Copyright © 1998, American Society for Microbiology
Clinical Application of PCR-Restriction Enzyme Pattern Analysis for
Rapid Identification of Aerobic Actinomycete Isolates
REBECCA W. WILSON,
1VINCENT A. STEINGRUBE,
2* BARBARA A. BROWN,
2AND
RICHARD J. WALLACE, JR.
1,2Center for Pulmonary and Infectious Disease Control
1and Department of Microbiology,
2The University of Texas Health Center at Tyler, Tyler, Texas
Received 9 July 1997/Returned for modification 23 September 1997/Accepted 17 October 1997
The accuracy and practicality of PCR-restriction enzyme pattern analysis (PRA) for routine identification of
aerobic actinomycete clinical isolates were evaluated for 299 cultures submitted to the Mycobacteria/Nocardia
Laboratory at the University of Texas Health Center at Tyler. PRA identification using an amplified 439-bp
segment (amplicon) of the 65-kDa heat shock protein gene was compared to identification by traditional
methods, including growth characteristics, susceptibility patterns, biochemical testing, and high-performance
liquid chromatography analysis. Microbiological examination of six cultures ruled out aerobic actinomycetes,
and they were omitted from the study. Amplicons were analyzed with BstEII, HaeIII, MspI, HinfI, and BsaHI.
When necessary, AciI, HhaI, and NarI were also used. From March 1995 through May 1997 (27 months), 274
of the remaining 293 (93.5%) isolates were accurately identified by PRA. Major diagnostic groups included 170
mycobacteria, 93 nocardiae, and 30 other aerobic actinomycetes. Mixed cultures were readily recognized by
PRA, including a wound culture that contained two Nocardia taxa that were indistinguishable morphologically.
Mycobacterium mucogenicum was identified in three cultures heavily contaminated with gram-positive cocci.
The 19 isolates that produced PRA patterns that did not match those in the current PRA database were
differentiated into 8 Mycobacterium species and 11 other aerobic actinomycetes by the presence or absence of
BstEII recognition sites. Identification of 15 of these 19 isolates was also equivocal by traditional methods. PRA
results were reportable within 2 to 5 working days and were as accurate as and faster and less expensive to
obtain than those of traditional methods.
Interest in the identification and taxonomy of aerobic
acti-nomycetes, nocardiae and mycobacteria in particular, has been
increasing as a result of the increasing number of
immunocom-promised individuals in the population who are at greater risk
for actinomycoses, especially those with advanced human
im-munodeficiency virus disease (1, 4, 5). Traditional methods for
differentiation of species and taxa of aerobic actinomycetes are
laborious and time-consuming and frequently require
special-ized testing that is beyond the capabilities of clinical
laborato-ries (2–5, 9, 11, 19, 21). The occurrence of clinical isolates of
aerobic actinomycetes that are inherently resistant to specific
antimicrobials increases the significance of timely and accurate
species and taxon recognition (4, 9, 17, 22).
Successful application of molecular biological methodology
to the development of protocols for rapid differentiation of
mycobacterial species was demonstrated by Telenti et al. in
1993 (13). These authors used PCR-restriction enzyme pattern
analysis (PRA) of an amplified 439-bp segment of the 65-kDa
heat shock protein (hsp-65) gene and introduced the
abbrevi-ation PRA for this method that has now gained wide
accep-tance (16). Application of this methodology has since been
expanded to include 50 commonly encountered pathogenic
species and taxa of aerobic actinomycetes comprising the
gen-era Mycobacterium (10, 13); Nocardia (9, 17, 22); and
Actino-madura, Gordona, Rhodococcus, Streptomyces, and
Tsukamu-rella (11). The rapidity and accuracy of PRA prompted the
current study (9–11, 13, 17). Clinical isolates of aerobic
actino-mycetes submitted to the Mycobacteria/Nocardia Laboratory
at the University of Texas Health Center at Tyler (UTHCT)
for identification and susceptibility testing were subjected to
PRA for identification in an effort to evaluate the efficacy and
cost-effectiveness of this methodology for routine clinical use.
(This study was presented in part at the 97th General
Meet-ing of the American Society for Microbiology, Miami Beach,
Fla., 1997.)
MATERIALS AND METHODS
Organisms.The present study included 293 clinical isolates of aerobic actino-mycetes submitted to the Mycobacteria/Nocardia Research Laboratory at the UTHCT for identification and susceptibility testing during the 27-month period from March 1995 through May 1997. All clinical isolates used in this study were subcultured onto Trypticase soy and Middlebrook 7H10 agar plates. One culture of each isolate was used for identification by colonial morphology and antimi-crobial susceptibility patterns (1, 17–21). Selected biochemical testing was done in order to differentiate species or taxa with similar susceptibility patterns (6, 10, 14, 15, 17, 19, 21). High-performance liquid chromatography (HPLC) and addi-tional biochemical testing were kindly performed by the Bureau of Laboratories, Texas State Health Department (Austin).
The ATCC type strains of Mycobacterium fortuitum (ATCC 6841) and
Nocar-dia brasiliensis (ATCC 19296) were utilized as internal controls for PRA.
PCR amplification.DNA was prepared from cells harvested from the initially submitted agar slants and/or the second subculture, when necessary, according to methods previously described (9–11, 13). A 439-bp segment of the hsp-65 gene was amplified from ground cell supernatants by PCR with 1.0 U of Taq DNA polymerase (Boehringer Mannheim, Indianapolis, Ind.) in optimized buffer E
(1.5 mM MgCl2[pH 9.0]; Invitrogen, San Diego, Calif.) containing 83mM (each)
deoxynucleoside triphosphates, 9% dimethyl sulfoxide, and 1mM (each) primers
TB11 (59-ACCAACGATGGTGTGTCCAT) and TB12 (59-CTTGTCGAACCG
CATACCCT) (Midland Certified Reagent Co., Midland, Tex.), together with the appropriate positive and negative controls according to a modification of the method of Telenti et al. (13). The PCR mixtures were run for 45 cycles of 94, 55, and 72°C for 1 min each and then for a 10-min extension period at 72°C.
Restriction enzyme analysis.Data from previous studies (9–11, 13) resulted in the selection of five commercially available restriction endonucleases, BstEII,
HaeIII, MspI, HinfI, and BsaHI (New England Biolabs, Beverly, Mass., and
Promega, Madison, Wis.), for routine use in the production of PRA band pat-terns. When indicated (references 10 and 22 and unpublished data), one or more
* Corresponding author. Mailing address: Department of
Microbi-ology, The University of Texas Health Center at Tyler, P.O. Box 2003,
Tyler, TX 75710-2003. Phone: (903) 877-7685. Fax: (903) 877-7652.
E-mail: [email protected].
148
on May 15, 2020 by guest
http://jcm.asm.org/
of a secondary set of endonucleases that included AciI, HhaI, and NarI was utilized. Restriction digests were incubated for the appropriate time periods, at the appropriate temperatures, and with the buffers recommended by the man-ufacturers, with the exception of the temperature and digest mixture for BsaHI. To achieve complete digestion with BsaHI, acetylated bovine serum albumin was substituted for bovine serum albumin and the digestion mixture was incubated at 60°C for 1 h.
Restriction fragments were electrophoresed on 3% Metaphor agarose (4-bp resolution; FMC Bioproducts, Rockland, Maine), containing ethidium bromide
(0.625mg/ml), in a Mini-Sub-Cell electrophoresis system (Bio-Rad Laboratories,
Richmond, Calif.) at 95 V for 1.5 to 2.0 h.
Isolate identification. PRA band sizes (base pairs) from each isolate were estimated visually by comparison with a 100-bp ladder (Life Technologies, Grand Island, N.Y.), a pGEM base pair ladder (Promega), and the PRA patterns obtained for control strains on each gel. Each isolate was then initially identified by one member of the staff (R.W.W.) to the species or taxon level by comparison of visually estimated PRA band sizes with those of species- and taxon-specific patterns contained in the PRA database (9–11, 13, 17, 22). Visual PRA isolate identifications were made prior to, and without knowledge of, identification results by traditional methods.
The PRA database was developed primarily for clinically significant nonpig-mented rapidly growing mycobacteria, nocardiae, and other clinically significant aerobic actinomycetes (9–11, 13). With the exception of Mycobacterium avium and Mycobacterium intracellulare, for which 83 and 129 isolates, respectively, have been studied by PRA, the entries in the database for slow-growing mycobacteria have been less extensively developed. This portion of the database (unpublished data) represented PRA patterns obtained from 2 to 10 isolates of each slow-growing mycobacterial species most commonly encountered in clinical samples, including Mycobacterium celatum, Mycobacterium kansasii, Mycobacterium
[image:2.612.300.546.79.602.2]scrofulaceum, and Mycobacterium triviale, in addition to the 10 species listed in
Table 1. These patterns were very similar to those published by Telenti et al. (13). To date, the PRA database has not been expanded to include pigmented rapidly growing mycobacterial species owing to their predominantly environmental or-igin and infrequent clinical occurrence as agents of traumatic wound infections. Species or genus (e.g., Streptomyces) identification was considered conclusive when the PRA pattern matched that of a single species or taxon of aerobic actinomycete in the PRA database and the resulting identification was in agree-ment with that based on traditional methods.
To test whether precise PRA band size measurements were required for accurate isolate identification, band sizes were measured on a computerized Bio Image system (Millipore, Bedford, Mass.) with the same molecular size stan-dards as noted above, and isolate identifications were made independently by a different member of the staff (V.A.S.) without prior knowledge of the initial visual identifications (R.W.W.). The two independent PRA identifications were then compared to one another and with identifications based on traditional methods including growth characteristics, susceptibility patterns, biochemical tests, and HPLC analysis.
Time studies of the PRA identification protocol were performed in order to provide an estimate of the amount of time and labor involved in applying this methodology under routine clinical conditions.
RESULTS
Organisms.
On microbiological evaluation, six of the
cul-tures submitted to the UTHCT laboratory for identification
and susceptibility testing were not aerobic actinomycetes.
These cultures did not yield PCR amplification products
(am-plicons) and were excluded from further study. The
distribu-tion of species and taxa identified among the remaining 293
isolates is listed in Table 1. The seven predominant species of
aerobic actinomycetes identified in this study comprised over
half, 174 of 293 (59%), of the isolates submitted and included
the following: Mycobacterium abscessus, 47 of 293 (16%)
iso-lates; M. fortuitum, 32 of 293 (11%) isoiso-lates; Nocardia nova, 24
of 293 (8%) isolates; Mycobacterium mucogenicum (formerly
Mycobacterium chelonae-like organism) (8), 20 of 293 (7%)
isolates; M. chelonae, 18 of 293 (6%) isolates; Nocardia
aster-oides type VI (20), 17 of 293 (6%) isolates; and N. brasiliensis,
16 of 293 (5%) isolates.
PRA identification.
As shown in Table 1, 274 of the 293
(93.5%) isolates produced PRA patterns that matched
species-or taxon-specific patterns in the PRA database and resulted in
an identification that correlated with the identification by
tra-ditional methods. The remaining 19 isolates produced PRA
patterns that did not match any of the patterns currently
avail-able in the database. Eight of these isolates produced
ampli-cons that contained BstEII recognition sites and were
there-fore tentatively identified as Mycobacterium species (9–11).
Only 4 of these 19 isolates were unequivocally identified to the
species level by traditional methods, as shown in Table 2. One
isolate (Mo 816) produced an amplicon lacking BstEII
recog-nition sites and an HaeIII pattern resembling that published
TABLE 1. Clinical isolates identified by PRA in this study
Sp. and taxon No. of clinicalisolates
Genus Mycobacterium Rapidly growing spp.
M. abscessus ... 47
M. abscessus-M. chelonae complex... 4
M. chelonae ... 18
M. fortuitum ... 32
M. fortuitum, third biovariant complex... 5
M. fortuitum, third biovar, sorbitol-positive– M. peregrinum Piprcomplex... 1
M. mucogenicum (MCLO)a... 20
M. smegmatis... 10
Slow-growing spp. M. avium... 3
M. gordonae... 1
M. haemophilum... 2
M. intracellulare ... 4
M. marinum... 7
M. simiae ... 2
M. szulgai... 1
M. terrae-M. nonchromogenicum complex... 2
M. tuberculosis ... 3
Mycobacterium sp. (BstEII sites present; taxon not distinguishable)... 8
Mycobacterium subtotal ... 170
Genus Nocardia Nocardia asteroides complex Type Ib... 9
Type II... 1
Type VI ... 17
Taxon not further differentiated ... 1
N. brasiliensis ... 16
N. farcinica... 11
N. nova ... 24
N. otitidiscaviarum... 3
N. pseudobrasiliensisc... 2
N. transvalensis complex... 9
Nocardia subtotal ... 93
Genera of other aerobic actinomycetes Actinomadura madurae... 1
Gordona sputi ... 2
Gordona sp... 1
Rhodococcus equi ... 1
Streptomyces sp. ... 10
Tsukamurella paurometabolum ... 4
Other aerobic actinomycete taxa (BstEII sites absent; taxon not distinguishable) ... 11
Actinomycete subtotal ... 30
Total clinical isolates ... 293
aDescribed by Springer et al. (8). MCLO, M. chelonae-like organism.
bAntibiogram types of N. asteroides complex were described by Wallace et al.
(20).
cDescribed by Ruimy et al. (7) and Wallace et al. (17).
on May 15, 2020 by guest
http://jcm.asm.org/
[image:2.612.307.541.359.599.2]for Mycobacterium vaccae (13) but was not identified as such
due to the lack of adequate data in the PRA database at the
time that the isolate was received. A second unique pattern
was observed for isolate N 1426, which was identified as
No-cardia sp., most likely N. asteroides complex, and which
pro-duced an amplicon containing BstEII recognition sites. This
sputum isolate gave unique PRA patterns with all enzymes
tested and was the first and only Nocardia isolate, among 210
Nocardia isolates examined by PRA in this laboratory, that
demonstrated BstEII recognition sites. Traditional methods
unequivocally identified 278 of the 293 (94.9%) clinical isolates
studied, while PRA correctly identified 274 isolates, resulting
in a comparative accuracy of 98.6%.
Three cultures that contained small numbers of
gram-posi-tive rods mixed with heavy overgrowth of gram-posigram-posi-tive cocci
were submitted. Direct PRA was carried out on cells taken
from the mixed cultures submitted, and in all three cases, the
amplicons obtained produced patterns that were typical of M.
mucogenicum. This identification was later confirmed by
tradi-tional microbiological methods. Two additradi-tional cultures that
appeared to be pure produced an excessive number of bands
on PRA gels that gave total base pair values in excess of the
expected 439 bp. One of these cultures was identified at a
ref-erence laboratory as containing both Corynebacterium
aqua-ticum and Actinomyces viscosus (Table 2). The second mixed
culture from a multiply infected wound site yielded two distinct
PRA patterns that, when reanalyzed on individual colony
picks, were typical for isolates of the N. asteroides complex
antibiogram type I (20) and the Nocardia transvalensis new
taxon 2 of the proposed N. transvalensis complex (9, 22).
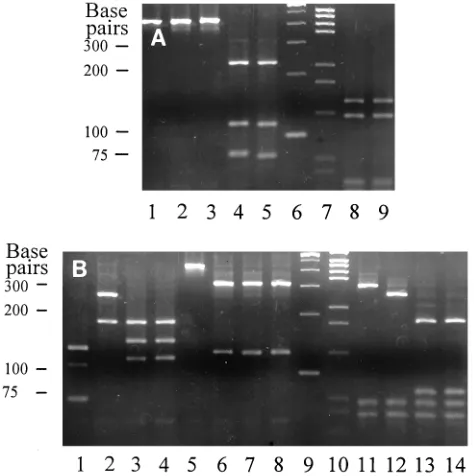
Comparison of visual PRA band size estimates and isolate
identifications from gel photographs (Fig. 1) with independent
isolate identifications based on computer-generated
measure-ments of PRA band sizes resulted in 100% agreement between
the two identification methods. Comparison of measured PRA
band size (base pair) values from Fig. 1 with published values
for the matching species and taxon (9–11) corroborated the
visual estimations of PRA band sizes and the identification of
the clinical isolates. For example, clinical isolate 96–113 gave
PRA band patterns of 235, 115, and 80 bp with BstEII (Fig. 1A,
lane 4) and 145 and 125 bp with HaeIII (Fig. 1A, lane 8), which
matched the published PRA patterns of M. fortuitum (10).
These patterns also matched those of the control strain (ATCC
6841) of M. fortuitum shown in Fig. 1A, lanes 5 and 9,
respec-tively. Likewise, clinical isolates 96–110, 96–111, and 96–112
were identified by matching their PRA band patterns from
BstEII, MspI, HinfI, and BsaHI digests with those published (9,
11) for N. nova, N. asteroides complex antibiogram type I, and
N. brasiliensis, respectively.
[image:3.612.50.289.87.380.2]A time course study was performed with the PRA procedure
in order to better define the amount of time and labor involved
and to provide a stepwise description of the procedure (Table
3). Final identification results can be achieved within 24 h of
FIG. 1. PRA patterns from BstEII, HaeIII, MspI, HinfI, and BsaHI digests of amplicons from clinical isolates and reference strains of aerobic actinomycetes. (A) Lanes 1 to 5, BstEII digests; lanes 8 and 9, HaeIII digests. Amplicon digests: lanes 1 to 5, clinical isolates 96–110, 96–111, 96–112, and 96–113 and reference strain ATCC 6841, respectively; lanes 6 and 7, size markers (100-bp and pGEM-bp ladders, respectively); lanes 8 and 9, clinical isolate 96–113 and ref-erence strain ATCC 6841, respectively. (B) Lanes 1 to 4, MspI digests; lanes 5 to 8, HinfI digests; lanes 11 to 14, BsaHI digests. Amplicon digests: lanes 1 to 4, clinical isolates 96–110, 96–111, and 96–112 and reference strain ATCC 19296, respectively; lanes 5 to 8, clinical isolates 96–110, 96–111, and 96–112 and reference strain ATCC 19296, respectively; lanes 9 and 10, size markers (100-bp and pGEM-bp ladders, respectively); lanes 11 to 14, clinical isolates 96–110, 96–111, and 96–112 and reference strain ATCC 19296, respectively. Clinical isolates 96–110, 96–111, 96–112, and 96–113 were identified as N. nova, N.
asteroides complex antibiogram type I (20), N. brasiliensis, and M. fortuitum,
[image:3.612.308.545.350.588.2]respectively, by matching each of their PRA band patterns with published values (9–11).
TABLE 2. Identification of clinical isolates that exhibited patterns
that did not match those in the current PRA database
Isolate no. Identificationa
Isolates distinguished as
Myco-bacterium sp. based on
the presence of BstEII
recognition sitesb,c
Mo 702 ...Mycobacterium sp. (pigmented rapid grower) Mo 748 ...Mycobacterium sp. (pigmented rapid grower) N 1426 ...Nocardia sp.
Mo 783 ...M. simiae
Mo 821 ...Mycobacterium sp. (pigmented rapid grower) Mo 830 ...Mycobacterium sp. (slow grower)
Mo 855 ...Mycobacterium sp. (pigmented rapid grower) Mo 875 ...Mycobacterium sp. (slow grower)
Isolates distinguished as aero-bic actinomycetes, other than Mycobacterium sp., based on the absence of
BstEII recognition sitesb,d
Mo 728#1 ...Corynebacterium aquaticum Mo 728#2 ...Actinomyces viscosus
N 1420 ...Unidentifiable; possible N. transvalensis Mo 780 ...Rhodococcus sp.
Mo 791 ...Gordona sp. Mo 798 ...N. asteroides complex Mo 802 ...Tsukamurella sp. As 121...Streptomyces sp. As 124...N. asteroides complex Mo 816 ...M. vaccae
Mo 820 ...Unidentifiable; possible Mycobacterium sp. (slow grower)
aIdentification by biochemical tests, antimicrobial susceptibility patterns, and
HPLC analysis.
bWork of Steingrube et al. (9, 11).
cTotal of 8.
dTotal of 11.
on May 15, 2020 by guest
http://jcm.asm.org/
receiving a culture under optimal conditions. As a routine
practice, however, separate analysis of individual isolates was
neither cost-effective nor practical. As a practical routine,
iden-tification results based on PRA were generally achievable on a
2- to 5-day schedule.
DISCUSSION
PRA correctly identified 274 of 293 (93.5%) aerobic
actino-mycete clinical isolates, compared to traditional identification
methods that unequivocally identified 278 of those isolates
(94.9%). Comparison of the two identification methods
re-sulted in a 98.6% relative accuracy for the molecular biological
identification method. PRA proved highly specific, enabling
identification of aerobic actinomycetes from cultures heavily
contaminated with other bacteria as well as identifying mixed
cultures that contained more than one taxon of aerobic
acti-nomycete that was not readily recognizable on isolation plates.
The specificity of PRA was further demonstrated with six
cul-tures that failed to yield amplicons on PCR and were found not
to contain aerobic actinomycetes on further microbiological
evaluation.
Identification of species and taxa of aerobic actinomycete
isolates commonly encountered in clinical specimens was
ac-complished with virtually 100% accuracy. Only very rarely
oc-curring species presented difficulties for identification, most
frequently as a result of the unavailability of reference patterns
in the PRA database. As noted above, the current PRA
data-base has been well developed for species and taxa of
nonpig-mented rapidly growing mycobacteria, nocardiae, and other
clinically significant aerobic actinomycetes (9–11). PRA data
have not been as well developed for species of slow-growing
mycobacteria, and to date, data from PRA of pigmented
rap-idly growing mycobacteria remain to be developed. As this
study has progressed over the past 2 years, data for less
fre-quently occurring species have been constantly added to the
database as sufficient numbers of isolates and reference strains
have been accumulated and examined by PRA. This steady
development of the scope and breadth of the PRA database
should continually expand the number of aerobic actinomycete
species identifiable by this method.
Seven categories listed in Table 1 appear as complex or
genus identifications and warrant further consideration. The
M. abscessus-M. chelonae complex; M. fortuitum third
biovari-ant complex; M. fortuitum third biovar,
sorbitol-positive–My-cobacterium peregrinum Pip
rcomplex (10); and the
Mycobac-terium terrae-MycobacMycobac-terium nonchromogenicum complex each
represent two groups containing very closely related
mycobac-terial taxa that are not readily differentiated by either
tradi-tional methods or PRA (10, 13). Three isolates listed as M.
abscessus-M. chelonae complex could not be differentiated
fur-ther by traditional methods and exhibited hybrid PRA patterns
that contained features of both M. abscessus and M. chelonae.
These isolates may represent a heretofore-unrecognized taxon
closely related to these mycobacterial species. The isolate
listed as an undifferentiated taxon of the N. asteroides complex
gave patterns with MspI and BsaHI that resembled those of the
N. asteroides complex antibiogram type I (20) but produced a
pattern with HinfI that was not represented in the PRA
data-base and, therefore, may represent a minor pattern for isolates
within this taxon, similar to the occurrence of major and minor
patterns previously observed for isolates of this taxon with
BsaHI (9). The single isolate listed as Gordona sp. exhibited a
PRA pattern with HinfI that was unique to this genus but gave
patterns with all other endonucleases tested that matched
those of the previously reported isolate Mo 315 (11). These
two isolates could be identified only to the genus level at the
Centers for Disease Control and Prevention (Atlanta, Ga.) and
were neither Gordona bronchialis nor Gordona sputi. As
pre-viously discussed (11), identification of Streptomyces isolates
beyond the genus level was considered to be clinically
irrele-vant and was not pursued in this study.
Identification of clinical isolates of aerobic actinomycetes by
PRA, performed on a continuing daily basis common to the
routine of clinical laboratories, provided final identification
results within 2 to 5 working days, compared to traditional
identification methods that required from 2 to 6 weeks for final
results. Prompt and accurate identification of pathogenic
aer-obic actinomycete isolates is particularly important when
inva-sive species such as Nocardia pseudobrasiliensis (7, 17) are
encountered or when innately drug-resistant species such as
those of the N. transvalensis complex, which are resistant to all
aminoglycosides (9, 20, 22), and Nocardia farcinica, which is
resistant to all extended-spectrum cephalosporins (12, 21), are
involved. This is particularly significant when
immunocompro-mised patients such as those with advanced human
immuno-deficiency virus disease are infected with these organisms (4, 5,
9, 17, 22).
Although absolute PRA band sizes have been found to vary
from 5 to 10 bp between laboratories (10, 13), the overall
patterns have proven highly reproducible and species- and
taxon-specific. Visual comparison of PRA patterns with
mo-lecular size standards and patterns produced by internal
con-trol isolates, such as M. fortuitum and N. brasiliensis reference
strains used in this study, resulted in successful clinical isolate
identifications that correlated perfectly with identifications
based on measured band size values. Consequently, there is no
requirement for costly computerized measurement systems, a
major expense consideration, in implementing this
methodol-ogy for routine clinical use.
[image:4.612.51.289.91.263.2]PRA was cost-effective, with the expenses of specialized
equipment and reagents being more than compensated for by
savings in time and labor, and could be economically
incorpo-rated into the clinical laboratory setting. This methodology has
proven both practical and cost-effective as a rapid, efficient,
and highly accurate identification system for use in identifying
clinically significant species and taxa of aerobic actinomycetes.
TABLE 3. Timed protocol for aerobic actinomycete
clinical isolate identification by PRA
aPRA
step no. PRA procedure required (h)Time
1 Cell harvest, suspension in buffer, heat kill, grind
in Mini-Beadbeater 1.5
2 Set up PCRs and program Temp-cycler 1
3 PCR amplification in Temp-cycler with 4°C
hold-ing mode Overnight
4 Preparation of four agarose gels (one for step 5
and three for step 8) 1
5 Gel loading and electrophoresis of PCR products
(amplicons) 1.75
6 Set up restriction endonuclease digests (three
restriction endonucleases and up to 36 digests) 1 7 Incubation of restriction endonuclease digests 1 8 Gel loading and electrophoresis of restriction
endonuclease digests (three gels) 2–2.5
9 Photograph gels, evaluate PRA patterns, and
identify clinical isolates 2.25
aTime required to analyze 12 clinical isolates per procedure.
on May 15, 2020 by guest
http://jcm.asm.org/
ACKNOWLEDGMENTS
This work was supported by the Department of Microbiology and
the Center for Pulmonary and Infectious Disease Control at UTHCT.
We express our appreciation to Phyllis Pienta, Collection Manager
of Bacteriology, American Type Culture Collection, Rockville, Md.,
who kindly provided the reference strains used in the development of
the PRA database; to the Bureau of Laboratories of the Texas State
Department of Health (Austin, Tex.); and to Kenneth C. Jost, Jr., for
his expertise with HPLC in assisting with the identification of clinical
isolates of aerobic actinomycetes evaluated in this study.
REFERENCES
1. Beaman, B. L., M. A. Saubolle, and R. J. Wallace, Jr. 1995. Nocardia,
Rhodococcus, Streptomyces, Oerskovia, and other aerobic actinomycetes of
medical importance, p. 379–399. In P. R. Murray, E. J. Baron, M. A. Pfaller, F. C. Tenover, and R. H. Yolken (ed.), Manual of clinical microbiology, 6th ed. ASM Press, Washington, D.C.
2. Collins, C. H., M. D. Yates, and A. H. C. Uttley. 1988. Presumptive identi-fication of nocardias in a clinical laboratory. J. Appl. Bacteriol. 65:55–59. 3. Georghiou, P. R., and Z. M. Blacklock. 1988. Infection with Nocardia species
in Queensland. A review of 102 clinical isolates. Med. J. Aust. 156:692–697. 4. McNeil, M. M., J. M. Brown, P. R. Georghiou, A. M. Allworth, and Z. M.
Blacklock.1992. Infections due to Nocardia transvalensis: clinical spectrum and antimicrobial therapy. Clin. Infect. Dis. 15:453–463.
5. McNeil, M. M., J. M. Brown, C. H. Magruder, K. T. Shearlock, R. A. Saul,
D. P. Allred, and L. Ajello.1992. Disseminated Nocardia transvalensis infec-tion: an unusual opportunistic pathogen in severely immunocompromised patients. J. Infect. Dis. 165:175–178.
6. Mishra, S. K., R. E. Gordon, and D. A. Barnett. 1980. Identification of nocardia and streptomycetes of medical importance. J. Clin. Microbiol. 11: 728–736.
7. Ruimy, R., P. Riegel, A. Carlotti, P. Boiron, G. Bernardin, H. Monteil, R. J.
Wallace, Jr., and R. Christen.1996. Nocardia pseudobrasiliensis sp. nov., a new species of nocardia which groups bacterial strains previously identified as Nocardia brasiliensis and associated with invasive diseases. Int. J. Syst. Bacteriol. 46:259–264.
8. Springer, B., E. C. Bo¨ttger, P. Kirschner, and R. J. Wallace, Jr. 1995. Phylogeny of the Mycobacterium chelonae-like organism based on partial sequencing of the 16S rRNA gene and proposal of Mycobacterium
muco-genicum sp. nov. Int. J. Syst. Bacteriol. 45:262–267.
9. Steingrube, V. A., B. A. Brown, J. L. Gibson, R. W. Wilson, J. Brown, Z.
Blacklock, K. Jost, R. F. Ulrich, and R. J. Wallace, Jr.1995. DNA amplifi-cation and restriction endonuclease analysis for differentiation of 12 species and taxa of Nocardia, including recognition of four new taxa within the N.
asteroides complex. J. Clin. Microbiol. 33:3096–3101.
10. Steingrube, V. A., J. L. Gibson, B. A. Brown, Y. Zhang, R. W. Wilson, M.
Rajagopalan, and R. J. Wallace, Jr.1995. PCR amplification and restriction endonuclease analysis of a 65-kilodalton heat shock protein gene sequence for taxonomic separation of rapidly growing mycobacteria. J. Clin. Microbiol.
33:149–153.
11. Steingrube, V. A., R. W. Wilson, B. A. Brown, K. C. Jost, Jr., Z. Blacklock,
J. L. Gibson, and R. J. Wallace, Jr.1997. Rapid identification of clinically significant species and taxa of aerobic actinomycetes, including
Actinoma-dura, Gordona, Nocardia, Rhodococcus, Streptomyces, and Tsukamurella
iso-lates, by DNA amplification and restriction endonuclease analysis. J. Clin. Microbiol. 35:817–822.
12. Steingrube, V. A., R. J. Wallace, Jr., B. A. Brown, Y. Zhang, L. C. Steele, G.
Young, and D. R. Nash.1993. Partial characterization of Nocardia farcinica
b-lactamases. Antimicrob. Agents Chemother. 37:1850–1855.
13. Telenti, A., F. Marchesi, M. Balz, F. Bally, E. C. Bo¨ttger, and T. Bodmer. 1993. Rapid identification of mycobacteria to the species level by polymerase chain reaction and restriction enzyme analysis. J. Clin. Microbiol. 31:175– 178.
14. Tsukamura, M. 1977. Extended numerical study of Nocardia. Int. J. Syst. Bacteriol. 27:311–323.
15. Tsukamura, M. 1978. Numerical classification of Rhodococcus (formerly
Gordona) organisms recently isolated from sputa of patients: description of Rhodococcus sputi Tsukamura sp. nov. Int. J. Syst. Bacteriol. 28:169–181.
16. Wallace, R. J., Jr. 1997. Presented at the 18th Annual Congress of the European Society of Mycobacteriology and the 2nd International Sympo-sium on Mycobacteria of Clinical Interest, Co´rdoba, Spain, 17 to 20 June 1997.
17. Wallace, R. J., Jr., B. A. Brown, Z. Blacklock, R. Ulrich, K. Jost, J. M. Brown,
M. M. McNeil, G. Onyi, V. A. Steingrube, and J. L. Gibson.1995. New
Nocardia taxon among isolates of Nocardia brasiliensis associated with
inva-sive disease. J. Clin. Microbiol. 33:1528–1533.
18. Wallace, R. J., Jr., and L. C. Steele. 1988. Susceptibility testing of nocardia species for the clinical laboratory. Diagn. Microbiol. Infect. Dis. 9:155–166. 19. Wallace, R. J., Jr., B. A. Brown, M. Tsukamura, J. M. Brown, and G. O.
Onyi.1991. Clinical and laboratory features of Nocardia nova. J. Clin. Mi-crobiol. 29:2407–2411.
20. Wallace, R. J., Jr., L. C. Steele, G. Sumter, and J. M. Smith. 1988. Antimi-crobial susceptibility patterns of Nocardia asteroides. Antimicrob. Agents Chemother. 32:1776–1779.
21. Wallace, R. J., Jr., M. Tsukamura, B. A. Brown, J. Brown, V. A. Steingrube,
Y. Zhang, and D. R. Nash.1990. Cefotaxime-resistant Nocardia asteroides strains are isolates of the controversial species Nocardia farcinica. J. Clin. Microbiol. 28:2726–2732.
22. Wilson, R. W., V. A. Steingrube, B. A. Brown, Z. Blacklock, K. C. Jost, Jr.,
A. McNabb, W. D. Colby, J. R. Biehle, J. L. Gibson, and R. J. Wallace, Jr.
1997. Recognition of a Nocardia transvalensis complex by resistance to ami-noglycosides, including amikacin, and PCR-restriction fragment length poly-morphism analysis. J. Clin. Microbiol. 35:2235–2242.