC,
Ludwig KR, Szabo BJ, Kolesar PT, Revesz KM. 1992 Continuous 500,000-year climate record from vein calcite in Devils Hole, Nevada.Science258, 255 – 260. (doi:10.1126/science.258.5080.255) 35. Heled J, Drummond AJ. 2010 Bayesian inference of
species trees from multilocus data.Mol. Biol. Evol. 27, 570 – 580. (doi:10.1093/molbev/msp274) 36. Ho SYW, Saarma U, Barnett R, Haile J, Shapiro B.
2008 The effect of inappropriate calibration: three case studies in molecular ecology. PLoS ONE3, e1615. (doi:10.1371/journal.pone. 0001615)
37. Warnock RCM, Parham JF, Joyce WG, Lyson TR, Donoghue PCJ. 2015 Calibration uncertainty in molecular dating analyses: there is no substitute for the prior evaluation of time priors.Proc. R. Soc. B 282, 20141013. (doi:10.1098/rspb.2014.1013) 38. Reed J, Stockwell C. 2014 Evaluating an icon of
population persistence: the Devil’s Hole pupfish. Proc. R. Soc. B281, 1648 – 1658. (doi:10.1098/rspb. 2014.1648)
39. Ellegren H. 2004 Microsatellites: simple sequences with complex evolution.Nat. Rev. Genet.5, 435 – 445. (doi:10.1038/nrg1348)
40. Smith G, Dowling T, Gobalet K, Lugaski T, Shiozawa D, Evans P. 2002 Biogeography and rates of evolution of Great Basin fishes. InGreat Basin aquatic systems history: Smithsonian contributions to the earth sciences(eds R Hershler, D Madsen, D Curry), pp. 175 – 234. New York, NY: Food and Agriculture Organization of the United Nations. 41. Lema S, Nevitt G. 2006 Testing an ecophysiological
mechanism of morphological plasticity in pupfish and its relevance to conservation efforts for endangered Devils Hole pupfish.J. Exp. Biol.209, 3499 – 3509. (doi:10.1242/jeb.02417)
42. Lynch M, Conery J, Burger R. 1995 Mutational meltdowns in sexual populations.Evolution49, 1067 – 1080. (doi:10.2307/2410432)
43. Peterson BK, Weber JN, Kay EH, Fisher HS, Hoekstra HE. 2012 Double digest RADseq: an inexpensive method for de novo SNP discovery and genotyping in model and non-model species.PLoS ONE7, e37135. (doi:10.1371/journal.pone.0037135) 44. Duvernell DD, Turner BJ. 1998 Variation and
divergence of death valley pupfish populations at retrotransposon-defined loci.Mol. Biol. Evol.16, 363 – 371. (doi:10.1093/oxfordjournals.molbev. a026117)
45. Martin CH, Cutler JS, Friel JP, Dening T, Coop G, Wainwright PC. 2015 Complex histories of repeated colonization and hybridization cast doubt on the clearest examples of sympatric speciation in the wild.Evolution(N. Y).69, 1406 – 1422. 46. Catchen J, Hohenlohe PA, Bassham S, Amores A,
Cresko WA. 2013 Stacks: an analysis tool set for population genomics.Mol. Ecol.22, 3124 – 3140. (doi:10.1111/mec.12354)
47. Pritchard JK, Stephens M, Donnelly P. 2000 Inference of population structure using multilocus genotype data.Genetics155, 945 – 959. 48. Heliconius T, Consortium G. 2012 Butterfly genome
reveals promiscuous exchange of mimicry adaptations among species.Nature487, 94 – 98. (doi:10.1038/nature11041)
49. Pickrell JK, Pritchard JK. 2012 Inference of population splits and mixtures from genome-wide allele frequency data.PLoS Genet.8, e1002967. (doi:10.1371/journal.pgen.1002967)
50. Willing E-M, Dreyer C, van Oosterhout C. 2012 Estimates of genetic differentiation measured by F(ST) do not necessarily require large sample sizes when using many SNP markers.PLoS ONE7, e42649. (doi:10.1371/journal.pone.0042649) 51. Drummond AJ, Rambaut A. 2007 BEAST: Bayesian
evolutionary analysis by sampling trees.BMC Evol. Biol.8, 1 – 8. (doi:10.1186/1471-2148-7-214) 52. Ho SYW, Phillips MJ. 2009 Accounting for calibration
uncertainty in phylogenetic estimation of evolutionary divergence times.Syst. Biol.58, 367 – 380. (doi:10.1093/sysbio/syp035)
53. Ho SYW, Phillips MJ, Cooper A, Drummond AJ. 2005 Time dependency of molecular rate estimates and systematic overestimation of recent divergence times.Mol. Biol. Evol.22, 1561 – 1568. (doi:10. 1093/molbev/msi145)
54. Martin CH, Wainwright PC. 2011 Trophic novelty is linked to exceptional rates of morphological diversification in two adaptive radiations of Cyprinodonpupfish.Evolution65, 2197 – 2212. (doi:10.1111/j.1558-5646.2011.01294.x)
55. Covich A, Stuiver M. 1974 Changes in the oxygen 18 as a measure of long-term fluctuations in tropical lake levels and molluscan populations.Limnol. Oceanogr.19, 682 – 691. (doi:10.4319/lo.1974. 19.4.0682)
56. Hodell D, Curtis J, Brenner M. 1995 Possible role of climate in the collapse of Classic Maya civilization. Nature375, 391 – 394. (doi:10.1038/375391a0) 57. Humphries J, Miller RR. 1981 A remarkable species
flock of pupfishes, genusCyprinodon, from Yucatan, Mexico.Copeia1981, 52– 64. (doi:10.2307/1444040)
58. Strecker U. 2006 Characterization and cross-species amplification of microsatellite loci in aCyprinodon species flock.Mol. Phylogenet. Evol.39, 843 – 846. (doi:10.1016/j.ympev.2006.01.008)
59. Strecker U, Meyer CG, Sturmbauer C, Wilkens H. 1996 Genetic divergence and speciation in an extremely young species flock in Mexico formed by the genusCyprinodon(Cyprinodontidae, Teleostei). Mol. Phylogenet. Evol.6, 143 – 149. (doi:10.1006/ mpev.1996.0066)
60. Strecker U. 2006 The impact of invasive fish on an endemicCyprinodonspecies flock (Teleostei) from Laguna Chichancanab, Yucatan, Mexico.Ecol. Freshw. Fish15, 408 – 418. (doi:10.1111/j.1600-0633.2006.00159.x)
61. Scally A, Durbin R. 2012 Revising the human mutation rate: implications for understanding human evolution.Nat. Rev. Genet.13, 824. (doi:10. 1038/nrg3353)
62. Brix KV, Grosell M. 2012 Comparative characterization of Naþtransport inCyprinodon variegatus variegatusandCyprinodon variegatus hubbsi: a model species complex for studying teleost invasion of freshwater.J. Exp. Biol.215, 1199 – 1209. (doi:10.1242/jeb.067496)
63. Gutenkunst R, Hernandez R. 2009 Inferring the joint demographic history of multiple populations from multidimensional SNP frequency data.PLoS Genet. 5, e1000695. (doi:10.1371/journal.pgen.1000695) 64. Fisher R. 1930Genetical theory of natural selection.
Oxford, UK: Clarendon Press.
65. Contreras-Balderas S, Almada-Villela P. 1996 Cyprinodon alvarezi. The IUCN Red List of Threatened Species. Gland, Switzerland: International Union for Conservation of Nature. 66. Easterbrook D. 1999Surface processes and landforms.
Upper Saddle River, NJ: Prentice-Hall, Inc. 67. Brewer W. 1930Up and down California in 1860 –
1864. New Haven, CT: Yale University Press. 68. Rivers IR. 1950 The Dryopoidea known or expected
to occur in the Nevada Area (Coleoptera).Wasmann J. Biol.8, 97 – 112.
69. Martin AP, Echelle AA, Zegers G, Baker S, Keeler-Foster CL. 2011 Dramatic shifts in the gene pool of a managed population of an endangered species may be exacerbated by high genetic load.Conserv. Genet.13, 349 – 358. (doi:10.1007/s10592-011-0289-7)
70. Liu S.et al.2014 Population genomics reveal recent speciation and rapid evolutionary adaptation in polar bears.Cell157, 785 – 794. (doi:10.1016/j.cell. 2014.03.054)
71. Harris K, Nielsen R. 2013 Inferring demographic history from a spectrum of shared haplotype lengths.PLoS Genet.9, e1003521. (doi:10.1371/ journal.pgen.1003521)
72. Price SA, Holzman R, Near TJ, Wainwright PC. 2011 Coral reefs promote the evolution of morphological diversity and ecological novelty in labrid fishes.Ecol. Lett.14, 462 – 469. (doi:10.1111/j.1461-0248.2011. 01607.x)
73. Hulsey CD, Mims MC, Parnell NF, Streelman JT. 2010 Comparative rates of lower jaw diversification in
cichlid adaptive radiations.J. Evol. Biol.23, 1456 – 1467. (doi:10.1111/j.1420-9101.2010.02004.x) 74. Pfennig DW, Wund MA, Snell-Rood EC, Cruickshank
T, Schlichting CD, Moczek AP. 2010 Phenotypic plasticity’s impacts on diversification and speciation. Trends Ecol. Evol.25, 459 – 467. (doi:10.1016/j.tree. 2010.05.006)
75. Braendle C, Flatt T. 2006 A role for genetic accommodation in evolution?BioEssays28, 868 – 873. (doi:10.1002/bies.20456) 76. Nadachowska-Brzyska K, Burri R, Olason PI,
Kawakami T, Smeds L, Ellegren H. 2013 Demographic divergence history of pied flycatcher and collared flycatcher inferred from whole-genome re-sequencing data.PLoS Genet.9, e1003942. (doi:10.1371/journal.pgen.1003942)
77. Friedman M, Keck BP, Dornburg A, Eytan RI, Martin CH, Hulsey CD, Wainwright PC, Near TJ. 2013 Molecular and fossil evidence place the origin of cichlid fishes long after Gondwanan rifting. Proc. R. Soc. B280, 20131733. (doi:10.1098/rspb. 2013.1733)
78. Woodhams M. 2006 Can deleterious mutations explain the time dependency of molecular rate estimates?Mol. Biol. Evol.23, 2271 – 2273. (doi:10. 1093/molbev/msl107)
79. Ho SYW, Lanfear R, Bromham L, Phillips MJ, Soubrier J, Rodrigo AG, Cooper A. 2011 Time-dependent rates of molecular evolution.Mol. Ecol. 20, 3087 – 3101. (doi:10.1111/j.1365-294X.2011. 05178.x)
80. Bolnick DI, Turelli M, Lo´pez-Ferna´ndez H, Wainwright PC, Near TJ. 2008 Accelerated mitochondrial evolution and ‘Darwin’s corollary’: asymmetric viability of reciprocal F1 hybrids in Centrarchid fishes.Genetics178, 1037 – 1048. (doi:10.1534/genetics.107.081364)
81. Coyne JA, Orr HA. 2004Speciation. Sunderland, MA: Sinauer Associates.
82. Turner B, Liu R. 1977 Extensive interspecific genetic compatibility in the New World killifish genus Cyprinodon.Copeia1977, 259 – 269. (doi:10.2307/ 1443907)
83. Cokendolpher JC. 2012 Hybridization experiments with the genusCyprinodon(Teleostei: Cyprinodontidae), vol. 1980, pp. 173 – 176. American Society of Ichthyologists and Herpetologists (http://www.jstor.org/s). 84. Martin CH, Wainwright PC. 2013 Multiple fitness
peaks on the adaptive landscape drive adaptive radiation in the wild.Science339, 208 – 211. (doi:10.1126/science.1227710)
85. Arnegard ME.et al.2014 Genetics of ecological divergence during speciation.Nature511, 307 – 311. (doi:10.1038/nature13301) 86. Schluter D. 2000Ecology of adaptive radiation.
Oxford, UK: Oxford University Press. 87. Seehausen O, Wagner CE. 2013 Speciation in
freshwater fishes.Annu. Rev. Ecol. Evol. Syst.45, 621 – 651. (doi:10.1146/annurev-ecolsys-120213-091818)
88. Stelkens RB, Young KA, Seehausen O. 2010 The accumulation of reproductive incompatibilities in African cichlid fish.Evolution64, 617 – 633. (doi:10. 1111/j.1558-5646.2009.00849.x)
89. Rosenblum EB, Sarver BAJ, Brown JW, Des Roches S, Hardwick KM, Hether TD, Eastman JM, Pennell MW, Harmon LJ. 2012 Goldilocks meets Santa Rosalia: an ephemeral speciation model explains patterns of diversification across time scales.Evol. Biol.39, 255 – 261. (doi:10.1007/s11692-012-9171-x) 90. Uyeda JC, Hansen TF, Arnold SJ, Pienaar J. 2011 The
million-year wait for macroevolutionary bursts. (doi:10.5061/dryad.7d580)
91. Cutter AD. 2012 The polymorphic prelude to Bateson-Dobzhansky-Muller incompatibilities.Trends Ecol. Evol.27, 209 – 218. (doi:10.1016/j.tree.2011. 11.004)
92. Wright KM, Lloyd D, Lowry DB, Macnair MR, Willis JH. 2013 Indirect evolution of hybrid lethality due to linkage with selected locus inMimulus guttatus. PLoS Biol.11, e1001497. (doi:10.1371/journal.pbio. 1001497)
93. Sankararaman S, Mallick S, Dannemann M, Pru¨fer K, Kelso J, Pa¨a¨bo S, Patterson N, Reich D. 2014 The genomic landscape of Neanderthal ancestry in present-day humans.Nature507, 354 – 357. (doi:10.1038/nature12961)
94. Tech C. 2006 Postzygotic incompatibilities between the pupfishes,Cyprinodon elegansandCyprinodon variegatus: hybrid male sterility and sex ratio bias. J. Evol. Biol.19, 1830 – 1837. (doi:10.1111/j.1420-9101.2006.01173.x)
95. Haas R. 1979 Intergeneric hybridization in a sympatric pair of Mexican cyprinodontid fishes. Copeia1979, 149 – 152. (doi:10.2307/1443742) 96. Schumer M, Cui R, Powell DL, Dresner R, Rosenthal
GG, Andolfatto P. 2014 High-resolution mapping reveals hundreds of genetic incompatibilities in hybridizing fish species.Elife3, 1 – 21. (doi:10. 7554/eLife.02535)
97. Hedrick P, Kalinowski S. 2000 Inbreeding depression in conservation biology.Annu. Rev. Ecol. Syst.31, 139 – 162. (doi:10.1146/annurev.ecolsys. 31.1.139)
98. Feuerbacher O, Weissenfluh D, Chaudoin A, Barlics R, Simons L, Linares-Casenave J. 2014 Ash Meadows fish conservation facility: establishing a Devils Hole pupfish population.Proc. Desert Fishes Counc. Ann. Meet., 21 November, San Jose de Cabo, Baja, California Sur, Mexico.