Oikos 000: 19, 2006 doi: 10.1111/j.2006.0030-1299.14655.x, Copyright#Oikos 2006, ISSN 0030-1299 Subject Editor: Pia Mutikainen, Accepted 5 September 2006
Combining population genetics and demographical approaches
in evolutionary studies of plant mating systems
Pierre-Olivier Cheptou and Daniel J. Schoen
P.-O. Cheptou1 ([email protected]), UMR 5175 CEFE-Centre d’Ecologie Fonctionnelle et Evolutive (CNRS), 1919 Route de Mende, FR-34293 Montpellier Cedex 05, France. D. J. Schoen, Dept of Biology, McGill Univ., 1205, Dr. Penfield Avenue, Montreal, Quebec H3A 1B1, Canada.
The striking amount of variation in the mating systems of higher plants has stimulated fruitful research by both ecologists and population geneticists. Historically, these two schools of thought have developed independent theoretical treatments and empirical approaches to account for the evolution of such diversity. We highlight the approach adopted by each field. Population geneticists have developed an approach centred on gene properties of individuals and their role on the evolution of self-fertilisation (transmission rules and the deleterious role of mutations), while ecologists have mostly focused on demographic properties of self-fertilisation (seed production, colonisation ability of selfers). As a result, the two approaches sometimes use conflicting notions of fitness. The recent empirical advances on inbreeding depression, a topic typically motivated by population genetic questions, have emphasized the need to adopt a demographical perspective for fitness. In this paper, we suggest generalizing this approach in mating system evolution and we expect further improvements by integrating demographic and genetics perspectives.
For more than two centuries, the diversity of plant mating systems has captured the curiosity and imagina-tion of biologists. Changes in the mating system are among the most frequent evolutionary transitions in plants. Not only is the mating system a central focus for the study of adaptation (e.g. adaptive changes in flower morphology related to plantpollinator interactions), but it also plays an important role in determining the distribution of genetic variation in populations (Baker 1955, Stebbins 1957, Holsinger 2000).
The adaptive nature of plant mating systems has been studied intensively, especially in hermaphroditic species, which represent more than 70% of the plant kingdom (Yampolsky and Yampolsky 1922). One focus has been on the evolution of self-fertilisation. Interest-ingly and though their scientific approach differ, both ecologists and population geneticists have contributed to our current understanding of the evolution of plant mating systems (Barrett and Harder 1996). Their studies have led to extensive theoretical developments (especially a large body of population genetics theory
for mating system evolution) along with many experi-mental tests of theoretical hypotheses. Over the years, an important body of data on variation in selfing rates in the Angiosperms has also been collected (Vogler and Kalisz 2001).
As noted by several authors (Uyenoyama et al. 1993, Barrett and Harder 1996, Holsinger 1996), these two traditions have been developed along independent paths, and while a number of researchers have com-bined tools from ecology and population genetics in studies of mating system evolution. It would be interesting to go one step further, since to date, despite numerous models and hypotheses, neither ecological nor genetic theory have provided a completely adequate explanation for the selection and maintenance of the enormous range of selfing rates found in hermaphro-ditic plants (Barrett and Harder 1996).
In this paper, our aim is to focus on the logic of the arguments developed in mating system theory and to clarify inconsistencies. We propose that adopting a demographical perspective for fitness components may be a way to unify the different factors affecting
self-fertilisation and to consider its evolution under general ecological scenarios.
Reviewing self-fertilisation theory
As reviewed by Holsinger (1996), the selective advan-tage of selfing can be viewed as arising in one of two ways (Fig. 1):
1. selfing increases ovule success when seed produc-tion is limited by pollen transfer (reproductive assurance hypothesis or fertility advantage) 2. selfing exhibits an intrinsic advantage in pollen
transmission (automatic selection or cost of out-crossing).
The first principle was proposed by Darwin (1876) who focused on self-pollination as the means of ensuring seed set either when outcrossing partners are absent or when pollinators are scarce. This has been referred to as the ‘‘reproductive assurance hypothesis’’ (Jain 1976). The second principle, based on gene transmission mechanisms, was proposed by Ronald Fisher (Fisher 1941), who argued that genes favoring selfing (mating system modifiers) are automatically selected because they benefit from a 50% transmission advantage compared to ‘‘outcrossing’’ genes. On the other hand, the deleterious effect of inbreeding on progeny perfor-mance (or inbreeding depression) (Knight 1799), is considered to be the main factor selecting against selfing. These three general components: pollen limitation, the cost of outcrossing and inbreeding depression are the cornerstones of most theories for the evolution of self-fertilisation (Lloyd 1979, 1980).
Basing one’s argument on the most influential papers (most highly cited) during the period 1985 to 2000, one of the most important theoretical developments in this field was the recognition of the importance of consider-ing both the cost of outcrossconsider-ing and inbreedconsider-ing depression and of including aspects relevant to the genetic architecture of inbreeding depression (Lande and Schemske 1985, Holsinger 1988, Charlesworth and Charlesworth 1990, Charlesworth et al. 1990, 1991). This studies have led to the important notion that inbreeding depression is dynamically linked to selfing rate through purging of deleterious mutations (though see Byers and Waller 1999), as well as the potential importance of genetic associations between fitness loci and modifier loci (Holsinger 1991a). Also, there have been important theoretical insights about the cost of outcrossing. First, Uyenoyama (1986) pointed out that the cost of outcrossing is less than 50% when outcrossed partners are related (biparental inbreeding; Ronfort and Couvet 1995). When outcrossing occurs among relatives the automatic advantage of selfing is no longer 50% but
less. Second, the cost of outcrossing may be reduced as a result of ‘‘pollen discounting’’, i.e. when selfers con-tribute less to outcrossing as pollen donors than out-crosses do. Of interest, Holsinger (1991b) analysed the evolution of selfing when pollen export is dynamically linked to selfing rate (the ‘‘mass action model’’) and showed that partial self-fertilisation is evolutionarily stable whenever selfing can evolve, thus demonstrating the fundamental importance of pollination biology for a
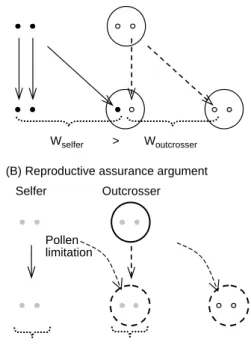
(A) Fisher’s argument
Wselfer > outcrosser
Wselfer > outcrosser
(B) Reproductive assurance argument Selfer Selfer Outcrosser Pollen limitation Outcrosser W W
Fig. 1. Two principles for the advantages of self-fertilization. (A) Mating system modifier genes are represented by dots; an allele coding for self-fertilization is shown as a black dot, while an allele coding for outcrossing is shown as an open dot. Transmission pathways from parent to offspring are shown as arrows; solid arrows represent gene transmission to progeny by the parent capable of self-fertilization, while dashed arrows represent transmission pathways for the outcrossing parent. The recognition that genes are transmitted by pollen and ovules leads to a definition of fitness expressed as the number of genes transmitted to the progeny (gene counting approach). Because the number of pollen grains produced is assumed to be large relative to the number of ovules (Bateman’s principle), a selfing genotype enjoy a 50% advantage in gene transmission relative to a outcrossing genotype). This results in the cost of outcrossing (Fisher 1941). (B) Reproductive assurance advantage of selfing. The large circles represent individual parents and their progeny. The arrows represent the production of discrete offspring by parents, with solid arrows representing pollinator-independent offspring produc-tion, and dashed arrows representing pollinator-dependent offspring production. Selfing provides an advantage by ensuring offspring production when lack of cross-pollinators limits seed set. This results in reproductive assurance.
proper understanding of the cost of outcrossing. All these developments share a common viewpoint, that is, fitness components are analysed as a genetic property of individuals (number of genes transmitted and deleter-ious effects of genes). At the same time, this approach has often been free of the demographical structure linked to mating system evolution and its effect on fitness components, which imposes a number of limita-tions on the scenarios considered (below).
An alternative tradition in mating system studies has focused on the importance of pollen limitation and, in particular, on the demographic impact of selfing. Specifically, Darwin’s ideas (below) were largely popu-larized by Baker (1955) and Baker (1967) in an island biology context. Baker proposed that: ‘‘With a self-compatible individual a single propagule is sufficient to start a sexually reproducing colony (after long distance dispersal), making its establishment much more likely than if the chance of the two self-incompatible yet cross-compatible individuals sufficiently close together spatially and temporally is required’’. Though Baker did not explicitly mention demography, he implicitly focused on the problems posed by low population densities for ensuring outcrossing during colonisation. More recently, the theoretical formalisation of Baker’s law by Pannell and Barrett (1998) explicitly analysed the demographic consequences of outcrossing and the importance of a minimum number of colonizers in a metapopulation to maintain outcrossing. Following this logic, the advantage of selfing in Pannell and Barrett’s model is measured in terms of seed production per individual because they focused on the role of selfing in colonisation ability. In contrast to the population genetic tradition which focuses on the number of genes transmitted, ecological approaches have emphasized on demographical factors such as population density and the colonisation process as important selection pressures driving mating system evolution.
If both fields have identified the major fitness components of selfing, it is fundamental to keep in mind that the automatic advantage of selfing and the fertility advantage do not apply to the same level of selection (see Lloyd 1977 and 1979 for a thoughtful discussion). On the one hand, automatic selection is linked to the number of genes transmitted by an individual (maternal and paternal contribution) and the evolutionary processes is considered as a change in gene frequency. This method is the relevant metrics for analysing individual selection and is classically used in population genetic models cited below. On the other hand, fertility advantage undeniably contributes to fitness but does not match individual gene number (maternal contribution only) and thus does not con-stitute the relevant metrics for analysing evolutionary processes (below). In particular, by emphasizing the fact that the rate of colonisation is favoured by selfing,
Baker’s law (Pannell and Barrett’s 1998 metapopulation model) considers that selection is governed by group selection (Lloyd 1979).
The way the fertility advantage of selfing is con-sidered in ecological studies is particularly telling and may potentially lead to confusion. Indeed, it is often noted that an outcrosser will be at a disadvantage when pollinators are scarce because it produces few seeds (Klips and Snow 1997, Herrera et al. 2001, Anderson et al. 2003, Schueller 2004, Ward et al. 2005). This is true, but it must be reminded that when focusing on genes, each outcrossed seeds (maternal contribution) as well as each sired seeds in the population (paternal contributions) contain exactly one copy of the out-crossing individual. Yet ecological studies seldom note that the outcrosser will be at disadvantage because it also exports less pollen,
We think that such an omission does not help to integrate the selective advantage of reproductive assur-ance in evolutionary models.
If we take a look at the bibliographic for the last 60 years (Source ISI Research), it shows that the seminal work of Fisher (1941) (335 citations) is mostly cited in genetically-oriented journals, whereas Baker (1955) (366 citations) or Baker (1967) (131 citations) are mostly cited in plant science journals. Moreover, papers citing both Baker (1955) and Fisher (1941) are few (54 citations) and none of the major theoretical contribu-tions (cited below) cite both.For the sake of simplicity, many models have considered situations where demo-graphy can be neglected. However, recent empirical results have demonstrated that demography cannot be neglected since classical mating system parameters have to be considered in the light of demographical interac-tions among individuals. That is what we develop in the next part.
Inbreeding depression in wild hermaphroditic populations: the need for integrating population genetics and demography
Some light can be shed on the necessity to include demographical components of fitness by considering the way in which inbreeding depression has been treated in studies of mating system evolution. First described by Thomas Knight (1799) more than 200 years ago, inbreeding depression is expressed at the individual level, and is interpreted as arising from increased homozygosity (Charlesworth and Charles-worth 1987). In hermaphrodites, inbreeding depression is classically defined as d1 Ws
Wout; where Ws and Wout are the performances of selfed and outcrossed
progeny respectively; e.g. biomass, number of flowers, survival, etc.).
Theory for the evolution of mating systems has stimulated many experimental measures of inbreeding depression (reviewed by Husband and Schemske 1996). The motivations for such studies include assessing how inbreeding depression acts to oppose the selection of selfing (Lloyd 1979) as well as understanding the genetic basis of inbreeding depression (Byers and Waller 1999). Unfortunately, other factors affecting inbreeding depression values (e.g. competition) have sometimes been considered as mere ‘complications’, things that should be avoided when studying inbreeding depression (reviewed by Barrett and Harder 1996).
Indeed, the experimental measure of inbreeding depression raises a number of challenges. For instance, consider one example of the analysis of inbreeding depression of survival to flowering in annual plants, where the environment could, in theory, play an extreme role. If growth conditions are very good for growth and survival, both inbred and outbred plants may survive until the end of the life cycle, and one would conclude that there is no inbreeding depression for survivorship. On the other hand, with intermediate mortality rates (e.g. due to competition among indivi-duals or environmental stress), experimental results would likely suggest a positive inbreeding depression value. And lastly, if all plants die before reproduction, (e.g. because of stressful environmental conditions), the experiment would give no information about inbreed-ing depression. Though these simple examples may seem extreme, they demonstrate that in addition to genetic architecture, the estimated magnitude of in-breeding depression must depend on the ecological conditions acting on individuals (physical stress or competition). To our knowledge, no theoretical models have included environmental effects on inbreeding depression when studying the genetic architecture of ID, though empirical results on this point are now clearly established (Armbruster and Reed 2005). We believe that taking into account this phenomenon is crucial for understanding the selective effect of inbreed-ing depression in natural populations. Indeed, it can modify the purging process (Bijlsma et al. 1999) as well as evolutionary predictions concerning selfing. In our own experimental work we have illustrated the im-portance of demographical feedbacks on inbreeding depression in the case when the environmental condi-tions experienced by individuals in a population are linked to the evolution of the reproductive system itself (Cheptou and Schoen 2003). In that study we measured inbreeding depression in competing stands and found that the relative composition of inbred and outbred plants (which in natural populations, is determined directly by population selfing rate) sub-stantially influences the magnitude of inbreeding depression (Fig. 2). In this context, the classical balance between the cost of outcrossing and inbreeding
depres-sion ought to be reinterpreted in the light of feedback from intraspecific competition, itself a function of the population’s rate of selfing (Cheptou and Dieckmann 2002). In other words, a gene favoring selfing may modify the competition environment in which its fitness is determined.
We would like to emphasize that including demo-graphical components of fitness into the population genetics framework may help to generalise the selection on self-fertilisation under general ecological scenarios.
An example: the evolution of self-fertilisation under stochastic pollination environment
In the first part, we emphasized the fact that, though reproductive assurance and the cost of outcrossing select for selfing, they do not operate on the same level of selection. As a consequence, their joint effect is not necessarily trivial. Adopting a demographical view of fitness may help to clarify selection processes and avoid oversimplistic (and intuitive) arguments for the selec-tion of selfing. We illustrate this point by analysing the selection of self-fertilisation under stochastic pollination environment.
When focusing on the role of reproductive assur-ance, people have often focused on the uncertainty of the pollination environment (temporal heterogeneity) (Baker 1967, Herlihy and Eckert 2002, Kalisz and Vogler 2003) and have considered selfing as a way to maximise seed production (Burd 1995). In population genetics models (Lande 1982, Charlesworth 1994), the classical assumption is the stability of environment, which rules out stochastic variations and density dependent effects-and so the exploration of evolution-ary processes under those scenarios has been left out (Caswell 2001). The conclusions drawn from popula-tion genetics models built on stable environment assumptions are not valid under general demographic scenarios. Note that taking into account general ecological scenarios may be particularly relevant as regards the maintenance of mixed mating systems because it often leads to frequency- and density-dependant fitness (Heino et al. 1998), a necessary conditions for negative feedbacks to arise (Goodwillie et al. 2005).
Here we define fitness components in a demogra-phical context, including for instance density-depen-dence or stochastic environments and integrate them into a population genetics framework in order to derive evolutionary predictions (Metz et al. 1992, Brommer 2000). Let us consider a simple model including pollen limitation (reproductive assurance) inspired from Lloyd’s (1979) work, and extending it to stochastic environments a variant of Morgan and Wilson’s (2005) model. Thus, consider a hermaphroditic annual
plant population experiencing inbreeding depression and pollinator limitation. According to the classical population genetics model, the fitness of a rare mutant with a selfing rate r in a resident population with selfing rate R can be measured as the number of genes transmitted to the next generation, that is:
Wheredis the inbreeding depression parameter and e characterizes pollinator efficiency, ‘‘the proportion of available ovules fertilized with the aid of an external agent’’ (Lloyd 1979).From the perspective of reproduc-tive assurance, we consider thate may fluctuate in time as the result of among-year variation in pollinator
Gene transmission via Selfed ovules Out. ovules Pollen export
) ( ) ( ) , , (r Rt r r R et W 2 1 2 1 1 δ
Fitness of inbred progeny Fitness of outbred progeny (a)
(b)
0.0 0.2 0.4 0.6 0.8 1.0
Number of flowers (for focal plants)
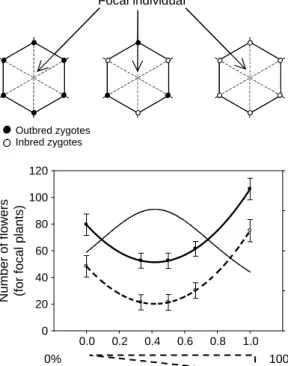
0 20 40 60 80 100 120 Inbreeding depression 0.0 0.2 0.4 0.6 0.8 0% 100% 100% (Inbred neighboors) 0% (Outbred neighboors) Focal individual Outbred zygotes Inbred zygotes Outbred populations Mixed populations Inbred populations
Fig. 2. Experimental evidence of ecological feedback on fitness driven by the mating system. Experiments on the partial selfer Amsinckia douglasiana(s/0.25). In partially selfing annual plant population, the population selfing rate in a large population is
defined as the proportion of inbred and outbred individuals. Consider the focal individual shown in (A). Ecological interactions (competition) in a highly selfing population occur mainly among inbreds, whereas in a highly outcrossing population, they occur mainly among outbreds. Cheptou and Schoen (2003) measured the performances of focal plants (inbred and outbred) by varying the proportion of inbred and outbred plants (on the hexagon), thus simulating competition from complete outcrossing to complete selfing. (B) shows experimental results (fitted number of flowers produced by focal plants: mean, SE) as a function of the proportion of inbred neighboors in a competing stand (310 plants m2) inAmsinckia douglasiana. Absolute fitness for inbred (dotted line) and oubred (solid line) as well as inbreeding depression (d1Wself
Wout:thin line) changes with the frequency of inbred plants, showing that the population selfing rate feedbacks on individual performances and on inbreeding depression as a result of ecological interactions in the population. Note: the possibility that inbreeding depression can be influenced by the composition of the stand, which was rediscovered by Schmitt and Ehrhardt (1990) was anticipated by Darwin (1876) who wrote:‘‘WithNicotianathe crossed were to the self-fertilized in height, when grown extremely crowded together in pots, as 100 to 54; when grown much less crowded in pots as 100 to 66, and when grown in the open ground, so as to be subjected to but little competition, as 100 to 72.’’
activity (Kalisz and Vogler 2003). As a particular case in point, when environmental conditions are constant, the evolution of selfing is favored if 1de
20 (Lloyd 1979). Next, let’s assume instead, that there is a stochastic pollination process resulting from among-year variation in the pollinator population. Specifically, each year, pollination efficiency, e, is taken from a probability distribution (with mean/e¯ and variance
s2). If the population demography is assumed to be globally stable on an ecological timescale--neither goes extinct nor grows exponentially-then, the direction of selection for self-fertilisation is given by the sign of
(1de=2)f(R):s2 (2)
(see Box 1 for mathematical details). This characterizes the selection of selfing under stochastic pollination. Interestingly, f(R) embodies the feedback on selfing, modulating the spread of selfing genes in stochastic environments. f(R)is always positive and increases with R. So contrary to the environmentally stable situation, selection in this case depends on the population selfing rate R, and consequently the selection gradient may vanish for intermediate selfing rates in other words, stochastic pollination may create negative feedback on selfing, which in turn allows for the maintenance of mixed mating (Fig. 3). This result may be likened to
bet-hedging theory, which states that a good strategy is the one that maximises the long-term fitness (geometric mean). Note that a qualitatively similar result may sometimes be found when inbreeding depression fluctuates in time (see Cheptou and Schoen 2002 for a theoretical model, and Dole and Ritland 1993 for an empirical example).
From the point of view of reproductive assurance, it may be tempting to consider that a population adapted to a stochastic pollination environment is the one that maximises seed production over years; i.e. the best strategy is a mix of outcrossing for good years and selfing bad pollination years (bet hedging). As shown by Fig. 4, it is not the case. This model reveals that evolutionary stable strategies do not maximize seed production, as seen in other examples from evolu-tionary biology (Maynard Smith 1982). On this point, it is important to note that Darwin’s and Baker’s verbal interpretations are not accurate since they are implicitly based on the maximisation of seed produc-tion (Ashman et al. 2004, Burd 1995). Indeed, the selection of selfing is driven by gene transmission for which seed production is only one component of the fitness. The practical consequence is that mere observation of a population that appears to be employing selfing for optimising seed production in the face of an uncertain pollination does not constitute
Box 1. The evolution of self-fertilisation under stochastic pollination environments.
The fitness at time t of a mutant, with selfing rate r in a resident population with selfing rate R in stochastic pollination environment, is given by the following equation:
Wt(r;R)r(1d) 1r 2 1R 2 et (1)
The long-term exponential growth rate is given by:
s(r;R)E[Lnw(r;R)] (2)
We assume that the population is demographically stable i.e. s(R;R)E[Lnw(R;R)]and we approximate s(r,R) using Jensen approximationE[Ln(w(r;R)]Ln[E(w(r;R)]Var(w(r;R)
2E(w(r;R)):Then, the direction of selection is given by the sign 1s
1revaluated at r/R i.e. the sign of the quantity (where e¯ ands
2are
mean and variance of the temporal pollination efficiency):
D(R) 1e=2d e(1R)R(1d) 1R2(1d 2 )dR(1:51:5d) [e(1R)R(1d)]3 s 2 A(1e=2d)Bs2 With A/0 and B/0
The evolutionary stable strategy R* is given by solving D(R)/0. The outcome of selection is summarised in Fig. 3.
a proof of adaptation to its environment. As we have shown (Fig. 4), a population producing fewer seeds may actually be closer to the evolutionary stable selfing rate.
In summary, this model reveals several interesting features. First, it combines a classical ecological scenario with basic population genetics models, which allows for the maintenance of mixed selfing rates. Second, it explicitly demonstrates that the criterion of seed maximisation under stochastic environment is not accurate from an evolutionary point of view.
Though it is true that: (1) selfing increases ovule success when seed production is pollen-limited; and (2) selfing exhibits an intrinsic advantage in pollen
trans-mission, we can see that the outcome is not trivial under general ecological scenarios.
Conclusions and perspectives
Our aim has been to integrate concepts of population genetics and concepts of the ecological theory of mating system evolution. The demographical view of ecologists has often led to the use the ‘‘optimization criteria’’ applied on seed production, whereas popula-tion geneticists have based their evolupopula-tionary analysis on the spread of a gene increasing the selfing rate, with the focus on conditions for its spread and invasion. Ecologists have explored various ecological scenarios with little theoretical formalisation, whereas population geneticists have mostly formalised gene transmission process within populations. On the one hand, population genetics have developed an impor-tant theoretical framework, but on the other hand the demographical theory allows the extension of evolu-tionary predictions to complex ecological scenarios. According to us, since models that do not integrate environmental components may not be relevant in the light of recent studies, future models in the field of mating systems ought to take these components into account.
Acknowledgements This article was initially entitled ‘‘The interplay of genes and individuals in plant mating systems evolution (or can population genetics overlook ecology much longer?). Several reviewers have pointed out the provocative nature of the title. We thank them for their clear-sightedness. We are deeply indebted to Kent Holsinger for his encourage-ments and helpful comencourage-ments. We thank Jeremiah Busch and numerous anonymous reviewers for improving the various versions of this manuscript. This article is part of the ACI project (P.-O. Cheptou) funded by the French Ministry of Research.
References
Anderson, B. et al. 2003. Facilitated selfing offers reproductive assurance: a mutualistism between a hemipteran and carnivorous plant. Am. J. Bot. 90: 10091015. Armbruster, P. and Reed, D. 2005. Inbreeding depression in
benign an stressfull environments. Heredity 95: 235 242.
Ashman, T. L. et al. 2004. Pollen limitatio n of plant reproduction: ecological and evolutionary causes and consequences. Ecology 85: 24082421.
Baker, H. G. 1955. Self-compatibility and establishment after ‘‘long distance’’ dispersal. Evolution 9: 347348. Baker, H. G. 1967. Support for Baker’s law as a rule.
Evolution 21: 853856. 0.2 0.4 0.6 0.8 1Selfing rate
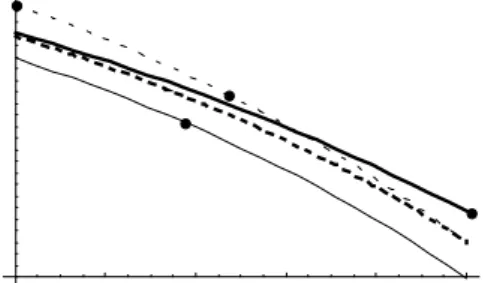
Seed production (geometr
ic mean)
Fig. 4. Geometric mean of seed production (approximation for E[ln l]) as a function of selfing rate under stochastic pollination for various sets of parameters. For each set of parameters, the evolutionary stable selfing rate is represented by a black dot. 1-continuous thin line: d/0.7; mu/0.7;
var/0.1. 2-dotted thin line:d/0.65; mu/0.85; var/0.2.
3-continuous thick line: d/0.6; mu/0.8; var/0.1.
4-dotted thick line:d/0.65; mu/0.8; var/0.08.
0.2 0.4 0.6 0.8 1 0.05 0.1 0.15 0.2 µ σ 2 Complete selfing Complete outcrossing Mixed selfing
Fig. 3. Selection of self-fertilisation in an environment with fluctuating pollination, as a function of mean and variance of pollination efficiency. In this example, inbreeding depression is fixed tod/0.6.
Barrett, S. C. H. and Harder, L. D. 1996. Ecology and evolution of plant mating system. Trends Ecol. Evol. 11: 7379.
Bijlsma, R. et al. 1999. Environmental dependence of inbreeding depression and purging inDrosophila melano-gaster. J. Evol. Biol. 12: 11251137.
Brommer, J. E. 2000. The evolution of fitness in life-history theory. Biol. Rev. 75: 377404.
Burd, M. 1995. Ovule packaging in stochastic pollination and fertilization environments. Evolution 49: 100109. Byers, D. L. and Waller, D. M. 1999. Do plant populations
purge their genetic load? Effects of population size and mating system history on inbreeding depression.Annu. Rev. Ecol. Syst. 30: 479513.
Caswell, H. 2001. Matrix population models: construction, analysis and interpretation. Sinauer.
Charlesworth B. 1994. Evolution in age-structured popula-tions (2nd ed.). Cambridge Univ. Press.
Charlesworth, B. et al. 1991. Multilocus models of inbreeding depression with synergistic selection and partial self-fertilization. Genet. Res. Camb. 57: 177194. Charlesworth, D. and Charlesworth, B. 1987. Inbreeding
depression and its evolutionary consequences. Annu. Rev. Ecol. Syst. 18: 237268.
Charlesworth, D. and Charlesworth, B. 1990. Inbreeding depression with heterozygote advantage and its effect on selection for modifiers changing the outcrossing rate.
Evolution 44: 870888.
Charlesworth, D. et al. 1990. Inbreeding depression, genetic load, and the evolution of outcrossing rates in a multilocus system with no linkage. Evolution 44: 14691489. Cheptou, P. O. and Dieckmann, U. 2002. The evolution of
self-fertilization in density-regulated populations.Proc. R. Soc. Lond. B 269: 11771186.
Cheptou, P. O. and Schoen, D. J. 2002. The cost of fluctuating inbreeding depression. Evolution 56: 10591062.
Cheptou, P. O. and Schoen, D. J. 2003. Frequency-dependent inbreeding depression in Amsinckia. Am. Nat. 162: 744753.
Darwin, C. R. 1876. The effects of cross and self fertilization in the vegetable kingdom. Murray, London.
Dole, J. and Ritland, K. 1993. Inbreeding depression in two Mimulus taxa measured by multigenerationnal changes in inbreeding coefficient. Evolution 47: 361373. Fisher, R. A. 1941. Average excess and average effect of a gene
substitution. Ann. Eugen. 11: 5363.
Goodwillie, C. et al. 2005. The evolutionary enigma of mixed mating systems in plants: occurrence, theoretical explana-tions, and empirical evidence. Annu. Rev. Ecol. Syst. 36: 4779.
Heino, M. et al. 1998. The enigma of frequency-dependent selection. Trends Ecol. Evol. 13: 367370.
Herlihy, C. R. and Eckert, C. G. 2002. Genetic cost of reproductive assurance in a self-fertilizing plants. Nature 416: 320323.
Herrera, C. M. et al. 2001. Geographical variation in autonomous self-pollination levels unrelated to pollinator service inHelleborus foetidus (Ranunculaceae). Am. J. Bot. 88: 10251032.
Holsinger, K. E. 1988. Inbreeding doesn’t matter: the genetic basis of mating system evolution.Evolution 42: 1235 1244.
Holsinger, K. E. 1991a. Inbreeding depression and the evolution of plant mating systems. Trends Ecol. Evol. 6: 307308.
Holsinger, K. E. 1991b. Mass action models of plant mating systems: the evolutionnary stability of mixed mating systems. Am. Nat. 138: 606622.
Holsinger, K. E. 1996. Pollination biology and the evolution of mating systems in flowering plant. Evol. Biol. 29: 107149.
Holsinger, K. E. 2000. Reproductive systems and evolution in vascular plants. Proc. Natl Acad. Sci. USA 97: 7037 7042.
Husband, B. and Schemske, D. W. 1996. Evolution of the magnitude and timing of inbreeding depression in plants. Evolution 50: 5470.
Jain, S. K. 1976. The evolution of inbreeding in plants. Annu. Rev. Ecol. Syst. 7: 469495.
Kalisz, S. and Vogler, D. W. 2003. Benefits of autonomous selfing under unpredictable pollinator environments.
Ecology 84: 29282942.
Klips, R. A. and Snow, A. A. 1997. Delayed autonomous self-pollination inHibiscus laevis (Malvaceae). Am. J. Bot. 84: 4853.
Knight, T. 1799. Experiments on the fecundation of vegetables. Philos. Trans. R. Soc. Lond. 89: 195 204.
Lande, R. 1982. A quantitative genetic theory of life history evolution. Ecology 63: 607615.
Lande, R. and Schemske, D. W. 1985. The evolution of self fertilization and inbreeding depression in plants. I. Genetic models. Evolution 39: 2440.
Lloyd, D. G. 1977. Genetic and phenotypic models of natural selection. J. Theor. Biol. 69: 543560.
Lloyd, D. G. 1979. Some reproductive factors affecting the selection of self-fertilization in plants. Am. Nat. 113: 6779.
Lloyd, D. G. 1980. Demographic factors and mating patterns in Angiosperms. In: Solbrig, O. T. (ed.), Demography and evolution in plant populations. Univ. California Press, pp. 6788.
Maynard Smith, J. 1982. Evolution and the theory of games. Cambridge Univ. Press.
Metz, J. A. J. et al. 1992. How should we define fitness for general ecological scenarios? Trends Ecol. Evol. 7: 198202.
Morgan, M. T. and Wilson, W. G. 2005. Self-fertilization and the escape from pollen limitation in variable pollina-tion environments. Evolution 59: 11431148. Pannell, J. R. and Barrett, C. H. 1998. Bakers’law:
repro-ductive assurance in a metapopulation. Evolution 52: 657668.
Ronfort, J. and Couvet, D. 1995. A stochastic model of selection on the selfing rates in structured populations.
Genet. Res. 65: 209222.
Schmitt, J. and Ehrhardt, D. W. 1990. Enhancement of inbreeding depression by dominance and suppression in Impatiens capensis. Evolution 44: 269278.
Schueller, S. K. 2004. Self-pollination in island and mainland populations of the introduced hummingbird-pollinated plant, Nicotiana glauca (Solanaceae). Am. J. Bot 91: 672681.
Stebbins, G. L. 1957. Self-fertilization and population variability in higher plants. Am. Midl. Nat. 91: 337 354.
Uyenoyama, M. K. 1986. Inbreeding and the cost of meiosis: the evolution of selfing in populations practcing bipar-ental inbreeding. Evolution 40: 344388.
Uyenoyama, M. K. et al. 1993. Ecological and genetic factors directing the evolution of self fertilization. Oxford Survey Evol. Biol. 9: 327381.
Vogler, D. W. and Kalisz, S. 2001. Sex among the flowers: the distribution of plant mating systems. Evolution 55: 202204.
Ward, M. et al. 2005. To self, or not to self...a review of outcrossing and pollen-mediated gene flow in neotropical trees. Heredity 95: 246254.
Yampolsky, C. and Yampolsky, H. 1922. Distribution of sex forms in the phanerogamic flora.Bibl. Genet. 3: 162.