JOURNAL OFVIROLOGY, Oct. 2004, p. 10706–10714 Vol. 78, No. 19
0022-538X/04/$08.00⫹0 DOI: 10.1128/JVI.78.19.10706–10714.2004 Copyright © 2004, American Society for Microbiology. All Rights Reserved.
Forced Selection of a Human Immunodeficiency Virus Type 1 Variant
That Uses a Non-Self tRNA Primer for Reverse Transcription:
Involvement of Viral RNA Sequences and the Reverse
Transcriptase Enzyme
Truus E. M. Abbink, Nancy Beerens,† and Ben Berkhout*
Department of Human Retrovirology, Academic Medical Center, University of Amsterdam, Amsterdam, The Netherlands
Received 11 February 2004/Accepted 6 May 2004
Human immunodeficiency virus type 1 uses the tRNA3Lysmolecule as a selective primer for reverse
tran-scription. This primer specificity is imposed by sequence complementarity between the tRNA primer and two motifs in the viral RNA genome: the primer-binding site (PBS) and the primer activation signal (PAS). In addition, there may be specific interactions between the tRNA primer and viral proteins, such as the reverse transcriptase (RT) enzyme. We constructed viruses with mutations in the PAS and PBS that were designed to employ the nonself primer tRNAProor tRNA
1,2
Lys. These mutants exhibited a severe replication defect, indicating
that additional adaptation of the mutant virus is required to accommodate the new tRNA primer. Multiple independent virus evolution experiments were performed to select for fast-replicating variants. Reversion to the wild-type PBS-lys3 sequence was the most frequent escape route. However, we identified one culture in which the virus gained replication capacity without reversion of the PBS. This revertant virus eventually optimized the PAS motif for interaction with the nonself primer. Interestingly, earlier evolution samples revealed a single amino acid change of an otherwise well-conserved residue in the RNase H domain of the RT enzyme, implicating this domain in selective primer usage. We demonstrate that both the PAS and RT mutations improve the replication capacity of the tRNA1,2Lys-using virus.
Reverse transcription is the replication step that converts a retroviral RNA genome into a DNA copy, a mechanism that is shared by retroviral elements and hepadnaviruses. Although a variety of primer molecules can be used to initiate reverse transcription, all retroviruses use a cellular tRNA primer (39,
46, 51, 52, 60). The most commonly used primers are tRNAPro
(e.g., murine leukemia virus and human T-cell leukemia virus),
tRNATrp(e.g., avian myeloblastosis virus), tRNA
1,2
Lys(e.g.,
Ma-son-Pfizer monkey virus), and tRNA3Lys(e.g., mouse mammary
tumor virus and all lentiviruses, including human immunode-ficiency virus type 1 [HIV-1]). Retroviruses are dedicated to the self tRNA primer, despite an excess of other tRNA mol-ecules in the infected cell (15, 40, 63). Selective tRNA usage seems less stringent for some retroviruses (47, 48, 64).
Primer tRNA selection in HIV-1 is accomplished by multi-ple mechanisms. First, the primer tRNA is selectively
pack-aged into virus particles (29, 30). All tRNALysisoacceptors are
packaged during particle assembly via their interaction with the Gag-Pol precursor and a protein complex composed of the cellular lysyl-tRNA synthetase and the viral Gag protein (11,
27, 28, 35, 49). Only tRNA3Lysis tightly associated with the viral
RNA (vRNA) genome (29). Second, the vRNA genome con-tains an 18-nucleotide sequence that is perfectly
complemen-tary to the 3⬘ end of the self tRNA3Lys and is termed the
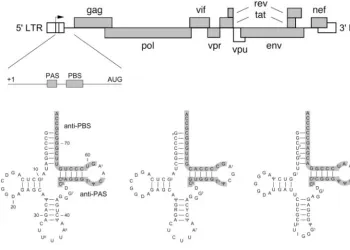
primer-binding site (PBS) (Fig. 1). It should be noted that the
PBS is not required for the selective packaging of tRNA3Lysinto
virus particles, but it is required for tight annealing of the
tRNA3Lysprimer (29). Thus, the PBS fulfills an indispensable
role in the placement of the tRNA primer onto the viral ge-nome and hence in reverse transcription (23, 45). No sponta-neous mutations or more gross tRNA switches have been re-ported. A single point mutation that is recurrently observed at a low incidence in the HIV-1 PBS results from the infrequent
usage of a low-abundance tRNA5Lys variant (16). In fact, the
viral PBS motif is inherited from the sequence of the tRNA primer during reverse transcription, and PBS mutations can revert to the wild-type (wt) sequence within one round of
replication through extension of a misaligned tRNA3Lysprimer
(14). This special reversion ability explains the almost com-plete conservation of the PBS sequence. HIV-1 mutants with an altered PBS identity exhibit a significant replication defect and rapidly revert to the wt PBS sequence (15, 40, 63). These results indicate that there are additional viral features that determine tRNA primer specificity. One obvious candidate is the reverse transcriptase (RT) polymerase, and a functional analysis of mutant HIV-1 virion particles revealed that RT is indeed involved in the selection and PBS annealing of the
tRNA3Lysprimer (45, 57). Biochemical studies have provided
additional information on the RT-tRNA3Lys complex and its
involvement with the RNase H domain (17, 54, 56, 58), but no high-resolution picture has emerged from these studies.
The PBS is not the only RNA sequence in the retroviral genome that is important for tRNA priming. Additional con-tacts between the vRNA and the self tRNA have been
re-* Corresponding author. Mailing address: Department of Human Retrovirology, Academic Medical Center, University of Amsterdam, Meibergdreef 15, 1105 AZ Amsterdam, The Netherlands. Phone: 31 20 566 4822. Fax: 31 20 566 6531. E-mail: [email protected].
† Present address: Department of Cell Biology and Genetics, Eras-mus University, Rotterdam, The Netherlands.
10706
on November 8, 2019 by guest
http://jvi.asm.org/
ported for Rous sarcoma virus (1, 2, 13, 55) and subsequently for HIV-1 and HIV-2 (3, 4, 6, 20, 22, 24, 25, 44). An interaction
was proposed between the U-rich anticodon loop of tRNA3Lys
and an A-rich loop of a hairpin structure in HIV-1 RNA that is positioned upstream of the PBS (24). Based on this base-pairing interaction, it was tested whether the usage of a non-self tRNA primer can be imposed by simultaneous adaptation of the PBS and the A-rich loop. Such double mutants exhibited a replication defect, but variants that stably used the nonself
primers tRNAHis, tRNAGlu, and tRNA
1,2
Lyswere selected upon
prolonged culturing (18, 32, 34, 41–43, 62, 65). This type of virus reversion analysis is complicated by the fact that the retroviral genome is densely packed with replication signals. For instance, the A-rich interaction site is also an important sequence element for binding of the integrase protein during insertion of the proviral HIV-1 DNA into the host chromo-some (19). Furthermore, important RNA structure motifs are encoded by this domain (3, 6, 7), and the destruction and subsequent repair of these structures may explain some of the virus reversion events (9). Despite an extensive search for second-site protein mutations that compensate for the switch of tRNA primer, no such coadaptive changes have been re-ported thus far (32, 65).
We recently presented evidence for an additional vRNA-tRNA contact between HIV-1 sequences that are positioned
upstream of the PBS and the T⌿C arm of tRNA3Lys(3, 6). This
motif does not contribute to tRNA-PBS annealing, but it is essential for the initiation of tRNA-primed reverse
transcrip-tion and was therefore termed the primer activatranscrip-tion signal (PAS) (Fig. 1). In vitro, HIV-1 primer usage can be switched
from tRNA3Lysto tRNA1,2Lysby the use of RNA templates
con-taining PAS and PBS double mutations (4). In this study, we set out to test whether PAS-PBS double mutant viruses can stably replicate with a non-self tRNA primer.
MATERIALS AND METHODS
DNA constructs.HIV-1 LAI viruses with mutations in the PBS were described previously (15). The PAS-PBS double mutants were made accordingly. All mu-tant constructs were verified by sequence analysis. The PAS-PBS–pro and –lys1,2 mutations are indicated in Fig. 3 and 4. The revertant R1 and R2 mutations (T126C and G3600A, respectively;⫹1 is the first nucleotide of the genomic HIV-1 RNA) were cloned via PCR mutagenesis into proviral DNA constructs. To clone the R1 mutation, we used the template pLAI-PBS1,2Lys(15).
Oligonu-cleotides T7-1 and TA025 (5⬘-140CTCTAGTTACGTGGGTCACACAACAGA
CGGG109-3⬘) were used to amplify nucleotides 1 to 140 of the HIV-1 leader
sequence (5). Oligonucleotides TA024 (5⬘-116GTTGTGTGACCCACGTAACT
AGAGATCCCT145-3⬘) and AD-GAG were used to amplify nucleotides 116 to
462 of the LAI-PBS1,2Lysgenome (3). The mutated nucleotides are underlined in
the primer sequences and the nucleotide positions are indicated with superscript numbers. The two PCR products were joined and amplified in a subsequent PCR with the T7-1 and AD-GAG primers. The R1 PCR fragment was sequenced, subsequently digested with HindIII and ClaI, and cloned into the Blue-5⬘LTR vector containing the XbaI-ClaI fragment of the pLAI proviral clone (36). The R1 mutated XbaI-ClaI fragment was finally cloned into pLAI-R37, a derivative of the full-length infectious clone pLAI (10). The mutant proviral construct was designated pLAI-R37-M-R1, in which M indicates the original PAS1,2Lys and
PBS1,2Lys mutations (T128A, G129C, G190A, A192C, C193G, A194T, and
A198G).
[image:2.603.117.468.72.322.2]The R2 mutation (G3600A) was cloned similarly, although Blue-Pol-ApaSal was used as a template in the initial PCRs. This pBluescript-derived plasmid
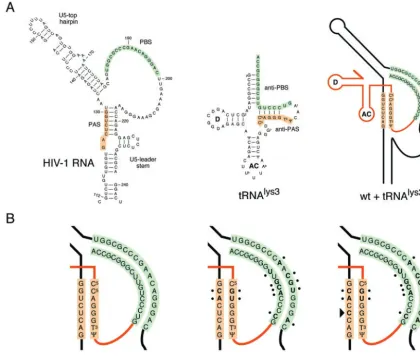
FIG. 1. HIV-1 genome and the PAS and PBS motifs that specify tRNA primer usage. The HIV-1 DNA genome is shown at the top. The 5⬘ long terminal repeat (LTR) is divided into three segments (U3, R, and U5). Transcription starts at the U3-R border (arrow). A close-up of the untranslated leader of the vRNA is shown (from the transcription start site⫹1 to the Gag start codon AUG). The PAS and PBS are indicated. The cloverleaf structures of the self tRNA3Lysprimer and the nonself primers tRNA1,2Lysand tRNAProare shown below. Base modifications in the tRNA molecules are indicated according to standard nomenclature (59). R29 in tRNA1,2Lysindicates that this position is G in tRNA1Lysand A in tRNA2Lys, and Y41 is C in tRNA1Lysand U in tRNA2LysThe anti-PAS and anti-PBS motifs are boxed. We mutated the HIV-1 PAS and PBS motifs to complement the nonself primers. Details of these mutations are shown in Fig. 3 and 4.
on November 8, 2019 by guest
http://jvi.asm.org/
contains the ApaI-SalI fragment (nucleotides 1550 to 5265) of the pLAI proviral clone. This plasmid includes the 3⬘part of the Gag coding sequence, all of the Pol and Vif coding sequences, and the 5⬘part of the Vpr coding sequence. Oligo-nucleotides TA022 (5⬘-3400GAAATTATGGTACCAGTTAGA3420-3⬘) and
TA021 (5⬘-3615ATATTTACTTCTAATTCCGAATCCT3591-3⬘) were used to
amplify nucleotides 3400 to 3615 of the HIV-1 genome. In a separate reaction, oligonucleotides TA023 (5⬘-5372AAGGTCGACACCCAATTCTGAAATGG 5350-3⬘) and TA020 (5⬘-3591AGGATTCGGAATTAGAAGTAAATAT3615-3⬘)
were used to amplify nucleotides 3591 to 5372. In a final PCR, these PCR products were joined and amplified with primers TA022 and TA023. Upon sequence analysis, the PCR fragment was digested with NheI and NcoI and then cloned into Blue-Pol-ApaSal. The R2-containing BclI (2011)-SalI fragment was cloned into R37 to obtain wt-R2, into R37-M to obtain pLAI-M-R2, and into pLAI-M-R1 to obtain the double revertant pLAI-M-R12 pro-viral construct.
Cells, virus replication, and evolution.The SupT1 T-cell line was either trans-fected by electroporation with proviral DNA or intrans-fected with a virus stock as previously described in detail (3, 6). CA-p24 levels in the culture medium were determined with an enzyme-linked immunosorbent assay. The protocol used for virus evolution by a prolonged cell-free passage of virus onto fresh, uninfected SupT1 cells was described previously (37). Isolation of total cellular DNAs was performed by proteinase K treatment (3). The long terminal repeat leader region was PCR amplified with primers T7-1 and AD-GAG. The complete RT region was PCR amplified in two overlapping segments. The 5⬘segment was amplified with the primers NB1 (5⬘-1966AAAATGATAGGGGGAATTGG1985-3⬘) and
3⬘RT-20 (5⬘-3043CTGCCAGTTCTAGCTCTGCTTC3022-3⬘), and the 3⬘segment
was amplified with 5⬘BRT (5⬘-2583GGGATGGAAAGGATCACC2600-3⬘) and
3⬘RT-22 (5⬘-3894AGGTTAAAATCACTAGCCATTGCTCTCC3866-3⬘). The
PCR products were directly sequenced, thus providing the average sequence of the viral quasispecies (population sequence).
Competition and virus stability assays.SupT1 cells were transfected as de-scribed above with a mixture of two proviral constructs (500 ng each) or were infected with the R1 or R12 virus (5 ng of CA-p24 per 5-ml culture). Competition assays were performed for at least three passages. The genetic stability of M-R1 and M-R12 was tested in cultures that were maintained for 101 to 125 days. Cell-free supernatant samples were taken at peak infection and passaged on fresh SupT1 cells every 7 to 10 days. Total cellular DNA was isolated and used as a template in a standard PCR to amplify the leader and partial RT regions. Oligonucleotides TA053 (5⬘-TGTAAAACGACGGCCAGTG1GGTCTCTCTG
GTTAGACCAG22-3⬘) and AD-GAG were used to amplify the leader region.
Primers TA052 (5⬘-TGTAAAACGACGGCCAGT3472CAGGGAGACTAAAT
TAGG3489-3⬘) and NB4 (5⬘-3865ATTACTGTGATATTTCTCATG3845-3⬘) were
used to amplify nucleotides 3472 to 3845 of the RNA genome (RT region). Both PCR products were extended with M13-derived sequences present in the TA052 and TA053 primers. The products were sequenced with BigDye-labeled⫺21 M13 primers (5⬘-TGTAAAACGACGGCCAGT-3⬘) by use of an ABI Prism BigDye primer cycle sequencing ready reaction kit (Applied Biosystems).
RESULTS
Replication of PAS-PBS mutant viruses. We constructed HIV-1 variants with PAS-PBS double mutations to enforce the
use of the non-self primers tRNAProand tRNA
1,2
Lys(Fig. 1). The
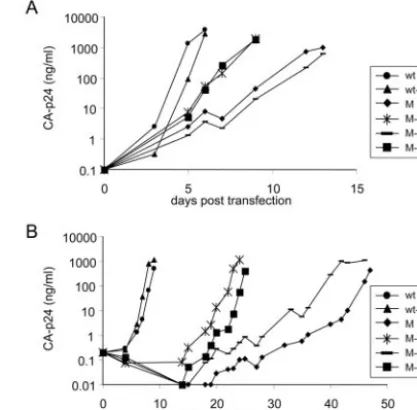
mutations were introduced in the LAI molecular clone, a CXCR4-using primary HIV-1 isolate, and the two mutants were termed PAS/PBS-pro and PAS/PBS-lys1,2. These plas-mids were transfected into the SupT1 T-cell line to monitor virus replication by the increase in CA-p24 production in the culture supernatant (Fig. 2). We included the wt LAI control and single PBS mutants (termed PBS-pro and PBS-lys1,2) (15).
The 10-g transfection reactions illustrated the ranking order
of replication: wt⬎PBS-lys1,2⬎PBS-pro⬎PAS/PBS-pro⬎
PAS/PBS-lys1,2. The 5-g transfection reactions confirmed the
severe replication defect of all mutants except PBS-lys1,2. The relatively efficient replication of this PBS-lys1,2 single mutant was reported previously (15). The addition of the PAS-lys1,2 mutation decreased virus replication. The single PBS-pro mu-tant showed a significant replication defect, and inclusion of the PAS-pro mutation further weakened virus replication. The
FIG. 2. Replication of wt HIV-1 LAI and PAS and PBS mutated viruses. SupT1 cells were transfected with 10g (A) or 5g (B) of the proviral constructs. CA-p24 production was measured in the culture medium at several days posttransfection.
FIG. 3. Evolution of PAS/PBS-pro mutant. SupT1 cells were trans-fected with 5g of the molecular clones. Breakthrough replication was observed in some cultures, and viruses could eventually be passaged onto fresh cells. The identity of the PBS motif is indicated as a function of the evolution time (A). The input mutant PBS is shown with open boxes, and wt revertants are shown with black boxes. (B) Culture number, day of harvest, and sequence of proviral DNA isolated from infected cells. The mutated PAS and PBS nucleotides are depicted in bold and are underlined. Nucleotide changes acquired during evolu-tion are shown in white surrounded by a black box (“N” indicates a mixed sequence).
10708 ABBINK ET AL. J. VIROL.
on November 8, 2019 by guest
http://jvi.asm.org/
negative impact of the PAS mutation seemed less dramatic for the PBS-pro virus than for the PBS-lys1,2 virus. In any case, the PAS-PBS double mutants did not rescue the replication of the PBS single mutants.
Evolution of PAS-PBS mutant viruses.We next set out to obtain faster replicating revertant viruses for the double mu-tants. A priori, two evolution routes could be envisaged. First,
the virus could restore tRNA3Lys usage by reversion to the wt
PBS motif. PBS reversion is a frequent event that is driven by
mispriming of tRNA3Lyson the mutant PBS, followed by
inher-itance of the fully wt PBS sequence from the tRNA primer.
However, priming by tRNA3Lyswas inhibited at two levels by
the PAS-PBS mutations, at the levels of tRNA annealing and initiation of reverse transcription. In contrast, the usage of the new primer was enhanced by these mutations. These combined effects may block the wt reversion route. Second, the virus could optimize replication with the new tRNA primer, and adaptive changes could thus be acquired in the viral RT en-zyme or other cofactors. This latter evolutionary route is very interesting because it may reveal critical amino acid residues within RT that play an important role in selective tRNA usage. We maintained 21 and 17 independent cultures with the PAS/PBS-pro and PAS/PBS-lys1,2 mutants, respectively, and breakthrough replication was monitored within a few weeks in 4 and 3 cultures. Viruses were passaged repeatedly. Cell sam-ples were taken and used to amplify proviral DNAs for subse-quent sequence analysis. We initially focused on the leader RNA that encodes the PAS and PBS motifs. The evolutionary routes are summarized in Fig. 3A and 4A. Partial leader
se-quences of the revertant PAS/PBS-pro and PAS/PBS-lys1,2 viruses are shown in Fig. 3B and 4B. Sequence analysis of the PAS/PBS-pro viral DNA revealed reversion to the wt PBS-lys3 within 47 days in three of the four positive cultures (Fig. 3, cultures P8, P11, and P18). The P18 virus contained an addi-tional mutation immediately upstream of the PBS (G181A). Such changes in the PBS-flanking residues have been de-scribed for other viral reversion events (8, 15, 31, 34). The fourth positive culture (P14) appeared more interesting be-cause the PAS/PBS-pro motifs were stably maintained. This culture was split at day 47 into six subcultures that were mon-itored over time (Fig. 3A). All samples eventually reverted back to the wt PBS-lys3 sequence at day 116 (Fig. 3B). In addition, we observed reversion to the wt PAS-lys3 by means of the G128U back mutation in two cultures (P14.3 and P14.6), and the population-based sequence showed a mixed sequence for one culture (P14.5). This finding confirms the importance of having a fully complementary PAS motif. In conclusion, we were unable to select a replicating virus that stably used the
tRNAProprimer.
Identification of a stable virus using tRNA1,2Lys. We
specu-lated that the evolutionary jump in primer usage from tRNA3Lys
to tRNA1,2Lyswould be easier for HIV-1 because these primers
are very similar (Fig. 1). Three cultures showed syncytia over time, indicating replication of PAS/PBS-lys1,2 revertants (Fig. 4A, cultures L3, L4, and L8). Variant L8 had reverted to wt PBS-lys3 at day 47 (Fig. 4B). The virus in culture L3 showed a mixed wt-mutant PBS sequence at day 73 and had reverted back to PBS-lys3 at day 97. The input PAS/PBS-lys1,2 motifs were maintained only in culture L4. The L4 virus continued to
replicate with a tRNA1,2Lys primer up to day 75, but a mixed
[image:4.603.317.526.74.279.2]FIG. 4. Evolution of PAS/PBS-lys1,2 mutant. See the legend for Fig. 3 for details. The two mutated PAS and PBS nucleotides are indicated in bold and are underlined. Mutations in the region just upstream of the PAS that were observed in some cultures may reflect G-to-A hypermutations. These transitions have been described previ-ously for other leader revertant viruses and were therefore not ana-lyzed further (8).
FIG. 5. Replication of wt, PAS/PBS-lys1,2 mutant, and cloned re-vertant viruses. SupT1 cells were transfected with 2g of the proviral constructs (A) or were infected with equal amounts of viruses (1 ng of CA-p24 per 5 ml of culture) (B). CA-p24 production was measured in the culture medium for several days. M indicates the original PAS/ PBS-lys1,2 mutant, R1 indicates the U126C reversion in the PAS motif, and R2 indicates the G490E reversion in the RNase H domain of RT.
on November 8, 2019 by guest
http://jvi.asm.org/
wt-mutant PBS sequence was detected at day 97. We used the day 47 sample to restart a second round of evolution by infect-ing six fresh SupT1 cell cultures in parallel. All six cultures became infected with L4-derived variants that stably main-tained the mutant PBS-lys1,2 up to day 116. This observation suggests that the L4 viruses acquired at least one adaptive
change outside the PBS motif to accommodate tRNA1,2Lysin the
second round of evolution. An interesting change was observed within the PAS motif in five of six revertants (Fig. 4B). The mutant PAS-lys1,2 motif differed from the wt PAS-lys3 ele-ment at two nucleotide positions. These nucleotides did not revert, but an additional PAS residue was altered (U126C). This is interesting, because a U-G base pair is replaced with a stronger C-G base pair in the context of the PAS–anti-PAS
interaction with tRNA1,2Lys.
The original L4 revertant did not yet contain the U126C PAS adaptation at day 47. Nevertheless, this virus replicated relatively efficiently, suggesting that at least one other critical mutation must be present elsewhere in the viral genome to
facilitate reverse transcription primed by the non-self tRNA1,2Lys
molecule. Replication studies confirmed the efficient replica-tion of the L4-d47 and L4-d76 viruses (results not shown). Thus, we assumed that the L4-d47 virus has a major adaptive change elsewhere in the viral genome that allows efficient
tRNA1,2Lysusage. Because no significant changes were present in
the leader domain surrounding the PAS and PBS motifs (re-sults not shown), we sequenced the complete RT gene. One mutation (G3600A) within the RT gene of the L4-d47 virus was identified. This nucleotide transition led to an amino acid change (G490E) in the RNase H domain of RT. The mutation was stably maintained in later samples (L4.1, L4.2, and L4.3 were sequenced). The G490 residue is absolutely conserved among virus isolates of all HIV-1 subtypes (data not shown), and the mutation was therefore further analyzed.
Role of U126C and G3600A mutations in stable tRNA1,2Lys
primer usage.To confirm that the PAS mutation U126C (des-ignated R1) and the RT mutation G3600A (des(des-ignated R2) increase the replication of the PAS/PBS-lys1,2 mutant virus
and stabilize the usage of the tRNA1,2Lysprimer, we cloned these
mutations into the original PAS/PBS-lys1,2 mutant (further designated with an “M,” yielding M-R1, M-R2, and M-R12 [a mutant with both R1 and R2 changes]). In addition, the RT mutation was tested in a wt virus background (wt-R2). Molec-ular clones were constructed and transfected into SupT1 cells to monitor virus replication (Fig. 5A). The R1 PAS change
significantly increased the replication of the tRNA1,2Lys-using
virus, even in the absence of the R2 reversion in RT. No gross effect of R2 on the replication of the wt and mutant viruses could be observed. More sensitive virus competition assays
were performed to measure small differences in virus replica-tion (Table 1). From these data, we determined the following
ranking order: wtⱖwt-R2⬎⬎M-R1ⱖM-R12⬎M-R2⬎M.
Apparently, the R2 reversion significantly improved replica-tion of the M virus, because M-R2 efficiently outcompeted M after a single passage. Interestingly, the same R2 mutation slightly decreased the replication capacity of the wt and M-R1 viruses.
To confirm these results, we monitored virus replication in SupT1 cells that were inoculated with a limiting amount of virus (Fig. 5B). The infection data confirmed the ranking order observed in the competition experiments and indicated that both the individual R1 mutation and the individual R2 muta-tion improve the replicamuta-tion of the PAS/PBS-lys1,2 mutant. The impact of the R1 mutation on the restoration of virus replication was significantly larger than that of the R2
muta-tion. R1 may also stabilize tRNA1,2Lysusage more potently than
R2. Virus L4-d47, containing only R2 and not R1, still reverted
to the tRNA3Lys-using variant L4-d97 (Fig. 4). However, L4-d47
remained stable in its tRNA1,2Lysusage upon acquiring R1
(L4-d116 variants 1, 2, 3, 5, and 6) in the second evolution round.
To confirm that R1 and R2 can stabilize tRNA1,2Lysusage, we
passaged the molecularly cloned M-R1 and M-R12 viruses in six cultures per virus for a period of 101 to 125 days. The sequences of the leader RNAs and the RT genes were
ana-lyzed. Most importantly, all viruses continued to use tRNA1,2Lys.
In a single M-R12 culture, the R2 mutation was mixed with the wt sequence after 125 days, confirming that M-R1 is a rela-tively fitter virus than M-R12. The R1 reversion was main-tained in the entire quasispecies population in all cultures. These data suggest that R1 effectively prevents the switch to
tRNA3Lysusage. A schematic overview of the evolution events
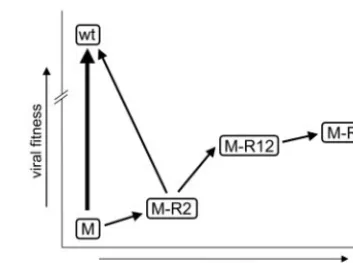
and their effect on viral fitness is shown in Fig. 6. Back
[image:5.603.329.506.69.200.2]muta-FIG. 6. Schematic representation of the gain in relative viral fitness during forced evolution of the PAS/PBS-lys1,2 mutant (M). Arrows indicate reversion events. The thickness of the arrows indicates the chance of reversion. The slope of the arrow indicates the gain of fitness. M indicates the original PAS/PBS-lys1,2 mutant, which pre-dominantly reverts to a wt, tRNA3Lys-using virus. In the L4 culture (Fig. 4), M acquired an R2 reversion in the RT gene (G3600A). The M-R2 virus is fitter, but it can still revert back to the wt. Viral fitness increases most significantly upon acquisition of the R1 mutation in the PAS element. The M-R12 double revertant is stable in prolonged cultures and does not revert back to the wt. From competition experiments, we concluded that M-R1 replicates slightly more efficiently than M-R12. Consistent with this, the M-R12 virus lost the R2 mutation in a single culture. M-R1 stably maintained the PBS-lys1,2 sequence in all cul-tures, highlighting the importance of the PAS motif in tRNA1,2Lysusage. TABLE 1. Comparisons of viruses for fitness
Competing virusesa Fittest virus Passage no.b
wt and wt-R2 wt 4
M and M-R1 M-R1 1
M-R1 and M-R12 M-R1 2
M and M-R12 M-R12 1
M and M-R2 M-R2 1
aMixed 1:1 at start of experiment.
bPassage at which the fittest virus was⬎95% of the total virus.
10710 ABBINK ET AL. J. VIROL.
on November 8, 2019 by guest
http://jvi.asm.org/
[image:5.603.42.285.81.150.2]tion to tRNA3Lysusage occurs frequently for the M mutant and
is still possible for the M-R2 revertant but is not observed for the M-R12 revertant and the more fit M-R1 revertant.
DISCUSSION
To obtain an HIV-1 virus that stably uses a primer other
than tRNA3Lys for reverse transcription, we altered the PAS
and PBS sequences in the HIV-1 leader RNA. Earlier exper-iments from our laboratory and other laboratories had shown that altering the PBS sequence alone was not sufficient to stably switch tRNA usage (15, 40, 63). Additional studies showed that an upstream RNA PAS motif is critically involved in tRNA-primed reverse transcription (3, 6). The PAS motif exerts its function not by enhancing tRNA annealing to the PBS, but by activating the initiation of reverse transcription. The PAS motif engages in a base-pairing interaction with the
complementary anti-PAS sequence in the tRNA, which likely results in the formation of a higher order RNA structure that is suitable for reverse transcription (Fig. 7). By adaptation of both the PAS and PBS motifs, the HIV-1 leader could be
changed to accommodate the tRNA1,2Lys primer for the
initia-tion of reverse transcripinitia-tion in vitro (4). We reasoned that the single PBS mutants could not optimally initiate non-self tRNA-primed reverse transcription in vivo because of the dis-rupted PAS–anti-PAS base-pairing interaction. We therefore constructed PAS-PBS double mutants that could properly
ac-commodate either tRNAProor tRNA
1,2
Lysas a reverse
transcrip-tion primer. The replicatranscrip-tion efficiencies of these mutant viruses were compared to those of single PBS mutants. The single PBS-lys1,2 mutant replicated relatively efficiently, but the sin-gle PBS-pro mutant showed a significant replication defect. The addition of a PAS mutation further decreased virus
rep-FIG. 7. Model for reverse transcription initiation on wt, PAS/PBS-lys1,2 mutant, and R1 revertant templates. (A) The secondary structures of the PBS region of the HIV-1 RNA genome and tRNA primers are shown schematically (black and orange lines, respectively) AC, anticodon loop; D, D loop. The tRNA primer anneals with its 3⬘-terminal 18 nucleotides to the PBS (the PBS and anti-PBS sequences are shown in green). An additional interaction between PAS and anti-PAS (orange) is required to activate the initiation of reverse transcription. These interactions are indicated for the wt leader with tRNA3Lysand for the PAS/PBS-lys1,2 mutant (M) and the M-R1 revertant with tRNA1,2Lys(B). The sequence differences between PAS and PBS sequences of wt, M, and R1 leader RNAs and the anti-PAS and anti-PBS sequences of tRNA3Lysand tRNA1,2Lys are indicated in bold and marked with dots. The R1 reversion (U126C; indicated by an arrowhead) stabilizes the PAS–anti-PAS interaction (replacement of a weak U-G base pair with a very stable C-G base pair).
on November 8, 2019 by guest
http://jvi.asm.org/
[image:6.603.82.503.75.438.2]lication, indicating that PAS adaptation does not rescue repli-cation of the PBS single mutants. Rescue would be expected if no other viral factors are implicated in selective tRNA usage. However, several viral factors that closely interact with the tRNA primer, such as the RT enzyme, may be involved in primer selection. Thus, other incompatibility problems may not be solved by the imposed usage of a non-self tRNA primer
in the PAS-PBS double mutants. The latter scenario is consis-tent with the experimental results.
The tRNAPro-using virus was not stable during prolonged
passaging, despite multiple attempts. Its PBS quickly reverted
to the wt tRNA3Lys complementary sequence. This negative
result may be explained by a lack of specific packaging of
tRNAProinto virus particles. tRNALysisoacceptors are
specif-ically packaged in wt virus particles via an interaction with Gag-Pol and the cellular lysyl-tRNA synthetase (11, 27, 35, 49, 50). Both Gag and Pol sequences are required for the forma-tion of this tRNA-packaging complex. Therefore, it may be
difficult for HIV-1 to acquire the ability to package tRNAPro.
Adaptations in both Gag and Pol may be required to allow
prolyl-tRNA synthetase and tRNAPropackaging and
appropri-ate primer annealing onto PBS-pro. At present, it is unclear how the specificity of tRNA synthetase packaging can be al-tered.
We were more successful in switching the reverse
transcrip-tion primer from tRNA3Lysto tRNA1,2Lys. Since tRNA1,2Lys
isoac-ceptors are selectively packaged via the Gag-Pol–lysyl-tRNA complex, the PAS/PBS-lys1,2 virus was not expected to
en-counter difficulties in tRNA1,2Lys packaging. Nevertheless, the
PAS/PBS-lys1,2 virus was severely affected in its replication
efficiency and could still revert to a tRNA3Lys-using virus, but
the acquisition of a second-site reversion in the PAS motif
(U126C [R1]) stabilized the tRNA1,2Lys usage. In addition, a
second-site reversion was located in the RNase H domain of the RT protein (G490E [R2]). The R2 reversion improved the
stability of tRNA1,2Lysusage, although not completely. Both R1
and R2 reversions were introduced in the original lys1,2 virus and improved the replication of the PAS/PBS-lys1,2 mutant virus.
Others have described similar non-tRNA3Lys-using viruses,
but no adaptive changes in the PAS motif or any of the viral proteins have been identified (18, 31–34, 43, 62, 65, 66). These authors did report changes in the A-rich loop located upstream of the PBS. The A-rich loop has been suggested to engage in a base-pairing interaction with the anticodon loop of the
prim-ing tRNA. The anticodon loop sequences of tRNA3Lys and
tRNA1,2Lysdiffer by only one nucleotide (position 34; Fig. 1). For
a restoration of this putative base pairing, one would expect an A170G change to occur. In our evolution studies, which lasted
⬎3 months, we did not observe this or an equivalent
adapta-tion of the A-rich loop to accommodate the tRNA1,2Lysmolecule.
These results indicate that full base-pairing potential between these RNA sequences is not absolutely required for tRNA selection. In general, mutant-revertant data should be inter-preted with caution because this part of the viral genome encodes multiple, overlapping functions. The RNA secondary structure surrounding the A-rich loop is important for viral replication and reverse transcription (5, 7), and these se-quences are required for proper integration of the provirus into the host genome (61).
The R1 reversion in the PAS motif stabilized base pairing
with the anti-PAS motif in the tRNA1,2Lysprimer (Fig. 7). This
reversion increased viral replication and stabilized tRNA1,2Lys
usage, indicating that the designed PAS–anti-PAS interaction of the original mutant was not sufficiently stable and was there-fore suboptimal. The PAS motif was recently questioned by
FIG. 8. Multiple interactions between HIV-1 RT enzyme, RNA genome, and tRNA3Lys primer. (A) X-ray structure of the p51-p66 heterodimer of the HIV-1 RT enzyme with a double-stranded DNA duplex (26). The enzymatically active p66 domain is shown in blue, and the RNase H domain is shown in yellow. The catalytically active resi-dues in both domains are shown as red dots. The p51 subunit is shown in green. The figure was drawn with Molscript (38) and Raster3D (53) software, using coordinates from the protein data bank (entry 1 HMI [26]). (B) Cartoon of the complex between the HIV-1 RT enzyme, the viral RNA genome, and the self tRNA3Lysprimer. The PBS–anti-PBS base-pairing interaction is indicated. We also marked other interaction domains, including the U-rich anticodon (AC) loop of tRNA3Lys, an A-rich segment in the viral RNA genome (green), and the anti-PAS and PAS motifs (yellow). We selected an HIV-1 variant that switched its primer usage from tRNA3Lysto tRNA1,2Lysby simultaneous mutation of the PBS and PAS motifs. This virus acquired a point mutation in the RNase H domain (Gly490Glu; shown in yellow), which implicates this RT domain in primer binding. The sequence of the anticodon stem is the most variable region between tRNA3Lysand tRNA1,2Lys(marked in red; see Fig. 1 for details), and this tRNA domain may interact with the gatekeeper residue 490. The anticodon hairpin is proposed to dock in the cleft between the RNase H and RT domains of the p66 subunit.
10712 ABBINK ET AL. J. VIROL.
on November 8, 2019 by guest
http://jvi.asm.org/
others (21), despite a wealth of experimental evidence,
includ-ing the successful switch to tRNA1,2Lysusage in vitro by a
simul-taneous change of the PAS and PBS sequences (3, 4, 6, 23). Also, an equivalent interaction has been described for HIV-2 and avian sarcoma virus (1, 2, 12, 20, 55). Our results obtained with the evolved PAS/PBS-lys1,2 virus clearly confirm the im-portance of the PAS motif in tRNA usage in vivo. Fine-tuning of the PAS–anti-PAS interaction strength turns out to be the most decisive change for yielding a virus that stably uses
tRNA1,2Lys. The R1 mutation also improves base-pairing
poten-tial with the natural tRNA3Lysprimer. A previous
demonstra-tion that this mutademonstra-tion enhances tRNA3Lys usage in vitro is
consistent with this notion (4).
The R2 reversion in the RT protein (G490E) specifically
improved the replication and stability of the tRNA1,2Lys-using
virus. This amino acid change did not significantly affect viral replication in a wt and R1 background. Apparently, R2 only stimulates viral replication when the tRNA-vRNA interaction is suboptimal. Glycine 490 is conserved in the RT gene of the
related SIV and HIV-2 viruses that also use the tRNA3Lys
primer. Interestingly, residue 490 (shown in yellow in Fig. 8) protrudes from the RNase H domain in the X-ray structure of the RT p51-p66 heterodimer. Thus, residue 490 seems ideally positioned to act as a “gatekeeper” for the cleft between the RNase H and RT domains. We speculate that part of the tRNA molecule binds in this cleft, which is consistent with previous cross-linking studies (54). Because residue 490 is a
candidate for the discrimination between the self tRNA3Lysand
non-self tRNA1,2Lys primers, we juxtaposed the most variable
tRNA domain in Fig. 8 (the stem of the anticodon [AC] hair-pin, shown in red). These results imply that the RNase H domain of HIV-1 RT is involved in selective tRNA binding. Further in vitro studies with recombinant RT may shed more light on the specific role of the RNase H domain in tRNA selection.
ACKNOWLEDGMENTS
We thank Rogier Sanders for generating the RasMol picture and Wim van Est for creating an artist’s impression.
This work was supported by NWO-CW (The Netherlands Organi-zation for Scientific Research).
REFERENCES
1. Aiyar, A., D. Cobrinik, Z. Ge, H. J. Kung, and J. Leis.1992. Interaction between retroviral U5 RNA and the TYC loop of the tRNA(Trp) primer is required for efficient initiation of reverse transcription. J. Virol.66:2464– 2472.
2. Aiyar, A., Z. Ge, and J. Leis.1994. A specific orientation of RNA secondary structures is required for initiation of reverse transcription. J. Virol.68:611– 618.
3. Beerens, N., and B. Berkhout.2002. The tRNA primer activation signal in the human immunodeficiency virus type 1 genome is important for initiation and processive elongation of reverse transcription. J. Virol.76:2329–2339. 4. Beerens, N., and B. Berkhout.2002. Switching the in vitro tRNA usage of
HIV-1 by simultaneous adaptation of the PBS and PAS. RNA8:357–369. 5. Beerens, N., F. Groot, and B. Berkhout.2000. Stabilization of the U5-leader
stem in the HIV-1 RNA genome affects initiation and elongation of reverse transcription. Nucleic Acids Res.28:4130–4137.
6. Beerens, N., F. Groot, and B. Berkhout.2001. Initiation of HIV-1 reverse transcription is regulated by a primer activation signal. J. Biol. Chem.276: 31247–31256.
7. Beerens, N., B. Klaver, and B. Berkhout.2000. A structured RNA motif is involved in the correct placement of the tRNALys3primer onto the human
immunodeficiency virus genome. J. Virol.74:2227–2238.
8. Berkhout, B., A. T. Das, and N. Beerens.2001. HIV-1 RNA editing, hyper-mutation and error-prone reverse transcription. Science292:7.
9. Berkhout, B.1997. The primer-binding site on the RNA genome of human
and simian immunodeficiency viruses is flanked by an upstream hairpin structure. Nucleic Acids Res.25:4013–4017.
10. Berkhout, B., J. van Wamel, and B. Klaver.1995. Requirements for DNA strand transfer during reverse transcription in mutant HIV-1 virions. J. Mol. Biol.252:59–69.
11. Cen, S., H. Javanbakht, S. Kim, K. Shiba, R. Craven, A. Rein, K. Ewalt, P. Schimmel, K. Musier-Forsyth, and L. Kleiman.2002. Retrovirus-specific packaging of aminoacyl-tRNA synthetases with cognate primer tRNAs. J. Virol.76:13111–13115.
12. Cobrinik, D., A. Aiyar, Z. Ge, M. Katzman, H. Huang, and J. Leis.1991. Overlapping retrovirus U5 sequence elements are required for efficient in-tegration and initiation of reverse transcription. J. Virol.65:3864–3872. 13. Cobrinik, D., L. Soskey, and J. Leis.1988. A retroviral RNA secondary
structure required for efficient initiation of reverse transcription. J. Virol. 62:3622–3630.
14. Das, A. T., and B. Berkhout.1995. Efficient extension of a misaligned tRNA-primer during replication of the HIV-1 retrovirus. Nucleic Acids Res.23: 1319–1326.
15. Das, A. T., B. Klaver, and B. Berkhout.1995. Reduced replication of human immunodeficiency virus type 1 mutants that use reverse transcription primers other than the natural tRNA(3Lys). J. Virol.69:3090–3097.
16. Das, A. T., B. Klaver, and B. Berkhout.1997. Sequence variation of the HIV primer-binding site suggests the use of an alternative tRNALys molecule in reverse transcription. J. Gen. Virol.78:837–840.
17. Dufour, E., J. Reinbolt, M. Castroviejo, B. Ehresmann, S. Litvak, L. Tar-rago-Litvak, and M.-L. Andreola.1999. Cross-linking localization of a HIV-1 reverse transcriptase peptide involved in the binding of primer tRNALys3. J. Mol. Biol.285:1339–1346.
18. Dupuy, L. C., N. J. Kelly, T. E. Elgavish, S. C. Harvey, and C. D. Morrow. 2003. Probing the importance of tRNA anticodon: human immunodeficiency virus type 1 (HIV-1) RNA genome complementarity with an HIV-1 that selects tRNA(Glu) for replication. J. Virol.77:8756–8764.
19. Esposito, D., and R. Craigie.1998. Sequence specificity of viral end DNA binding by HIV-1 integrase reveals critical regions for protein-DNA inter-action. EMBO J.17:5832–5843.
20. Freund, F., F. Boulme, S. Litvak, and L. Tarrago-Litvak.2001. Initiation of HIV-2 reverse transcription: a secondary structure model of the RNA-tRNA(Lys3) duplex. Nucleic Acids Res.29:2757–2765.
21. Goldschmidt, V., C. Ehresmann, B. Ehresmann, and R. Marquet.2003. Does the HIV-1 primer activation signal interact with tRNA(3)(Lys) during the initiation of reverse transcription? Nucleic Acids Res.31:850–859. 22. Huang, Y., A. Shalom, Z. Li, J. Wang, J. Mak, M. A. Wainberg, and L.
Kleiman.1996. Effects of modifying the tRNALys3anticodon on the
initia-tion of human immunodeficiency virus type 1 reverse transcripinitia-tion. J. Virol. 70:4700–4706.
23. Huthoff, H., K. Bugala, J. Barciszewski, and B. Berkhout.2003. On the importance of the primer activation signal for initiation of tRNA(Lys3)-primed reverse transcription of the HIV-1 RNA genome. Nucleic Acids Res. 31:5186–5194.
24. Isel, C., C. Ehresmann, G. Keith, B. Ehresmann, and R. Marquet.1995. Initiation of reverse transcription of HIV-1: secondary structure of the HIV-1 RNA/tRNA(3Lys) (template/primer). J. Mol. Biol.247:236–250. 25. Iwatani, Y., A. E. Rosen, J. Guo, K. Musier-Forsyth, and J. G. Levin.2003.
Efficient initiation of HIV-1 reverse transcription in vitro. Requirement for RNA sequences downstream of the primer binding site abrogated by nu-cleocapsid protein-dependent primer-template interactions. J. Biol. Chem. 278:14185–14195.
26. Jacobo-Molina, A., J. Ding, R. G. Nanni, A. D. C. Clark, Jr., X. Lu, C. Tantillo, R. L. Williams, G. Kamer, A. L. Ferris, P. Clark, A. Hizi, S. H. Hughes, and E. Arnold.1993. Crystal structure of human immunodeficiency virus type 1 reverse transcriptase complexed with double-stranded DNA at 3.0 A˚ resolution shows bent DNA. Proc. Natl. Acad. Sci. USA90:6320–6324. 27. Javanbakht, H., R. Halwani, S. Cen, J. Saadatmand, K. Musier-Forsyth, H. Gottlinger, and L. Kleiman.2003. The interaction between HIV-1 gag and human lysyl-tRNA synthetase during viral assembly. J. Biol. Chem.278: 27644–27651.
28. Jiang, M., J. Mak, Y. Huang, and L. Kleiman.1994. Reverse transcriptase is an important factor for the primer tRNA selection in HIV-1. Leukemia 8:(Suppl. 1):S149–S151.
29. Jiang, M., J. Mak, A. Ladha, E. Cohen, M. Klein, B. Rovinski, and L. Kleiman.1993. Identification of tRNAs incorporated into wild-type and mutant human immunodeficiency virus type 1. J. Virol.67:3246–3253. 30. Jiang, M., J. Mak, M. A. Wainberg, M. A. Parniak, E. Cohen, and L.
Kleiman.1992. Variable tRNA content in HIV-1 IIIB. Biochem. Biophys. Res. Commun.185:1005–1015.
31. Kang, S. M., and C. D. Morrow.1999. Genetic analysis of unique human immunodeficiency virus type 1 (HIV-1) with a primer binding site comple-mentary to tRNAMetsupports a role for U5-PBS stem-loop RNA structures
in initiation of HIV-1 reverse transcription. J. Virol.73:1818–1827. 32. Kang, S.-M., J. K. Wakefield, and C. D. Morrow.1996. Mutations in both the
U5 region and the primer-binding site influence the selection of the tRNA used for the initiation of HIV-1 reverse transcription. Virology222:401–414.
on November 8, 2019 by guest
http://jvi.asm.org/
33. Kang, S.-M., Z. Zhang, and C. D. Morrow.1997. Identification of a sequence within U5 required for human immunodeficiency virus type 1 to stably maintain a primer binding site complementary to tRNAMet. J. Virol.71:207–
217.
34. Kang, S.-M., Z. Zhang, and C. D. Morrow.1999. Identification of a human immunodeficiency virus type 1 that stably uses tRNAlys1,2 rather than tRNAlys3 for initiation of reverse transcription. Virology257:95–105. 35. Khorchid, A., H. Javannbakht, S. Wise, R. Halwani, M. A. Parniak, M. A.
Wainberg, and L. Kleiman.2000. Sequences within Pr160gag-pol affecting the selective packaging of primer tRNAlys3 into HIV-1. J. Mol. Biol.299: 17–26.
36. Klaver, B., and B. Berkhout.1994. Comparison of 5⬘and 3⬘long terminal repeat promoter function in human immunodeficiency virus. J. Virol.68: 3830–3840.
37. Klaver, B., and B. Berkhout.1994. Evolution of a disrupted TAR RNA hairpin structure in the HIV-1 virus. EMBO J.13:2650–2659.
38. Kraulis, P. J.1991. MOLSCRIPT: a program to produce both detailed and schematic plots of protein structures. J. Appl. Crystallogr.24:946–950. 39. Leis, J., A. Aiyar, and D. Cobrinik.1993. Regulation of initiation of reverse
transcription of retroviruses, p. 33–48.InA. M. Skalka and S. P. Goff (ed.), Reverse transcriptase. Cold Spring Harbor Laboratory Press, Cold Spring Harbor, N.Y.
40. Li, X., J. Mak, E. J. Arts, Z. Gu, L. Kleiman, M. A. Wainberg, and M. A. Parniak.1994. Effects of alterations of primer-binding site sequences on human immunodeficiency virus type 1 replication. J. Virol.68:6198–6206. 41. Li, Y., S.-M. Kang, and C. D. Morrow.1997. Stability of HIV type 1 proviral
genomes that contain two distinct primer-binding sites. AIDS Res. Hum. Retrovir.13:253–262.
42. Li, Y., Z. Zhang, S.-M. Kang, J. L. Buescher, and C. D. Morrow.1997. Insights into the interaction between tRNA and primer binding site from characterization of a unique HIV-1 virus which stably maintains dual PBS complementarity to tRNAGly and tRNAHis. Virology238:273–282. 43. Li, Y., Z. Zhang, J. K. Wakefield, S.-M. Kang, and C. D. Morrow.1997.
Nucleotide substitutions within U5 are critical for efficient reverse transcrip-tion of human immunodeficiency virus type 1 with a primer binding site complementary to tRNAHis. J. Virol.71:6315–6322.
44. Liang, C., X. Li, L. Rong, P. Inouye, Y. Quan, L. Kleiman, and M. A. Wainberg.1997. The importance of the A-rich loop in human immunodefi-ciency virus type 1 reverse transcription and infectivity. J. Virol.71:5750– 5757.
45. Liang, C., L. Rong, N. Morin, E. Cherry, Y. Huang, L. Kleiman, and M. A. Wainberg.1997. The roles of human immunodeficiency virus type 1 Pol protein and the primer binding site in the placement of primer tRNALys3
onto viral genomic RNA. J. Virol.71:9075–9086.
46. Litvak, S., and A. Araya.1982. Primer transfer RNA in retroviruses. Trends Biochem. Sci.7:361–364.
47. Lund, A. H., M. Duch, J. Lovmand, P. Jorgensen, and F. S. Pedersen.1993. Mutated primer binding sites interacting with different tRNAs allow efficient murine leukemia virus replication. J. Virol.67:7125–7130.
48. Lund, A. H., M. Duch, and F. S. Pedersen.2000. Selection of functional tRNA primers and primer binding site sequences from a retroviral combi-natorial library: identification of new functional tRNA primers in murine leukemia virus replication. Nucleic Acids Res.28:791–799.
49. Mak, J., M. Jiang, M. A. Wainberg, M. L. Hammarskjold, D. Rekosh, and L. Kleiman.1994. Role of Pr160gag-pol in mediating the selective incorpora-tion of tRNA(Lys) into human immunodeficiency virus type 1 particles. J. Virol.68:2065–2072.
50. Mak, J., A. Khorchid, Q. Cao, Y. Huang, I. Lowy, M. A. Parniak, V. R. Prasad, M. A. Wainberg, and L. Kleiman.1997. Effects of mutations in Pr160gag-pol upon tRNALys3 and Pr160gag-pol incorporation into HIV-1. J. Mol. Biol.265:419–431.
51. Mak, J., and L. Kleiman.1997. Primer tRNAs for reverse transcription. J. Virol.71:8087–8095.
52. Marquet, R., C. Isel, C. Ehresmann, and B. Ehresmann.1995. tRNAs as primer of reverse transcriptases. Biochimie77:113–124.
53. Merritt, E. A., and D. J. Bacon.1997. Raster3D: photorealistic molecular graphics. Methods Enzymol.277:505–525.
54. Mishima, Y., and J. A. Steitz.1995. Site-specific crosslinking of 4-thiouri-dine-modified human tRNALys3 to reverse transcriptase from human im-munodeficiency virus type 1. EMBO J.14:2679–2687.
55. Morris, S., M. Johnson, E. Stavnezer, and J. Leis.2002. Replication of avian sarcoma virus in vivo requires an interaction between the viral RNA and the TC loop of the tRNATrpprimer. J. Virol.76:7571–7577.
56. Oude Essink, B. B., A. T. Das, and B. Berkhout.1995. Structural require-ments for the binding of tRNA Lys3 to reverse transcriptase of the human immunodeficiency virus type 1. J. Biol. Chem.270:23867–23874.
57. Oude Essink, B. B., A. T. Das, and B. Berkhout.1996. HIV-1 reverse transcriptase discriminates against non-self tRNA primers. J. Mol. Biol. 264:243–254.
58. Robert, D., M. L. Sallafranque-Andreola, B. Bordier, L. Sarih-Cottin, L. Tarrago-Litvak, P. V. Graves, P. J. Barr, M. Fournier, and S. Litvak.1990. Interactions with tRNA(Lys) induce important structural changes in human immunodeficiency virus reverse transcriptase. FEBS Lett.277:239–242. 59. Sprinzl, M., T. Hartmann, J. Weber, J. Blank, and R. Zeidler.1989.
Com-pilation of tRNA sequences and sequences of tRNA genes. Nucleic Acids Res.17(Suppl.):1–172.
60. Telesnitsky, A., and S. P. Goff.1997. Reverse transcriptase and the genera-tion of retroviral DNA, p. 121–160.InJ. M. Coffin, S. H. Hughes, and H. E. Varmus (ed.), Retroviruses. Cold Spring Harbor Laboratory Press, Cold Spring Harbor, N.Y.
61. Vicenzi, E., D. S. Dimitrov, A. Engelman, T.-S. Migone, D. F. J. Purcell, J. Leonard, G. Englund, and M. A. Martin.1994. An integration-defective U5 deletion mutant of human immunodeficiency virus type 1 reverts by elimi-nating additional long terminal repeat sequences. J. Virol.68:7879–7890. 62. Wakefield, J. K., S.-M. Kang, and C. D. Morrow.1996. Construction of a
type 1 human immunodeficiency virus that maintains a primer binding site complementary to tRNAHis. J. Virol.70:966–975.
63. Wakefield, J. K., A. G. Wolf, and C. D. Morrow.1995. Human immunode-ficiency virus type 1 can use different tRNAs as primers for reverse tran-scription but selectively maintains a primer binding site complementary to tRNALys3. J. Virol.69:6021–6029.
64. Whitcomb, J. M., B. A. Ortiz Conde, and S. H. Hughes.1995. Replication of avian leukosis viruses with mutations at the primer binding site: use of alternative tRNAs as primer. J. Virol.69:6228–6238.
65. Zhang, Z., S.-M. Kang, A. LeBlanc, S. L. Hajduk, and C. D. Morrow.1996. Nucleotide sequences within the U5 region of the viral RNA genome are the major determinants for an human immunodeficiency virus type 1 to maintain a primer binding site complementary to tRNAHis. Virology226:306–317. 66. Zhang, Z., S.-M. Kang, Y. Li, and C. D. Morrow.1998. Genetic analysis of
the U5-PBS of a novel HIV-1 reveals multiple interactions between the tRNA and RNA genome required for initiation of reverse transcription. RNA4:394–406.
10714 ABBINK ET AL. J. VIROL.