Copyright © 2000, American Society for Microbiology. All Rights Reserved.
Hypervariable Region 1 Sequence Stability during Hepatitis C
Virus Replication in Chimpanzees
STUART C. RAY,1* QING MAO,1ROBERT E. LANFORD,2SUZANNE BASSETT,2† OLIVER LAEYENDECKER,1YU-MING WANG,1‡ANDDAVID L. THOMAS1
Department of Medicine, Johns Hopkins University School of Medicine, Baltimore, Maryland 21205,1and
Department of Virology and Immunology, Southwest Regional Primate Research Center, Southwest Foundation
for Biomedical Research, San Antonio, Texas 782272
Received 18 August 1999/Accepted 21 December 1999
The putative envelope 2 (E2) gene of hepatitis C virus (HCV) contains a highly variable region referred to as hypervariable region 1 (HVR1). We hypothesized that this genetic variability is driven by immune selection pressure, rather than representing the accumulation of random mutations in a region with relatively little functional constraint. To test this hypothesis, we examined the E2 sequence of a human inoculum that was passaged through eight chimpanzees, which appear to have a replicative rate (opportunity for chance muta-tion) similar to that of humans. Acute-phase plasma samples from a human (the inoculum) and six of eight serially infected chimpanzees were studied. For each, 33 cloned cDNAs were examined by a combined hetero-duplex–single-stranded conformational polymorphism assay to assess quasispecies complexity and optimize selection of clones with unique gel shift patterns (clonotypes) for sequencing. The sequence diversity of HCV was significantly lower in the chimpanzees than in the humans, and during eight serial passages there was no change in the sequence of the majority clonotype from each animal examined. Similarly, the rates of protein sequence altering (nonsynonymous) substitution were lower in the chimpanzees than in the humans. These findings demonstrate that nonsynonymous mutations indicate selection pressure rather than being an inci-dental result of HCV replication.
An estimated 170 million people worldwide are infected with hepatitis C virus (HCV) (4). More than 80% of infections result in persistent viremia, which may be associated with chronic hepatitis, cirrhosis, liver failure, and hepatocellular cancer (3, 41, 44). HCV-associated disease is responsible for more than 10,000 deaths each year in the United States alone, and this mortality is expected to rise (8). Because of limitations of in vitro replication systems, studies of HCV pathogenesis have been limited to observation of natural infection of hu-mans and experimental infection of chimpanzees.
The chimpanzee is the best experimental model of HCV infection. HCV does not grow efficiently in tissue culture, and the only other candidate model is a tree shrew that is too small to permit adequate sampling with current technology (52). In chimpanzees, HCV replication has been demonstrated within days of experimental infection, and the potential for reinfec-tion has been demonstrated (14, 15, 37, 51). Serum HCV RNA levels and the extent of hepatic HCV involvement are similar in chimpanzees and humans, suggesting that replication rates are similar (1). The chimpanzee has also been used to test the infectivity of HCV clones and vaccine candidates. However, Bassett et al. recently demonstrated that the natural history of HCV infection in a cohort of experimentally infected chim-panzees differed from what is typically found in humans (5, 6). These chimpanzees had a higher rate of clearance of viremia, lower rate of antibody production to envelope proteins, and
lower rate of envelope amino acid change. Similarly, others have reported little change in E2 amino acid sequence in chimpanzees infected with molecular clones (24). In contrast, comparisons of E2 sequences in humans (both within and between infected subjects) reveal substantial heterogeneity, especially in the 27-amino-acid region at the N terminus of E2 that has been called hypervariable region 1 (HVR1).
HCV variants coexist in each infected individual as a swarm of genetically distinct but related variants, called a quasispecies (10–12, 23, 25). HCV sequences can shift rapidly during chronic human infection (20) and interferon therapy (31), con-sistent with the predicted behavior of a quasispecies in a rap-idly changing selective environment dominated by the immune system (40). Another prediction of the quasispecies model is that the master (or most common) sequence in the quasispe-cies will not change in a stable environment, while changes in minor variants will continue to occur. Evidence from immuno-suppressed individuals, in whom the pace of sequence varia-tion is reduced, suggests that this predicvaria-tion applies to HCV (7, 28, 42).
If HCV persists by escaping the immune response through sequence variation, then that sequence variation will reveal important characteristics of the immune response. If, however, persistence is due to other factors and sequence variation is simply the random product of an error-prone polymerase and a rapidly replicating virus (47), then sequence variation will always occur when HCV is allowed to replicate, and sequence variation will be uninformative except to define areas tolerant of sequence variation.
We hypothesized that despite an HCV replication rate sim-ilar to that in humans (37), reduced immune pressure in chim-panzees would result in limited sequence variation of HCV. To test this hypothesis, we studied the HCV quasispecies in a cohort of animals through which HCV was serially passed and in two animals with persistent infection. The serial passage * Corresponding author. Mailing address: Division of Infectious
Diseases, 720 Rutland Ave., Ross 1159, Baltimore, MD 21205. Phone: (410) 955-0349. Fax: (410) 955-7889. E-mail: [email protected].
† Present address: Department of Microbiology and Immunology, University of Texas Medical Branch, Galveston, TX 77555.
‡ Present address: Department of Infectious Diseases, Southwest Hospital, Third Military Medical University, Chongqing, Peoples’ Re-public of China.
3058
on November 9, 2019 by guest
http://jvi.asm.org/
experiment was initiated using a well-characterized strain of HCV (H77), obtained via plasmapheresis from a patient dur-ing acute HCV infection (5).
Using a highly sensitive method for detection of distinct variants, we determined the complexity of the quasispecies in each specimen and selected representative cloned cDNAs for sequence analysis. Sequences from these specimens revealed minimal changes in the envelope sequence examined on trans-fer from human to chimpanzee, in serial passage among chim-panzees, and during chronic infection of chimpanzees. These findings suggest that passage in chimpanzees does not affect the E2 sequence and that HCV replication does not inherently result in the accumulation of substitutions in the master se-quence.
MATERIALS AND METHODS
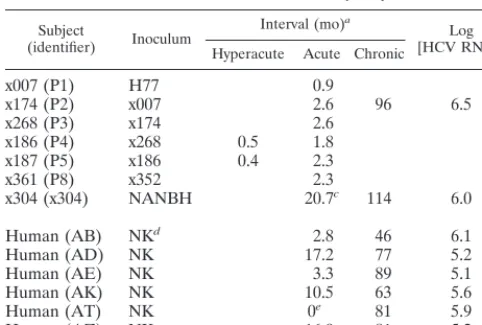
Study animals.As part of a study of non-A, non-B hepatitis (NANBH), chimpanzees were inoculated with human serum as follows (Table 1): animal x007 (passage 1 [P1]) was inoculated with 102.5chimpanzee infectious doses of
serum obtained via plasmapheresis from patient H in 1977, designated H77 (2, 17). Weekly assessments for rise in alanine aminotransferase allowed identifica-tion of the acute phase of infecidentifica-tion, and acute-phase serum was used to inoculate the next animal in the series. Animal x174 (P2) had persistent viremia, whereas the seven other animals in the passage had transient viremia (5).
In a separate experiment at another facility, animal x304 was inoculated with NANBH serum and developed persistent viremia. The inoculation date for x304 was recorded as “prior to November 1986,” and so November 1, 1986, was used for calculations that involved the date of inoculation.
Human subjects.Since 1988, a cohort of approximately 1,350 former and current injection drug users have been followed in Baltimore (46), including 43 subjects who acquired HCV infection during follow-up (45). The viral load trajectories and temporal sequence of HCV RNA and antibody detection for these subjects are described elsewhere (44). The six subjects chosen for this investigation continued to have detectable HCV RNA in the last specimen tested after at least 6 years of semiannual follow-up. The date of infection was esti-mated by calculating the midpoint between the date of the first specimen in which HCV RNA or antibody was detected and the date of the last prior sample in which evidence of HCV infection was not detected.
Storage of serum and testing for anti-HCV.Serum samples were immediately centrifuged, stored for less than 1 week at⫺20°C, and subsequently stored at
⫺70°C. They were tested for antibodies to HCV (HCV EIA [enzyme immuno-assay] 2.0; Ortho Diagnostics, Raritan, N.J.) and, if EIA positive, by strip im-munoblot assay (RIBA HCV 2.0; Chiron Corporation, Emeryville, Calif.) as previously described (45).
initially detected with a quantitative reverse transcriptase PCR assay (AMPLI COR HCV MONITOR; Roche Diagnostic Systems, Branchburg, N.J.), the lin-ear range of which was 500 to 500,000 copies per ml of serum in this and other laboratories (21, 34). For chimpanzees with persistent viremia, HCV RNA was quantitated using the COBAS AMPLICOR 2.0 detection system (Roche Diag-nostic Systems). In a comparison of these two methods using paired patient specimens and dilutions of H77, the COBAS method generally gave slightly higher values, but the difference was consistently less than a factor of 10 (A. Valsamakis, personal communication).
Envelope region amplification. HCV RNA characterization was based on examination of 33 cloned cDNAs spanning the 1,026-nucleotide (nt) region thought to encode envelope protein E1 and a segment of E2, including HVR1 (33). RNA was extracted from 100l of plasma or serum by using a QIAamp viral RNA mini kit as specified by the manufacturer (Qiagen, Valencia, Calif.). One-fifth of the extract was used to generate cDNA in a 20-l reaction at 37°C for 1 h with 20 U of Moloney murine leukemia virus reverse transcriptase (Perkin-Elmer, Foster City, Calif.) and first-round PCR reverse primer. The entire 20-l cDNA synthesis reaction was used for first-round PCR in a 25-l reaction containing 0.625 U Expand HF polymerase mixture (Boehringer Mann-heim, Indianapolis, Ind.), 1.5 mM MgCl2, 0.2 mM deoxynucleoside triphosphates
and 0.4M primers. The primers (and positions relative to the HCV-1 genome polyprotein [9]) were outer forward (493 to 518; 5⬘-GCAACAGGGAACCTTC CTGGTTGCTC-3⬘), outer reverse (1745-1723; 5⬘-GGGCAGDBCARRGTGT TGTTGCC-3⬘), inner forward (502-527; 5⬘-AACCTTCCTGGTTGCTCTTTCT CTAT-3⬘), and inner reverse (1527 to 1507; 5⬘-GAAGCAATAYACYGGRCC ACA-3⬘). Degenerate bases are indicated with standard International Union of Pure and Applied Chemistry codes. Ten microliters of the first reaction product was used as template for the inner nested PCR. Thermal-cycling conditions for both the inner and outer reactions were denaturation for 120 s at 94°C, followed by 35 cycles of 15 s at 94°C, 30 s at 65°C, and 60 s at 72°C (during the last 25 cycles, the elongation time was increased by 20 s per cycle).
Cloning of cDNA and complexity analysis of 33 cloned cDNAs by gel shift.The 1-kb HCV cDNA product was ligated into vector pCR 2.1 and used to transform INV␣F⬘cells (TA cloning kit; Invitrogen, Carlsbad, Calif.). Transformants were detected per manufacturer’s protocol, and cloning efficiency was⬎90%.
For each subject, the gel shift patterns of 33 cloned cDNAs were examined by amplifying a 452-bp region spanning HVR1 and by using a nonradioactive method that detects distinct variants within a sample by using a combination of heteroduplex analysis (HDA) and single-stranded conformational polymorphism (SSCP) on a single gel (HDA⫹SSCP) (48). Sequences obtained from the serial passage were analyzed by using a divergent variant from the chronic-phase specimen from P2 as the driver. A clonotype is defined as two or more cloned cDNAs that have indistinguishable patterns of electrophoretic migration by HDA⫹SSCP. In our earlier study, the mean (⫾standard deviation) genetic diversity of cloned cDNAs belonging to the same clonotype (intraclonotype diversity) was 0.6% (⫾0.9%), with 98.7% differing by less than 2% (48). The complexity of the quasispecies was characterized by the clonotype ratio, calcu-lated as the number of clonotypes divided by 33, the number of cloned cDNAs examined. The clonotype ratio therefore varies from 0.03 (homogenous) to 1 (highly complex).
Sequencing of representative cloned cDNAs.To examine each specimen’s quasispecies for trends in sequence variation, a subset of cloned cDNAs was identified for sequencing. For each subject, at least three cloned cDNAs were selected for sequencing based on gel shift patterns: two from the majority clono-type, one from each clonotype consisting of more than 10% of the 33 cloned cDNAs examined, and the cloned cDNA with the largest heteroduplex gel shift. Plasmid DNA was isolated from a 3.5-ml broth culture (High Pure plasmid isolation kit; Boehringer Mannheim) according to the manufacturer’s protocol. Sequences were determined from this DNA by using universal reverse primers with a PRISM version 2.1.1 automated sequencer (ABI, Foster City, Calif.). Sequences were assembled and edited in Sequencher (Gene Codes, Ann Arbor, Mich.) by a technician who was blinded to our hypotheses. Primer sequences were removed prior to analysis.
Phylogenetic analysis.Sequence alignments were randomly permuted 100 times by using the SEQBOOT program from the PHYLIP package version 3.572c (18, 19). DNA distance matrices were calculated by using the DNADIST program, maximum likelihood model, with a transition-to-transversion ratio of 4.25 (39). Permuted trees were generated using the NEIGHBOR program with random addition, and bootstrap values were obtained by using CONSENSE. Subtype reference sequences used for phylogenetic analysis had the following accession numbers (subtype designations in quotes remain controversial): 1a, AF009606 and M62321; 1b, D90208; 1c, D14853; 2a, D00944; 2b, D10988; 3a, D17763; 4a, Y11604; 5a, Y13184; 6a, Y12083; “7a,” D84263; “8a,” D84264; “9a,” D84265; “10a,” D63821; and “11a,” D63822. Nonsynonymous and synonymous substitution frequencies were calculated by the method of Nei and Gojobori (27).
Statistical analysis.After examination of the distribution of data, statistical inference was made by using the nonparametric Mann-Whitney test of medians. Subject
(identifier) Inoculum [HCV RNA]Log b
Hyperacute Acute Chronic
x007 (P1) H77 0.9
x174 (P2) x007 2.6 96 6.5
x268 (P3) x174 2.6
x186 (P4) x268 0.5 1.8
x187 (P5) x186 0.4 2.3
x361 (P8) x352 2.3
x304 (x304) NANBH 20.7c 114 6.0
Human (AB) NKd 2.8 46 6.1
Human (AD) NK 17.2 77 5.2
Human (AE) NK 3.3 89 5.1
Human (AK) NK 10.5 63 5.6
Human (AT) NK 0e 81 5.9
Human (AZ) NK 16.9 81 5.2
aInterval from date of inoculation (chimpanzees) or estimated date of
infec-tion (humans).
bFor chimpanzees, chronic-phase specimen, measured by the COBAS
meth-od; For humans, average of available specimens (n⫽5 to 12), excluding acute phase, measured by the Amplicor method.
cDate of inoculation for x304 estimated as described in Materials and
Meth-ods.
dNK, inoculum strain not known.
eHCV RNA was detected at first visit, prior to seroconversion.
on November 9, 2019 by guest
http://jvi.asm.org/
[image:2.612.53.294.83.245.2]FIG.
1.
Clonotype
distributions
and
alignment
of
predicted
amino
acid
sequences
from
chimpanzees
in
the
passage
experiment
(A)
and
for
animal
x304
(B
).
Clonotype
designations
were
assigned
sequentially
from
a
to
z
and
then
aa
to
ww
for
the
passage
and
from
A
to
U
for
x304.
In
the
upper
portion
of
each
panel,
the
numbers
below
the
clonotype
designations
indica
te
the
number
of
cloned
cDNAs
(out
of
33
assessed
for
the
animal
in
that
row)
with
HDA
⫹
SSCP
pattern
consistent
with
the
clonotype
indicated
in
that
column.
Periods
indicate
identity
to
the
reference
sequence
in
the
first
row.
The
asteris
k
in
sequence
P4_b
indicates
a
nonsense
mutation.
on November 9, 2019 by guest
http://jvi.asm.org/
[image:3.612.61.539.106.708.2]Nucleotide sequence accession numbers.The sequences obtained from chim-panzees were submitted to GenBank and assigned accession no. AF230416 through AF230459.
RESULTS
Amplification and HDAⴙSSCP analysis.From the six
ani-mals (P1, P2, P3, P4, P5, and P8) that comprised the serial eight passage lineage, the acute-phase plasma (collected 4 to 11 weeks after inoculation and representing the inoculum for the next numbered animal) was studied. Specimens collected within 2 weeks of inoculation (designated hyperacute) were also available for study from P4 and P5 (Table 1). Chronic-phase specimens from animals P2 and x304 were studied. An-imal P2 sustained chronic viremia 8 years after inoculation with x7 (H77) serum, and x304 sustained chronic viremia 10 years after inoculation with NANBH (HCV-1). The quasispecies complexity was examined by assessing 33 cDNA clones from each specimen using HDA⫹SSCP. The median clonotype ra-tio (range) of the acute phase of chimps, 0.27 (0.18 to 0.39), was similar (P⬎0.5) to that found in the first RNA-positive specimen from acutely infected humans, 0.39 (0.09 to 0.67).
Detection of persistent clonotypes through passages.Use of
a common cDNA clone to drive the HDA⫹SSCP gels permit-ted comparison of clonotypes among specimens. Among 198 cloned cDNAs from acute-phase specimens, 49 distinct pat-terns (clonotypes) were identified (Fig. 1). A subset of clono-types persisted during multiple passages in chimpanzees (Fig. 2). Clonotype a was the most frequently detected clonotype in animals P1, P3, P4, P5, and P8, representing 24 to 76% of cloned cDNAs. Clonotype b was detected in animals P1, P3, P4, and P5. Four clonotypes were observed in three animals each, and nine clonotypes were detected in two animals. For animals P1, P3, P4, P5, and P8, 91 to 94% of cloned cDNAs were observed in at least one other animal. In contrast, the six clonotypes observed for P2 were distinct from all others. An-imals P6 and P7 were not examined. Therefore, HDA⫹SSCP analysis indicated the persistence of clonotype a through chim-panzee passages P1 to P8, with the exception of animal P2, which was the animal that developed persistent viremia. In contrast to the persistence of clonotypes during serial passage, acute and chronic specimens from animals P2 and x304 shared
Increase in complexity during infection of chimps and
hu-mans.In each pair of specimens from the same chimpanzee,
the complexity of the quasispecies assessed using the clonotype ratio increased with time from inoculation or seroconversion, both during the acute phase (0.08 and 0.05 per month in animals P4 and P5) and also during the chronic phase (0.004 and 0.002 per month in animals P2 and x304). This was also true of four of the six humans who had clonotype ratio changes of⫹0.003,⫹0.005,⫹0.006, and⫹0.007. However, decreasing complexity was observed in subjects AT and AZ, who had per-month clonotype ratio changes of⫺0.007 and⫺0.004 (P⬎
0.1 for comparisons of clonotype ratios and rates of change). Of note, the infecting subtype of subjects AT and AZ (1b) differed from that of the other hosts (1a), and subject AT seroconverted for human immunodeficiency virus during the period of observation. Having found that human and chimpan-zee quasispecies were similar in the number of distinct variants in both acute and chronic infection, we compared the substi-tution rates.
Initial sequence analysis. By using HDA⫹SSCP to select
representative cloned cDNAs, 72 distinct cloned cDNAs were identified for sequencing. To determine the genetic identity of cDNA clones of the same clonotype, two representative se-quences representing the majority clonotype were compared for each specimen. There were nine differences among 11 pairs of 390-nt sequences (99.8% identity), and all but one difference represented sporadic substitutions (occurring only once in a set of related sequences) as defined by Smith et al. (38). The eight sporadic substitutions were probably artifactual, occurring at a frequency of 3.1 ⫻ 10⫺5 sporadic substitutions per site per PCR cycle, consistent with the misincorporation frequency of the thermostable polymerase and sequencing reactions (38) and similar to the rate others and we have observed (26, 33). For each majority clonotype, one of the two sequences was free of sporadic substitutions and was used in all subsequent anal-yses to represent that clonotype. In addition, no two cloned cDNAs identified as being distinct by HDA⫹SSCP analysis had identical sequences (data not shown). Therefore, the HDA⫹SSCP method is both highly sensitive and specific in detecting differences among cDNA clones, as previously re-ported (48). Since the cDNA sequence of one member of a clonotype represents the other members of that clonotype, the 72 cDNA sequences that we obtained represented 200 (72.2%) of the 363 cDNA clones examined.
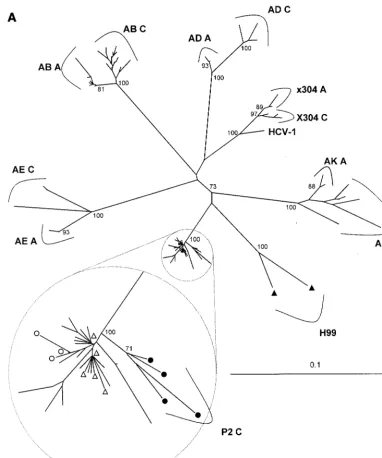
The most striking aspect of the chimpanzee HCV sequences was the low frequency of substitutions. A phylogenetic tree (Fig. 3A) shows that the acute-phase sequences form a cluster without monophyletic grouping of sequences from individual specimens representing eight sequential passages. Acute-phase sequences from P2 and inoculum strain H77 appear in this cluster. In contrast to the indistinct acute-phase sequences, chronic-phase sequences formed monophyletic clusters. Se-quences from animal x304 clustered with reference sequence HCV-1, agreeing with previous studies that suggested a close relationship between the inoculum of x304 and HCV-1 (6). This similarity is also apparent in Fig. 1.
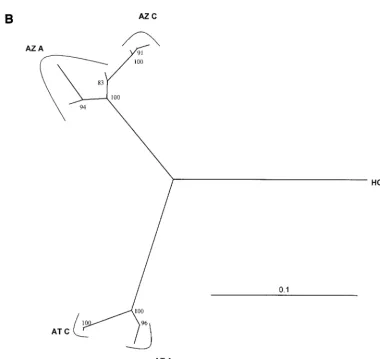
Comparison of acute- and chronic-phase sequences. The
[image:4.612.77.269.73.219.2]mean pairwise genetic distance between acute- and chronic-phase sequences was consistently lower in chimpanzees than in humans. This is illustrated by phylogenetic analysis (Fig. 3), which shows shorter branch lengths (lower genetic distance) between acute- and chronic phase sequences from chimpan-zees than those from humans. For both chimpanchimpan-zees with persistent viremia, the mean genetic distance between acute
FIG. 2. Polyacrylamide gel electrophoresis of HDA⫹SSCP assay as de-scribed in Materials and Methods. This photograph of a single gel shows the SSCP bands (upper panel) and HDA bands (lower panel) of cloned cDNAs chosen to illustrate the persistence of electrophoretic patterns. Clonotypes (groups of electrophoretically indistinguishable cloned cDNAs) were assigned sequential letter designations (a through z, then aa, bb, etc.). Chimpanzee iden-tifiers reflect the passage number (e.g., P2) and whether the specimen was hyperacute (H), acute (A), or chronic (C).
on November 9, 2019 by guest
http://jvi.asm.org/
and chronic sequences (interval of 8 to 10 years) was 0.013. In contrast, the mean genetic distance between acute- and chron-ic-phase sequences for human subject H (interval of 22 years) was 0.093, and in the other six humans (interval of 4 to 7 years) it was 0.044 (range, 0.028 to 0.054). Substitution rates calcu-lated from these data were lower for chimpanzees P2 and x304 (0.0018 and 0.0015 substitutions per site per year) than for human subject H (0.0042) and the other humans (mean, 0.0083; range, 0.0075 to 0.010) (P⫽0.055 by Mann-WhitneyU
test).
In humans, HVR1 is unlike the rest of the HCV genome
[image:5.612.111.493.72.529.2]because nonsynonymous substitutions are as frequent as syn-onymous substitutions (26, 39), but this was not the case in the chimpanzees in this study. The substitutions observed during chronic infection of chimpanzees were predominantly synony-mous, resulting in no change in the amino acid sequence, whereas they were predominantly nonsynonymous (resulting in changes in amino acid sequence) in humans. For chimpanzees P2 and x304, the nonsynonymous substitution frequencies (dN) were 0.011 and 0.012, the synonymous substitution frequencies (dS) were 0.025 and 0.023, anddN/dSratios were 0.44 and 0.52, respectively. For human subject H,dNwas 0.082,dSwas 0.13, FIG. 3. Neighbor-joining phylogenetic trees constructed from 372-nt envelope gene sequences of chimpanzees and human subjects infected with HCV subtype 1a (A) and 1b (B). The scale in genetic distance is indicated, and the number of 100 permuted trees supporting a clade is indicated when that proportion was greater than 70%. Identifiers correspond to those in Table 1, followed by the timing of the specimen (acute [A] or chronic [C], as defined in Materials and Methods). Triangles indicate sequences from subject H, and circles indicate sequences from animal P2 (open and filled symbols indicate acute and chronic specimens, respectively). Unlabeled taxa in the enlargement (inset) included sequences from P1, P3, P4, P5, and P8. All sequences from the six chimpanzees in the acute-phase passage series were included in the small encircled clades, without significant clustering. Trees that included 14 HCV subtype reference sequences confirmed clustering of sequences represented in panels A and B with subtypes 1a and 1b, respectively (data not shown).
on November 9, 2019 by guest
http://jvi.asm.org/
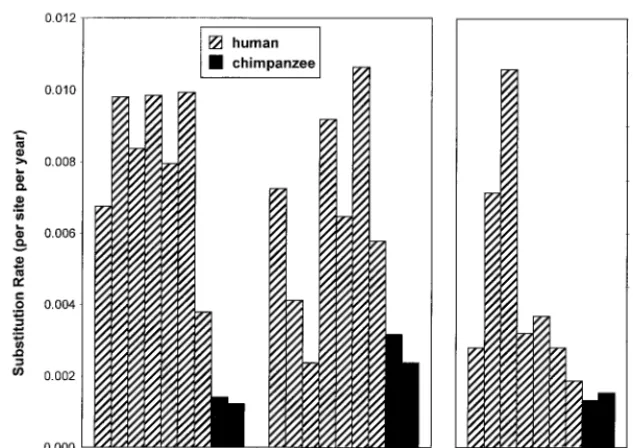
anddN/dSwas 0.65, whereas for the other six humans, mean (range)dN was 0.047 (0.024 to 0.060), mean (range)dS was 0.034 (0.017 to 0.057), and mean (range)dN/dSwas 1.7 (0.9 to 3.5). Rates derived from these data (Fig. 4) show that the nonsynonymous rate was higher for humans than for chimpan-zees (P⫽0.055 by Mann-WhitneyUtest), synonymous rates varied widely (P ⬎ 0.10), and the ratio of these rates was uniformly higher for humans than for chimpanzees (P⫽0.055 by Mann-WhitneyUtest).
Detailed analysis of sequences from serial passages. The
lack of change in the E2 sequence during eight sequential chimpanzee passages was striking. The majority acute-phase nucleotide sequence for each animal was identical to the ma-jority sequence for the inoculum (H77) strain, except for a single nucleotide change in the third HVR1 codon in animal P2. When all sequenced variants were included in the analysis, there was less than 0.009 weighted genetic sequence distance between any two sequential animals, and from P1 to P8 the weighted genetic distance was 0.0028. This limited variation was not focused in HVR1 as occurs in humans but was distrib-uted evenly across the region sequenced (data not shown).
Figure 5 shows the HVR1 amino acid sequence of the pre-dominant clonotype from each chimpanzee and human speci-men. It is notable for the persistence of the majority HVR1 sequence from H77 through all passages (except P2) and the persistence of the majority clonotype sequence in x304. The arginine substitution at residue 3 in all P2 acute-phase se-quences was not seen in any other chimpanzee specimen in this
study (Fig. 1) but has been described for other chimpanzees infected with H77 (29).
The substitutions observed during serial passage in chimpan-zees were predominantly synonymous (resulting in no change in the amino acid sequence). Between P1 and P8, dN was 0.0022 anddSwas 0.0050. For the intervening sequential pairs of animals, dNranged from 0.0020 to 0.0072 and dS ranged from 0.010 to 0.0180, except for the second passage (P1 to P2), for whichdSwas 0.0016. The ratio dN/dS, an indicator of im-mune selection pressure, ranged from 0.16 to 0.54 for all pas-sages except the second, for which it was 2.4. Of note, animal P2 was the only animal in the passage series to develop per-sistent viremia.
DISCUSSION
[image:6.612.115.496.70.429.2]In this investigation we demonstrated that HVR1 of two strains of HCV subtype 1a underwent very little sequence variation in chimpanzees, whether assessed during serial pas-sage or between acute and chronic phases. This is a striking finding since substantially greater sequence variation was noted in humans with similar HCV RNA levels, indicating the laboratory methods and number of replication cycles were sufficient to detect change. These results suggest that these HCV-infected chimpanzees exerted more negative selection (suppressing changes in the amino acid sequence of HCV) than humans or less positive selection (driving changes in the amino acid sequence), or both.
FIG. 3—Continued.
on November 9, 2019 by guest
http://jvi.asm.org/
Strong negative selection is not a likely explanation for the limited change in E2 protein sequences from chimpanzees, since this would imply that there are greater functional con-straints on the E2 protein in chimpanzees than in humans. However, a wide spectrum of E2 sequences can be detected in chimpanzees, suggesting that the observed stability of E2 se-quences is not due to restrictions imposed by the host.
Weaker positive selection is a more likely explanation for the observed sequence stability. A number of studies indicate that positive selective forces affect the HCV sequence. In par-ticular, HCV envelope sequence evolution has been correlated with antibody production (22, 35, 49), and lower rates of evo-lution have been observed in agammaglobulinemic subjects. In addition, HVR-specific antibodies may neutralize HCV infec-tivity (16, 36, 53). van Doorn and coworkers also demonstrated the temporal correlation of anti-HVR antibody production
and amino acid changes in chimpanzees (43). In this regard, it is important to note that the sequence stability observed in this study through the lineage of chimpanzees represented samples taken before the measured humoral and cellular immune re-sponse.
The supposition that positive selection drives sequence evo-lution for HCV is supported by studies of other RNA viruses. There have been a number of investigations correlating cyto-lytic T-lymphocyte responses with human immunodeficiency virus sequence variation (32, 50). In addition, recent data cor-relate nonsynonymous sequence changes with escape from dominant cytolytic T-lymphocyte responses in chimpanzees ex-perimentally infected with simian immunodeficiency virus (13). These data indicate that replication alone does not drive amino acid mutation in a quasispecies.
[image:7.612.144.461.74.298.2]The HDA⫹SSCP analysis used in this investigation reveals FIG. 4. Nonsynonymous rate, synonymous rate, and nonsynonymous/synonymous (Nonsyn/Syn) rate ratio for seven humans and two chimpanzees with chronic HCV viremia. In each panel, the humans and chimpanzees are presented in the order AB, AD, AE, AK, AT, AZ, H, P2, and x304.
FIG. 5. Alignment of deduced amino acid sequences of HVR1, one sequence for each of the majority clonotypes detected in each specimen from chimpanzees and humans infected with HCV. Periods indicate identity at that position with the reference sequence above. To the left of each sequence, the host is identified, followed by the timing of the specimen (acute [A] or chronic [C], as defined in Materials and Methods), followed by numbers indicating the number of cloned cDNAs (out of 33 total) represented by each clonotype. The x304 sequences are compared to HCV-1 because the inoculum for x304 was probably related to HCV-1.
on November 9, 2019 by guest
http://jvi.asm.org/
swarm of distinct but related variants. The number of variants (complexity) was similar to that found in acute human HCV infection. In each animal there was a master (most commonly observed) sequence representing 24 to 85% of the 33 variants examined, accompanied by minor variants which were nearly always (⬎90%) found in another animal, usually the prior or subsequent animal in the passage (Fig. 2), except for animal P2. The master sequence (represented by clonotype a in Fig. 2) did not change during the eight passages, except for a single nonsynonymous substitution in the one animal (P2) that de-veloped persistent viremia. The inoculum for P3 was obtained from P2 18 days prior to the P2 specimen available for this study. Therefore, explanations for the reappearance of clono-type a in P3 include (i) the mutation occurred or became dominant during the 18-day interval, (ii) a reversion mutation occurred in P3, and (iii) clonotype a was present in P2 below the level of detection but was preferentially expanded in P3.
Our findings are limited to the envelope gene region exam-ined. It is possible that significant changes occurred in other regions during the passage experiments or during chronic in-fection. Nonetheless, full-genome sequence data from 15-month follow-up of two chimpanzees infected with a molecular HCV clone (constructed from H77 sequence) also found little variability in the N terminus of E2 (24). In that study, one animal had a single amino acid substitution in HVR1 at week 51, despite readily detectable anti-HVR1 in both animals.
Although this study involves a relatively large number of chimpanzees, only two strains of HCV were assessed: H77 and NANBH (similar to HCV-1). In addition, both animals with chronic viremia (P2 and x304) received serum from an infected chimpanzee rather than a (possibly more complex) human inoculum. Other studies of chimpanzees infected with different HCV strains or human serum have shown higher rates of substitution (30, 43), while 1-year follow-ups of chimpanzees infected with HCV molecular clones have demonstrated lower substitution rates in E2 (24; J. Bukh, M. Yanagi, S. U. Emer-son, and R. H. Purcell, Abstr. Fifth International Meeting on Hepatitis C Virus and Related Viruses: Molecular Virology and Pathogenesis, abstr. 28, 1998). These different rates could be due to a variety of factors, including differences in the complexity of the inoculum, duration of follow-up, or possibly the subtype. In the two studies with higher rates, the infecting subtype was 1b, while our study and the studies of molecular clones involved subtype 1a. Although too little is understood about the factors that affect sequence evolution, our data dem-onstrate that chronic HCV infection need not result in the accumulation of nonsynonymous mutations. This finding im-plies that immunodominant domains can be identified by ex-amination of nonsynonymous change in a quasispecies, an im-portant adjunct to existing measurements of the host-virus interaction.
As shown in Fig. 4, the nonsynonymous substitution rate observed for human subject H was intermediate between the rates of the other humans and those of the chimpanzees. Be-cause the H77 strain was the inoculum for animal P1, the low rate of substitution for animal P2 might be attributed to some characteristic of the H77 strain. However, this would not ex-plain the similar rate found in animal x304. In addition, the rate obtained in this study from patient H is based on a 22-year sampling interval, and saturation at nonsynonymous sites may have occurred (26).
The possibility of PCR contamination was carefully evalu-ated. Negative control samples were carried through every reaction and were consistently negative, and the highly diverse
majority clonotype sequences from each chimpanzee was con-sistently identical to H77, the minority clonotype sequences consistently revealed differences, in accordance with a quasi-species rather than H77 contamination. In addition, the same minor variants were frequently observed in sequential animals in the passage (Fig. 3B). If these differences had been due to misincorporations during amplification and cloning, a more random distribution of variants would have been expected.
Because we found that dN/dS is low in chimpanzees and strong negative selection is unlikely, these results indicate that positive selection pressure on the HCV quasispecies is weaker in chimpanzees than in humans. These findings have important implications for interpretation of data obtained using the chimpanzee model of HCV infection, and they support the prediction that a quasispecies under reduced selective pressure will undergo reduced change in the master sequence.
ACKNOWLEDGMENTS
This study was supported by NIH grants IU19 AI-40035 and P51 RR13986. The ALIVE cohort is supported by NIH grants DA04334 and DA08009.
We thank Harvey Alter for generously providing sera from subject H and John Ticehurst for many helpful discussions.
REFERENCES
1.Agnello, V., G. Abel, G. B. Knight, and E. Muchmore.1998. Detection of widespread hepatocyte infection in chronic hepatitis C. Hepatology28:573– 584.
2.Alter, H. J., R. H. Purcell, P. V. Holland, and H. Popper.1978. Transmissible agent in non-A, non-B hepatitis. Lanceti:459–463.
3.Alter, M. J., H. S. Margolis, K. Krawczynski, F. N. Judson, A. Mares, W. J. Alexander, P. Y. Hu, J. K. Miller, M. A. Gerber, R. E. Sampliner, E. Meeks, and M. J. Beach.1992. The natural history of community acquired hepatitis C in the United States. N. Engl. J. Med.327:1899–1905.
4.Anonymous.1997. Hepatitis C: global prevalence. Weekly Epidemiol. Rec.
72:341–348.
5.Bassett, S. E., K. M. Brasky, and R. E. Lanford.1998. Analysis of hepatitis C virus-inoculated chimpanzees reveals unexpected clinical profiles. J. Virol.
72:2589–2599.
6.Bassett, S. E., D. L. Thomas, K. M. Brasky, and R. E. Lanford.1998. Viral persistence, antibody to E1 and E2 and HVR-1 sequence stability in hepatitis C virus-inoculated chimpanzees. J. Virol.73:1118–1126.
7.Booth, J. C., U. Kumar, D. Webster, J. Monjardino, and H. C. Thomas.1998. Comparison of the rate of sequence variation in the hypervariable region of E2/NS1 region of hepatitis C virus in normal and hypogammaglobulinemic patients. Hepatology27:223–227.
8.Centers for Disease Control and Prevention.1998. Recommendations for prevention and control of hepatitis C virus (HCV) infection and HCV-related chronic disease. Morbid. Mortal. Weekly Rep.47:1–39.
9.Choo, Q. L., K. H. Richman, J. H. Han, K. Berger, C. Lee, C. Gallegos, D. Coit, A. Medina-Selby, P. J. Barr, A. J. Weiner, D. W. Bradley, G. Kuo, and M. Houghton.1991. Genetic organization and diversity of the hepatitis C virus. Proc. Natl. Acad. Sci. USA88:2451–2455.
10. Domingo, E., E. Martinez-Salas, F. Sobrino, J. C. de la Torre, A. Portela, J. Ortin, C. Lopez-Galindez, P. Perez-Brena, N. Villanueva, and R. Najera.
1985. The quasispecies (extremely heterogeneous) nature of viral RNA ge-nome populations: biological relevance—a review. Gene40:1–8.
11. Domingo, E., D. Sabo, T. Taniguchi, and C. Weissmann.1978. Nucleotide sequence heterogeneity of an RNA phage population. Cell13:735–744. 12. Eigen, M.1971. Self organization of matter and the evolution of biological
macromolecules. Naturwissenschaften58:465–523.
13. Evans, D. T., D. H. O’Connor, P. Jing, J. L. Dzuris, J. Sidney, J. da Silva, T. M. Allen, H. Horton, J. E. Venham, R. A. Rudersdorf, T. Vogel, C. D. Pauza, R. E. Bontrop, R. DeMars, A. Sette, A. L. Hughes, and D. I. Watkins.
1999. Virus-specific cytotoxic T-lymphocyte responses select for amino-acid variation in simian immunodeficiency virus Env and Nef. Nat. Med.5:1270– 1276.
14. Farci, P., H. J. Alter, S. Govindarajan, D. C. Wong, R. Engle, R. R. Lesniewski, I. K. Mushahwar, S. M. Desai, R. H. Miller, and N. Ogata.1992. Lack of protective immunity against reinfection with hepatitis C virus. Eur. J. Biochem.258:135–140.
15. Farci, P., H. J. Alter, D. Wong, R. H. Miller, J. W. Shih, B. Jett, and R. H. Purcell.1991. A long-term study of hepatitis C virus replication in non-A, non-B hepatitis. N. Engl. J. Med.325:98–104.
on November 9, 2019 by guest
http://jvi.asm.org/
16.Farci, P., A. Shimoda, D. Wong, T. Cabezon, D. De Gioannis, A. Strazzera, Y. Shimizu, M. Shapiro, H. J. Alter, and R. H. Purcell.1996. Prevention of hepatitis C virus infection in chimpanzees by hyperimmune serum against the hypervariable region 1 of the envelope 2 protein. Proc. Natl. Acad. Sci. USA93:15394–15399.
17. Feinstone, S. M., H. J. Alter, H. P. Dienes, Y. Shimizu, H. Popper, D. Blackmore, D. Sly, W. T. London, and R. H. Purcell.1981. Non-A, non-B hepatitis in chimpanzees and marmosets. J. Infect. Dis.144:588–598. 18. Felsenstein, J.1985. Confidence limits on phylogenies: an approach using
the bootstrap. Evolution39:783–791.
19. Felsenstein, J.1989. PHYLIP—phylogeny inference package (version 3.2). Cladistics5:164–166.
20. Gretch, D. R., S. J. Polyak, J. J. Wilson, R. L. Carithers, Jr., J. D. Perkins, and L. Corey.1996. Tracking hepatitis C virus quasispecies major and minor variants in symptomatic and asymptomatic liver transplant recipients. J. Vi-rol.70:7622–7631.
21. Hadziyannis, E., M. W. Fried, and F. S. Nolte.1997. Evaluation of two methods for quantitation of hepatitis C virus RNA. Mol. Diagn.2:39–46. 22. Kato, N., Y. Ootsuyama, H. Sekiya, S. Ohkoshi, T. Nakazawa, M. Hijikata,
and K. Shimotohno.1994. Genetic drift in hypervariable region 1 of the viral genome in persistent hepatitis C virus infection. J. Virol.68:4776–4784. 23. Kato, N., Y. Ootsuyama, T. Tanaka, M. Nakagawa, T. Nakazawa, K.
Mu-raiso, S. Ohkoshi, M. Hijikata, and K. Shimotohno.1992. Marked sequence diversity in the putative envelope proteins of hepatitis C viruses. Virus Res.
22:107–123.
24. Major, M. E., K. Mihalik, J. Fernandez, J. Seidman, D. Kleiner, A. A. Kolykhalov, C. M. Rice, and S. M. Feinstone.1999. Long-term follow-up of chimpanzees inoculated with the first infectious clone for hepatitis C virus. J. Virol.73:3317–3325.
25. Martell, M., J. I. Esteban, J. Quer, J. Genesca, A. Weiner, R. Esteban, Guardia, and J. Gomez. 1992. Hepatitis C virus (HCV) circulates as a population of different but closely related genomes: quasispecies nature of HCV genome distribution. J. Virol.66:3225–3229.
26. McAllister, J., C. Casino, F. Davidson, J. Power, E. Lawlor, P. L. Yap, P. Simmonds, and D. B. Smith.1998. Long-term evolution of the hypervariable region of hepatitis C virus in a common-source-infected cohort. J. Virol.
72:4893–4905.
27. Nei, M., and T. Gojobori.1986. Simple methods for estimating the numbers of synonymous and nonsynonymous nucleotide substitutions. Mol. Biol. Evol.3:418–426.
28. Ni, Y. H., M. H. Chang, P. J. Chen, H. Y. Hsu, T. W. Lu, K. H. Lin, and D. T. Lin.1999. Decreased diversity of hepatitis C virus quasispecies during bone marrow transplantation. J. Med. Virol.58:132–138.
29. Ogata, N., H. J. Alter, R. H. Miller, and R. H. Purcell.1991. Nucleotide sequence and mutation rate of the H strain of hepatitis C virus. Proc. Natl. Acad. Sci. USA88:3392–3396.
30. Okamoto, H., M. Kojima, S. Okada, H. Yoshizawa, H. Iizuka, T. Tanaka, E. E. Muchmore, D. A. Peterson, Y. Ito, and S. Mishiro.1992. Genetic drift of hepatitis C virus during an 8.2-year infection in a chimpanzee: variability and stability. Virology190:894–899.
31. Pawlotsky, J. M., G. Germanidis, P. O. Frainais, M. Bouvier, A. Soulier, M. Pellerin, and D. Dhumeaux.1999. Evolution of the hepatitis C virus second envelope protein hypervariable region in chronically infected patients receiv-ing alpha interferon therapy. J. Virol.73:6490–6499.
32. Phillips, R. E., S. Rowland-Jones, D. F. Nixon, F. M. Gotch, J. P. Edwards, A. O. Ogunlesi, J. G. Elvin, J. A. Rothbard, C. R. Bangham, and C. R. Rizza.
1991. Human immunodeficiency virus genetic variation that can escape cy-totoxic T cell recognition. Nature354:453–459.
33. Ray, S. C., Y. M. Wang, O. Laeyendecker, J. Ticehurst, S. A. Villano, and D. L. Thomas.1998. Acute hepatitis C virus structural gene sequences as predictors of persistent viremia: hypervariable region 1 as decoy. J. Virol.
73:2938–2946.
34. Roth, W. K., J. H. Lee, B. Ru¨ster, and S. Zeuzem.1996. Comparison of two quantitative hepatitis C virus reverse transcriptase PCR assays. J. Clin. Mi-crobiol.34:261–264.
35. Shimizu, Y. K., M. Hijikata, A. Iwamoto, H. J. Alter, R. H. Purcell, and H. Yoshikura.1994. Neutralizing antibodies against hepatitis C virus and the emergence of neutralization escape mutant viruses. J. Virol.68:1494–1500. 36. Shimizu, Y. K., H. Igarashi, T. Kiyohara, T. Cabezon, P. Farci, R. H. Purcell, and H. Yoshikura.1996. A hyperimmune serum against a synthetic peptide corresponding to the hypervariable region 1 of hepatitis C virus can prevent viral infection in cell cultures. Virology223:409–412.
37. Shimizu, Y. K., A. J. Weiner, J. Rosenblatt, D. C. Wong, M. Shapiro, T. Popkin, M. Houghton, H. J. Alter, and R. H. Purcell.1990. Early events in hepatitis C virus infection of chimpanzees. Proc. Natl. Acad. Sci. USA
87:6441–6444.
38. Smith, D. B., J. McAllister, C. Casino, and P. Simmonds. 1997. Virus ‘quasispecies’: making a mountain out of a molehill? J. Gen. Virol.78:1511– 1519.
39. Smith, D. B., and P. Simmonds.1997. Characteristics of nucleotide substi-tution in the hepatitis C virus genome: constraints on sequence change in coding regions at both ends of the genome. J. Mol. Evol.45:238–246. 40. Steinhauer, D. A., and J. J. Holland.1987. Rapid evolution of RNA viruses.
Annu. Rev. Microbiol.41:409–433.
41. Tong, M. J., N. S. El-Farra, A. R. Reikes, and R. L. Co.1995. Clinical outcomes after transfusion-associated hepatitis C. N. Engl. J. Med.332:
1463–1466.
42. Toyoda, H., Y. Fukuda, Y. Koyama, J. Takamatsu, H. Saito, and T. Hay-akawa.1997. Effect of immunosuppression on composition of quasispecies population of hepatitis C virus in patients with chronic hepatitis C coinfected with human immunodeficiency virus. J. Hepatol.26:975–982.
43. van Doorn, L. J., I. Capriles, G. Maertens, R. DeLeys, K. Murray, T. Kos, H. Schellekens, and W. Quint.1995. Sequence evolution of the hypervariable region in the putative envelope region E2/NS1 of hepatitis C virus is corre-lated with specific humoral immune responses. J. Virol.69:773–778. 44. Villano, S. A., D. Vlahov, K. E. Nelson, S. Cohn, and D. L. Thomas.1999.
Persistence of viremia and the importance of long-term follow-up after acute hepatitis C infection. Hepatology29:908–914.
45. Villano, S. A., D. Vlahov, K. E. Nelson, C. M. Lyles, S. Cohn, and D. L. Thomas.1997. Incidence and risk factors for hepatitis C among injection drug users in Baltimore, Maryland. J. Clin. Microbiol.35:3274–3277. 46. Vlahov, D., J. C. Anthony, A. Mun˜oz, J. Margolik, D. D. Celentano, L.
Solomon, and B. F. Polk.1991. The ALIVE Study: a longitudinal study of HIV-1 infection in intravenous drug users: description of methods. J. Drug Issues21:759–776.
47. Wain-Hobson, S.1996. Running the gamut of retroviral variation. Trends Microbiol.4:135–141.
48. Wang, Y. M., S. C. Ray, O. Laeyendecker, J. R. Ticehurst, and D. L. Thomas.
1998. Assessment of hepatitis C virus sequence complexity by electrophoretic mobilities of both single- and double-stranded DNAs. J. Clin. Microbiol.
36:2982–2989.
49. Weiner, A. J., H. M. Geysen, C. Christopherson, J. E. Hall, T. J. Mason, G. Saracco, F. Bonino, K. Crawford, C. D. Marion, K. A. Crawford, et al.1992. Evidence for immune selection of hepatitis C virus (HCV) putative envelope glycoprotein variants: potential role in chronic HCV infections. Proc. Natl. Acad. Sci. USA89:3468–3472.
50. Wolinsky, S. M., B. T. Korber, A. U. Neumann, M. Daniels, K. J. Kunstman, A. J. Whetsell, M. R. Furtado, Y. Cao, D. D. Ho, and J. T. Safrit.1996. Adaptive evolution of human immunodeficiency virus-type 1 during the natural course of infection. Eur. J. Biochem.272:537–542.
51. Wyatt, C. A., L. Andrus, B. Brotman, F. Huang, D. H. Lee, and A. M. Prince.
1998. Immunity in chimpanzees chronically infected with hepatitis C virus: role of minor quasispecies in reinfection. J. Virol.72:1725–1730. 52. Xie, Z. C., J. I. Riezu-Boj, J. J. Lasarte, J. Guillen, J. H. Su, M. P. Civeira,
and J. Prieto.1998. Transmission of hepatitis C virus infection to tree shrews. Virology244:513–520.
53. Zibert, A., E. Schreier, and M. Roggendorf.1995. Antibodies in human sera specific to hypervariable region 1 of hepatitis C virus can block viral attach-ment. Virology208:653–661.