Copyright © 2002, American Society for Microbiology. All Rights Reserved.
Complement-Mediated Enhancement of Antibody Function for
Neutralization of Pseudotype Virus Containing Hepatitis C
Virus E2 Chimeric Glycoprotein
Keith Meyer,
1Arnab Basu,
1Craig T. Przysiecki,
2L. Martin Lagging,
3Adrian M. Di Bisceglie,
1Anthony J. Conley,
2and Ranjit Ray
1,4*
Departments of Internal Medicine1and Molecular Microbiology and Immunology,4Saint Louis University, St. Louis, Missouri;
Department of Virus and Cell Biology, Merck Research Laboratories, West Point, Pennsylvania2; and Department of
Infectious Diseases, Göteborg University, Göteborg, Sweden3
Received 1 October 2001/Accepted 15 November 2001
We previously reported a number of features of hepatitis C virus (HCV) chimeric glycoproteins related to pseudotype virus entry into mammalian cells. In this study, pseudotype virus was neutralized by HCV E2 glycoprotein-specific antibodies and infected human sera. Neutralization (50% reduction of pseudotype virus plaque formation) was observed with two human immunoglobulin G1 monoclonal antibodies (MAbs) at concentrations of between 2.5 and 10g/ml. A hyperimmune rabbit antiserum to an E2 hypervariable region 1 (HVR1) mimotope also exhibited an HCV E2 pseudotype virus neutralization titer of ⬃1/50. An E1 pseudotype virus used as a negative control was not neutralized to a significant level (<1/10) by these MAbs or rabbit antiserum to E2 HVR1. Since HCV probably has a lipid envelope, the role of complement in antibody-mediated virus neutralization was examined. Significant increases in the neutralization titers of the human MAbs (⬃60- to 160-fold higher) and rabbit antiserum to HVR1 mimotopes (⬃10-fold higher) were observed upon addition of guinea pig complement. Further, these studies suggested that complement activation occured primarily by the classical pathway, since a deficiency in the C4 component led to a significant decrease in the level of virus neutralization. This same decrease was not observed with factor B-deficient complement. We also determined that 9 of 56 HCV-infected patient sera (16%) had detectable pseudotype virus neutral-ization activity at serum dilutions of between 1/20 and 1/50 and that complement addition enhanced the neutralization activity of some of the HCV-infected human sera. Taken together, these results suggest that during infection, HCV E2 glycoprotein induces a weak neutralizing antibody response, that those antibodies can be measured in vitro by the surrogate pseudotype virus plaque reduction assay, and that neutralization function can be augmented by complement.
Hepatitis C virus (HCV) is a major causative agent of par-enterally transmitted hepatitis (6) and is associated with liver cirrhosis which may develop into hepatocellular carcinoma (4). The majority of HCV-infected individuals do not resolve the infection, leading to the development of chronic hepatitis. Ap-proximately 25% of infected individuals appear to clear HCV viremia without therapeutic intervention (5, 24). The mecha-nism leading to this natural resolution of HCV infection is unknown. The HCV genome is a linear, positive-sense, single-stranded RNA molecule of⬃9,500 nucleotides. It encodes a polyprotein precursor of⬃3,000 amino acids (7). This polypro-tein is cleaved by both host and viral proteases (17, 19) to generate several distinct polypeptides. The glycosylated virus polypeptides (E1 and E2-p7) comprise the viral envelope and facilitate virus entry into susceptible host cells.
Immunity to HCV infection is weak, and the reasons for this weak immunity are not clear. Although the immune response to the E1 glycoprotein has not been critically analyzed, some important observations have already been made regarding the E2 glycoprotein of HCV. Both E1 and E2 have N-terminal
hypervariable domains (29). Despite amino acid sequence vari-ability, the structure and global conformation of E2 hypervari-able region 1 (HVR1) are conserved (31). HVR1 contains basic residues at specific sequence positions. HVR1 also con-tains a sequence-specific immunological epitope which can in-duce antibodies restricted to the specific viral isolate (22, 45). HVR1 is probably the major site of HCV genetic drift, with amino acid substitutions in two overlapping B-cell epitopes. This scenario may lead to escape from neutralization by pre-existing HVR1 antibodies as changes in HVR anti-body specificity accompany HVR1 sequence shifts during the course of infection. An alternative suggestion is that anti-HVR1 reactivity is related more to the overall level of antibody response to HCV than to the HVR1 sequence itself (2). A correlation between the heterogeneity of the viral quasi-spe-cies and the quality of the immune response to HVR1 epitopes was not observed (2). On the contrary, an early appearance of antibody to the N terminus of E2 has been suggested as a possible indication of self-limiting HCV infection (49, 50). Binding of HCV to cells, as measured by reverse transcription (RT)-PCR, seems to parallel the in vitro infectivity of HCV for HPB-Ma cells. In this scenario, the neutralization of virus is mediated by isolate-specific antibodies recognizing the HVR1 region (39, 40). Indeed, in the chimpanzee infectivity model, ex vivo neutralization of HCV by patient sera and hyperimmune
* Corresponding author. Mailing address: Division of Infectious Diseases and Immunology, Saint Louis University, 3635 Vista Ave., St. Louis, MO 63110. Phone: (314) 577-8648. Fax: (314) 771-3816. E-mail: [email protected].
2150
on November 8, 2019 by guest
http://jvi.asm.org/
serum to E2 HVR1 further supports the importance of anti-body responses to this region (13, 14). However, the suggestion still remains that although the majority of antibodies are di-rected against E2 HVR1, the existence of high titers of HVR1-specific antibodies may not predict virus neutralization and may not be sufficient to block the binding of virus to human fibroblast cells (48).
The ability of antibody to neutralize the binding of E2 from genotype 1 is equally distributed among sera from patients infected with HCV genotypes 1, 2, and 3. An in vitro interac-tion between E1 and E2 and their role as a heterodimeric subunit for HCV infection have been suggested (11, 35). The E2 glycoprotein has been shown to bind human cells with a high affinity (36) and to interact with CD81 in vitro (33). Virus particles appear to use primarily the low-density lipoprotein (LDL) receptor for binding and entry (47). The specific mech-anism by which HCV particles interact with LDL or the LDL receptor is unknown.
In this study, we have generated a pseudotype virus by in-corporation of chimeric E1 or E2 in the viral envelope of a temperature-sensitive mutant of vesicular stomatitis virus (VSV) (ts045) and have shown infectivity of this virus in mam-malian cells. Earlier results with VSV or HCV pseudotype infectivity (26, 28) suggested that the HCV E1 and E2 glyco-proteins may be independently involved in virus attachment to and entry into a number of mammalian cells. Since VSV has a wide host range for replication, delivery of the virus genome into permissive cells by the pseudotype virus allows for an assessment of the ability of the envelope proteins to initiate infection. However, this does not indicate that HCV genome replication occurs in mammalian cells which allow pseudotype virus replication, as the entry of pseudotype virus will facilitate VSVts045 replication at the permissive temperature. Sera de-rived from chimpanzees immunized with homologous HCV envelope glycoproteins (genotype 1a) neutralized pseudotype virus infectivity. This finding suggested that antibody responses can be induced against HCV envelope glycoproteins. In this investigation, we have used the pseudotype virus as an alter-native to an in vitro infection system for HCV to study the neutralizing antibody response, the role of complement in an-tibody-mediated functions, and the neutralizing response in HCV-infected human sera.
MATERIALS AND METHODS
VSV-HCV pseudotype virus.Pseudotype virus was generated by using genomic sequences from HCV genotype 1a. The generation of VSV-HCV pseudotype virus from chimeric gene constructs encoding the putative ectodomains of the HCV E1 and E2 envelope glycoproteins and the transmembrane domain and
cytoplasmic tail of the VSV G protein by using VSVts045 in serum-free medium
has been described previously (26). In this study, a minor modification of the earlier method was adopted to avoid the use of recombinant vaccinia virus expressing T7 polymerase to facilitate the transient expression of the HCV glycoproteins. Stable transfectants of baby hamster kidney (BHK) cells
express-ing E1G or E2G were infected with VSVts045 and grown at 40.5°C
(nonper-missive temperature for VSVts045) for 20 h. Parallel mock-transfected BHK
cells were used as a VSVts045 infection control. Virus titer was determined by
a plaque assay at 32°C. The culture fluid was flash frozen and stored at⫺70°C in
aliquots, with each aliquot being thawed once for use.
Human MAbs.We selected two human monoclonal antibodies (MAbs) for study of the neutralizing efficacy for the E2 pseudotype virus. These two anti-bodies (CBH4G and CBH8C), of the immunoglobulin G1 (IgG1) subclass, recognize conformational epitopes of the HCV E2 glycoprotein. An unrelated MAb of the IgG1 subclass (R04) directed to cytomegalovirus p64 was also
included as an isotype-matched negitive control. These three human MAbs (18) were kindly provided by Steven K. H. Foung and Kenneth G. Hadlock (Stanford University, Stanford, Calif.). The HCV MAbs were generated from peripheral B cells of an individual infected with HCV genotype 1b (18). These two antibodies recognize epitopes which are common to HCV genotypes 1a, 1b, 2a, and 2b but do not appear to recognize identical epitopes. CBH8C has been found to inhibit HCV E2 binding to human CD81. The inhibitory activity of CBH8C may be due either to steric hindrance of E2 binding to CD81 or to binding of the MAb to E2, initiating a conformational change in E2 that prevents subsequent binding to CD81. CBH4G does not inhibit CD81 binding and recognizes a preformed E2-CD81 complex (18).
Preparation of HCV mimotopes, preparation of simultaneously conjugated peptide vaccine, and generation of rabbit antiserum.A mimotope peptide is a peptide that does not contain the primary amino acid sequence found in the hypervariable region (HVR) of E2 but reacts with antibody to the HVR. These peptides were first identified by panning a “biased” 27-mer phage display library with human HCV-positive sera known to recognize the HVR of E2. The mimo-tope peptides of HVR1 of HCV E2 have the following designations and sequences, as reported previously (34): D6, QTTTTGGQVSHATHGLTG LFSLGPQQK (norvaline); F78, QTHTTGGQAGHQAHSLTGLFSPGAKQN (norleucine); and H1, QTHTTGGVVGHATSGLTSLFSPGPSQK (aminobu-tyric acid).
These peptides were custom synthesized as linear sequences by solid-phase 9-fluorenylmethoxy carbonyl chemistry (Bio-Synthesis, Inc., Lewisville, Tex.). All
peptides were⬎90% pure, as judged by reversed-phase high-pressure liquid
chromatography. Each peptide consisted of 29 residues, including the 27-mer mimotope, a carboxy-penultimate unnatural residue, such as norleucine, and a
carboxy-terminal cysteine as a carboxamide.Neisseria meningitidisouter
mem-brane-protein complex (OMPC) (Merck Manufacturing Division, West Point, Pa.) was activated at pH 8.0 (10 mM HEPES buffer) by reaction with an excess
of the heterobifunctional cross-linker sulfosuccinimidyl 4-(N-maleimidomethyl)
cyclohexane-1-carboxylate (sSMCC; Pierce Chemical Co., Rockford, Ill.) for 2 h at 2 to 8°C. The activated OMPC was dialyzed against 10 mM HEPES (pH 8.0) or 1.0 mM morpholinepropanesulfonic acid (MOPS) (pH 7.1) for 24 h at 2 to 8°C
to removeN-hydroxysulfosuccinimide and excess sSMCC. The reaction
pro-ceeded with the addition of an equimolar mixture of the three thiol-containing mimotope peptides in order to generate a product consisting of three simulta-neously conjugated mimotope peptides. Nonreacting groups on the OMPC were
allowed to react withN-acetylcysteine, followed by dialysis against 10 mM
HEPES (pH 8.0) to remove excess unconjugated peptide andN-acetylcysteine.
The coupling load of each mimotope peptide for OMPC, expressed as a per-centage of total lysine, was calculated from the unnatural amino acid and lysine quantitation based on amino acid analysis of the acid-hydrolyzed (6 N HCl for 20 h at 110°C) conjugate. For the lysine-containing mimotopes, a correction was made.
Protein concentrations were determined by the Lowry assay with bovine serum
albumin as a standard. The percentage ofN.meningitidisprotein p42 was
deter-mined for the OMPC lot used to prepare conjugates. Sodium dodecyl sulfate-polyacrylamide gel electrophoresis analysis was performed on each preparation to provide evidence of covalent linkage of the mimotope peptides to protein components of OMPC. The coupling was in the range of 7 to 14%. The
peptide-OMPC conjugate was prepared with 300g of total protein in 10 mM HEPES
(pH 8.0) and was formulated in Freund’s complete and incomplete adjuvants. New Zealand White rabbits (2 to 2.5 kg) were injected at day 1 and at weeks 4 and 8. The animals were bled periodically to monitor the course of the immune responses and were bled to prepare serum (V7R4) at week 12.
Antibody responses to HVR peptides were determined against the homolo-gous immunizing peptides, heterolohomolo-gous mimotopes, or HCV HVR peptides by an enzyme-linked immunosorbent assay (ELISA) or by use of an ELISA with streptavidin capture of biotinylated peptide. The peptide ELISA titers (recipro-cal of the geometric mean titer [GMT]) for immune sera against the three
immunizing mimotopes ranged from 40,000 to 2.6⫻106. The peptide ELISA
titers for sera against the four most abundant HCV strain H HVR peptides (13, 14) ranged from 7,400 to 29,000. The peptide ELISA titer for sera against the most abundant HCV strain H HVR sequence was 21,000.
HCV-infected human sera.All patients had negative serological markers for human immunodeficiency virus (HIV) as well as a possible ongoing hepatitis B virus infection. Sera were tested with a second- or third-generation HCV enzyme immunoassay, and seropositivity was confirmed by a second- or third-generation recombinant immunoblot assay (Chiron Corporation, Emeryville, Calif.). Sero-typing was performed by using an HCV seroSero-typing 1-6 assay (Murex Biotech Limited, Dartford, United Kingdom). Virus load was determined by RT-PCR with a Cobas Amplicor HCV test (Roche Diagnostics, Branchburg, N.J.) with a
on November 8, 2019 by guest
http://jvi.asm.org/
detection limit of between 103and 106copies/ml. Alternatively, virus load was
measured with a Quantiplex HCV RNA kit (Chiron) as described by Fan et al. (12). HCV genotyping was done by using a multiplex PCR method with geno-type-specific primers as described by Widell et al. (46) or by using restriction fragment length polymorphism analysis as described by Fan et al. (12). Aliquots of the human sera were heat inactivated at 56°C for 30 min prior to use to destroy complement activity.
Neutralization of pseudotype virus infectivity.The infectivity of the E1 or E2 pseudotype virus depends upon the interaction of HCV glycoproteins with target cells (26). Once the VSV genome enters the cell after interaction of E1 or E2 with the cell surface, normal replication of VSV at the permissive temperature (32°C) occurs, and clear plaques are detected. We have used HCV E1 or E2 pseudotype virus and mammalian cell lines to test the neutralization of virus infectivity. Occasional pH alterations and clumping are observed when human hepatoma (HepG2) cells are used with an agar overlay in this assay (28). This situation leads to interference with distinct plaque formation and often to un-readable results. However, results obtained with BHK or human breast carci-noma (MCF7) cells are very consistent; thus, these cells represent an attractive cell substrate for the pseudotype plaque assay. Large-plaque formation of the pseudotype virus, particularly on BHK cells, offers the advantages of easy and convenient readout of the assay. All results are correlated for each of the three cell lines (BHK, MCF7, and HepG2), and variations are minimal (28). A plaque assay for virus infectivity and an assay for neutralization by antibodies were
performed at 32°C (permissive temperature for the VSVts045 mutant) with
BHK cells in serum-free medium (28).
To determine a neutralizing antibody response, pseudotype virus (⬃100 PFU)
was incubated with serial dilutions of antibodies or test sera at 37°C for 1 h. The
incubation mixture (total volume, 200l) was added to BHK cell monolayers in
individual wells of a six-well plate and incubated at 32°C for 1 h with intermittent tilting. Cells were washed two times with Dulbecco’s modified Eagle’s medium
(DMEM) and overlaid with a 1:1 mixture of 1.6% agarose and 2⫻DMEM
containing 2% (final concentration) fetal bovine serum. An additional agar overlay containing 0.0005% neutral red was added after 24 h to clarify plaque
counts. A reduction of⬃50% pseudotype PFU, compared to the control, was
considered the neutralization titer of the antibody or test serum.
To analyze any enhancement of plaque reduction via the complement path-way, after the virus-antibody mixture had been incubated at 37°C, guinea pig complement (Sigma, St. Louis, Mo.), optimized for use by previous titration, was added at a final dilution of 1/20. Samples were incubated for an additional 45 min at 37°C, added to cell monolayers, and incubated at 32°C for 1 h with intermittent tilting. Cells were overlaid with agar as described above. Virus neutralization by using guinea pig complement with or without the C4 component (Quidel, Santa Clara, Calif.) and human complement with or without factor B (Quidel) was also measured in this manner. When component-deficient complement sera were supplemented with their respective components, complement sera were adjusted to keep the levels of all other components constant with their controls. An unrelated human MAb (R04) of the same isotype or pooled normal rabbit sera were used as a negative control. Whenever applicable, pseudotype virus express-ing only the E1 glycoprotein was used as a negative control for comparison. The experiments were done at minimum in triplicate, and plaque numbers varied
within⫾5% in each independent experiment.
Complement-induced antibody-dependent cell damage.The Cyto Tox assay (Promega) was used to measure lactate dehydrogenase (LDH), a stable cytosolic enzyme that is released upon cell lysis (10). Released LDH in culture superna-tants was measured with a coupled enzymatic assay, which results in the conver-sion of a tetrazolium salt to a red formazan product. The intensity of the color formed is proportional to the number of lysed cells. In this assay, BHK cells expressing chimeric E2 glycoprotein on the cell surface were grown in triplicate wells in a 96-well cell culture plate. Cells were washed after overnight incubation and incubated with serial dilutions of E2-specific antibodies for 3 h at 37°C. Guinea pig complement was added at a predetermined final dilution (1/20) in the antibody-containing medium, and cells were incubated for an additional 1 h. Cells were centrifuged at 4°C, and the culture supernatant was separated to measure released LDH activity. Cell-associated LDH activity was measured by freeze-thaw and lysis following the manufacturer’s protocol (Promega). The intensity of color development was monitored by using an ELISA reader at a visible wavelength (492 nm).
RESULTS
E2-specific antibodies exhibit pseudotype virus neutraliza-tion activity.HCV-infected human sera have been tested for antibody responses for seroconversion or binding properties
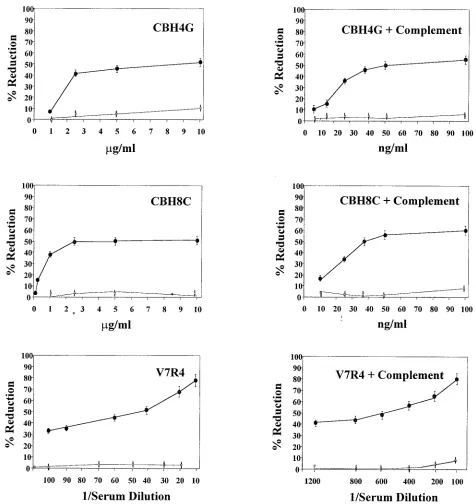
with a number of recombinant HCV proteins. However, inves-tigation into the functional role of the antibodies has not been convenient due to the lack of a dependable HCV replication system. Disease-specific mimotopes are efficient immunogenic mimics of the natural antigen, as they induce a specific immune response when injected into different animals. The peptide ligands for HVR1 are true mimotopes, as they do not neces-sarily share the same amino acid sequence as the original epitope but mimic its binding properties. In this study, we analyzed the roles of E2 glycoprotein-specific human MAbs and an HVR1 mimotope-specific rabbit antiserum in pseudotype virus neutralization. The human MAbs recognize epitopes which are shared among HCV genotypes 1a, 1b, 2a, and 2b. MAb CBH8C inhibits HCV E2 binding to human CD81, while CBH4G binds an epitope which does not inhibit this interaction (18). These two MAbs and the antiserum to an HVR1 mimotope exhibited dose-dependent neutralization of E2 pseudotype virus. CBH4G exhibited 50% virus reduction at ⬃5 to 10g of IgG1/ml, and CBH8C had a similar titer at⬃2.5 g/ml. However, none of these antibodies displayed a signifi-cantly increased level of plaque reduction, even at a higher concentration. On the other hand, V7R4 displayed better ef-ficacy (⬃80% reduction) at a lower dilution, and the reduction in pseudotype PFU gradually decreased with increasing dilu-tions, with a 50% reduction at an antiserum dilution of 1/50 (Fig. 1). A similar plaque reduction titer was observed with HepG2 or MCF7 cells. The overall efficacy of antibody-medi-ated neutralization was higher with the HVR1 mimotope-spe-cific antiserum than with the human MAbs. This difference could be due to the distinct nature of the antigenic sites on E2 recognized by the antibodies, their relationship to relevant E2 binding domains, or the relative affinities of the antibodies for their respective antigenic sites. Detectable neutralization ac-tivity was not found for pseudotype virus generated from cells expressing the E1 glycoprotein, included as a negative control, in parallel control experiments. Similarly, an unrelated MAb (R04) of the IgG1 subclass did not inhibit E1 or E2 pseudotype virus plaque formation.
Complement-mediated enhancement of antibody titers in pseudotype virus neutralization. Both positive and negative strands of HCV RNA in infected patient serum were observed to be associated with lipids and presumably lipid membrane particles (41). Complement is a major system of innate immu-nity which provides an initial line of defense against viral in-fection. To investigate the role of complement in neutraliza-tion, pseudotype virus was initially incubated with guinea pig complement at different dilutions and analyzed for infectivity by a plaque assay. Complement alone at a dilution ofⱖ1/20 was observed not to have an appreciable inhibitory effect on pseudotype virus plaque formation. Next, we determined whether serum complement has a role in the enhancement of antibody-mediated virus neutralization. The addition of com-plement to the reaction mixture led to a display of higher neutralization titers for E2 pseudotype virus, as indicated by the ⬃50% reduction in virus plaque formation (Fig. 1). A similar analysis with E1 pseudotype virus as a negative control failed to yield significant pseudotype virus neutralization activ-ity. These results further indicated that the human MAbs to HCV genotype 1b neutralize pseudotype virus containing the E2 glycoprotein from genotype 1a. Both of these IgG1 subclass
on November 8, 2019 by guest
http://jvi.asm.org/
antibodies exhibited a 60- to 160-fold increase in neutralization titers in the presence of complement, while rabbit antiserum to HVR1 mimotopes displayed an approximate 10-fold-higher neutralization activity. However, more complete neutralization (⬃100%) of pseudotype virus was observed at a higher con-centration of the human MAbs (5g) or rabbit antiserum to HVR1 mimotopes (1/50 dilution) in the presence of
[image:4.587.53.529.75.579.2]comple-ment (data not shown in Fig. 1). An unrelated human MAb (R04) used as an isotype-matched negative control or pooled normal rabbit sera did not exhibit any detectable inhibitory effect on pseudotype virus. Thus, the results of this study sug-gest a role for serum complement in the enhancement of virus neutralization and further supports its important biological role in the clearance of virus from the circulation.
FIG. 1. Neutralization of pseudotype virus generated from HCV E1 or E2 glycoprotein by human MAbs (CBH4G and CBH8C) and antiserum to an HVR1 mimotope (V7R4) in the absence or presence of guinea pig complement. Antibody dose-response curves for a reduction in E1 (open squares) or E2 (filled circles) pseudotype virus PFU in triplicate experiments are shown. Concentrations or reciprocal serum dilutions shown correspond to the individual antibodies or antiserum used. The errors bars for percent reduction at each antibody concentration or serum dilution represent standard deviations from three independent experiments.
on November 8, 2019 by guest
http://jvi.asm.org/
Complement facilitates antibody-dependent damage of E2 glycoprotein-expressing cells.We further investigated whether antibody-dependent activation of the complement pathway is effective in mammalian cells expressing chimeric HCV E1 or E2 glycoprotein on the BHK cell surface. Here, the cytotoxicity assay was used to measure LDH activity present in the cyto-plasm of intact cells. Quantitation in culture supernatants can be done only if they are lysed to release the LDH present in the cells. The MAbs and the antiserum to the E2 HVR1 mimotope were used to examine this effect. Both of the antibodies and the antiserum exhibited⬎26% cell cytotoxicity in the presence of guinea pig complement only with cells expressing E2 (Table 1). Similar experiments with E1-expressing BHK cells used as a negative control resulted in a much lower or negligible level of LDH release with anti-E2 antibodies in the presence of complement. The results of this study further suggest that antibody-dependent complement activation on HCV envelope glycoprotein-expressing cells leads to cell membrane damage.
MABs and an HVR1 mimotope-specific serum do not ex-hibit a synergistic effect on pseudotype virus neutralization.
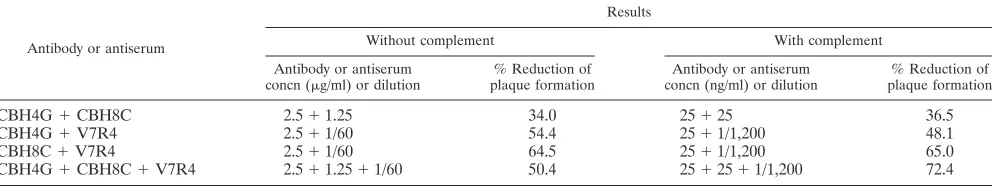
The mechanisms of virus neutralization are quite diverse, and multiple factors may contribute to antibody synergism. Here, we examined whether the antibodies and an antiserum exhib-ited a synergistic effect when used in combination at a range of ratios based on when they were equipotent for pseudotype virus neutralization (40 to 50%). For this purpose, the anti-bodies and the antiserum were tested in various combinations for their neutralizing effect on E2 pseudotype virus (Table 2). None of the antibodies or antiserum, when combined, signifi-cantly increased the titers of pseudotype virus neutralization in comparison to their individual effect. Interestingly, human MAb CBH4G displayed slightly higher E2 pseudotype virus reduction when used alone than when used along with CBH8C or antiserum to V7R4. A similar study did not suggest a
syn-ergistic effect on E2 pseudotype virus neutralization at lower concentrations of the antibodies in the presence of serum complement. In this study, the antibodies and antiserum ana-lyzed were unable to function in a synergistic fashion to in-crease plaque reduction efficiency despite their recognition of different antigenic sites.
[image:5.587.301.541.93.212.2]Complement-mediated enhancement of antibody function for pseudotype virus neutralization follows the classical path-way.We examined the pathway involved in complement-me-diated enhancement of antibody function. For this purpose, C4-deficient guinea pig serum and factor B-deficient human complement were used separately along with E2-specific anti-bodies to determine their effect on pseudotype virus neutral-ization. Factor B-deficient human complement did not result in a detectable change in antibody-mediated E2 pseudotype virus neutralization with CBH4G, CBH8C, or rabbit antiserum to the HVR1 mimotope. In contrast, the use of C4-deficient complement significantly decreased neutralization, and this ef-fect could be compensated for by the reintroduction of the C4 component into the reaction mixture (Table 3). These results suggested that the protective antiviral antibody response is augmented following the complement-dependent classical pathway. The subclass of an antibody may therefore affect the ability of the antibody to neutralize circulating HCV, as only selected subclasses have the ability to efficiently fix comple-ment. However, we do not exclude the possibility that the enhancing effect measured in the presence of whole serum as a source of complement is due in part to complement activa-tion by other pathways. Indeed, activaactiva-tion of the classical path-way will also generate the C3b fragment, which itself is ex-pected to trigger activation of the alternative pathway.
TABLE 1. Antibody- and complement-dependent cellular cytotoxicitya
Antibody (concn) or antiserum (dilution)
% Cytotoxicitybfor cells
expressing:
E1 E2
CBH4G (2.3 ng/ml) 8.3 26.1
CBH8C (1.9 ng/ml) 6.2 36.1
V7R4 (1/20) 4.6 33.9
aGuinea pig complement was used at a 1/20 dilution.
bDetermined as [(OD
492with antibody and complement⫺OD492with
com-plement)/OD492for total LDH activity]⫻100, where OD492is optical density at
[image:5.587.40.284.94.166.2]492 nm.
TABLE 2. E2 pseudoytpe virus neutralization by antibodies in the absence or presence of complement
Antibody or antiserum
Results
Without complement With complement
Antibody or antiserum
concn (g/ml) or dilution plaque formation% Reduction of concn (ng/ml) or dilutionAntibody or antiserum plaque formation% Reduction of
CBH4G⫹CBH8C 2.5⫹1.25 34.0 25⫹25 36.5
CBH4G⫹V7R4 2.5⫹1/60 54.4 25⫹1/1,200 48.1
CBH8C⫹V7R4 2.5⫹1/60 64.5 25⫹1/1,200 65.0
[image:5.587.45.541.635.729.2]CBH4G⫹CBH8C⫹V7R4 2.5⫹1.25⫹1/60 50.4 25⫹25⫹1/1,200 72.4
TABLE 3. Role of C4 complement component in E2 pseudotype virus neutralization
Antibody (concn) or
antiserum (dilution) Serum complement
Deficient component
suppliedb
%
Neutrali-zation
CBH4G (100 ng/ml) Complementa NA 54.6
CBH4G (100 ng/ml) Complement without C4 No ⬍5 CBH4G (100 ng/ml) Complement without C4 Yes 45.7 CBH 8C (100 ng/ml) Complementa NA 60.1
CBH 8C (100 ng/ml) Complement without C4 No ⬍5 CBH 8C (100 ng/ml) Complement without C4 Yes 48.4
V7R4 (1/100) Complementa NA 83.3
V7R4(1/100) Complement without C4 No ⬍5 V7R4 (1/100) Complement without C4 Yes 68.1
aAll complement components were present in guinea pig whole serum.
bNA, not applicable.
on November 8, 2019 by guest
http://jvi.asm.org/
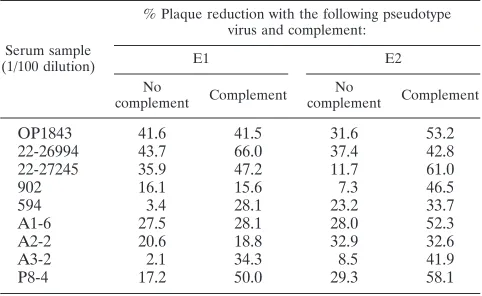
Pseudotype virus neutralization activity in HCV-infected human sera.Sera from 56 infected patients with different ge-notypes of HCV and virus loads were tested for neutralization of pseudotype virus generated separately from the E1 or E2 glycoprotein. Only 9 of the 56 sera (16%) exhibited neutral-ization at serum dilutions of between 1/20 and 1/50, with minor difference in titers between E1 and E2 pseudotype viruses (Table 4). Three of the sera tested (A1-6, A2-2, and A3-2) were taken during the acute phase, and the remaining six were from the chronic phase. All of these sera exhibited the ability to reduce pseudotype plaque number. The remaining 47 sera had titers of⬍1/10 against either of the pseudotypes used in this assay. The addition of complement in an antibody-virus incubation mixture enhanced the neutralization titers of some sera (Table 5), but the majority of the negative sera did not display a detectable change in titers. Complement-mediated enhancement of antibody titers was slightly more prevalent with E2 than with E1 pseudotype virus, and the reason for this difference is not clear at this time. Results from these experi-ments suggest that HCV infection elicits detectable neutraliza-tion activity for the pseudotype virus only in a small number of
HCV-infected individuals, independently of the infecting ge-notype and virus load.
DISCUSSION
Antibodies are responsible for neutralization of virus, and augmentation of neutralization or lysis of virions that have lipoprotein membranes is an important role of complement in antiviral immunity (25). The coating of the virion with C3 and, to a lesser extent, C4 produces the major effect in complement-mediated enhancement of virus neutralization. In this study, the neutralization of E2 pseudotype virus infectivity was ob-served with two different human MAbs recognizing distinct conformational epitopes and an antiserum directed to an HVR1 mimotope. Complement enhances the neutralization of antibody-bound virus by several mechanisms, including direct lysis of enveloped viruses (1, 8). The addition of complement enhanced the neutralization titer of the E2-specific antibodies by between ⬃10- and 160-fold. Complement-mediated anti-body-dependent cellular cytotoxicity was also evident when mammalian cells expressing chimeric HCV E2 glycoprotein on the cell surface were used. Complement-mediated enhance-ment of antibody titers was inhibited upon removal of the C4 component, indicating that complement activation occurs pri-marily through the classical pathway. Thus, mature extracellu-lar HCV may be efficiently inactivated and lysed by specific antibodies in the presence of complement. Additional results from our study indicated that the HCV glycoproteins consti-tute epitopes inducing neutralizing antibodies in only a small number of HCV-infected humans (16%); a number of the neutralization titers increased ⬃5-fold upon the addition of complement. Sera collected from HCV-infected patients from the onset of acute-phase disease were found to have detectable neutralization activity against pseudotype virus bearing HCV E1 or E2 envelope glycoprotein. Interestingly, in another study (L. M. Lagging et al., submitted for publication), it became more apparent that the percentage of acute-phase patient sera which had an identifiable titer was much greater (⬃80%) than that of chronic-phase sera. However, a majority of patients with chronic HCV infection failed to display detectable pseudotype virus neutralization activity, and serum from pa-tients with apparently resolved HCV infection had a neutral-ization titer which waned within a short time from the onset of infection (unpublished data). These results may have been due to poor immunogenicity of the virus envelope glycoproteins and/or generation of an inappropriate immune response in infected humans.
[image:6.587.43.284.92.235.2]A brief life span of circulating HCV-specific antibodies in recovered patients has also been observed by Takaki et al. (42). When seen, the neutralization activity of infected human sera appeared to be independent of the infecting HCV genotype or viral load, indicating the presence of common neutralizing epitopes on the pseudotype virus bearing E1 or E2. In earlier studies (20, 33), serum antibodies from patients chronically infected with different HCV genotypes were analyzed by a neutralization-of-binding (NOB) antibody assay with the E2 glycoprotein of HCV. Results from this study suggested that HCV infection did not elicit NOB in a significant number of cases, although low neutralization titers were detectable in the remaining infections. Low titers for NOB were also detected
TABLE 4. Neutralization of pseudotype viruses by HCV-infected human sera
Serum
sample Diseasephase Genotypeof virusa Viral load
Neutralization titer with the
following pseudotype:
E1 E2
OP1843b Chronic ND ⬍5⫻102 1/20 ⬍1/10 22-26994b Chronic ND 1.27⫻106 1/50 1/50 22-27245b Chronic ND 2.85⫻106 1/10 1/20
902c Chronic 2b 0.713 ⬍1/10 1/50
594c Chronic 1a 1.569 1/20 ⬍1/10
A1-6d Acute 1a 7⫻105 1/50 1/50
A2-2d Acute 3 Undetected 1/50 1/50
A3-2d Acute 3 2.5⫻103 1/20 1/20
P8-4d Chronic 3 1.5⫻104 1/50 1/50
aND, not determined.
bViral load was estimated with Roche RT-PCR (copies per milliliter).
cViral load was estimated with a Chiron Quantiplex HCV RNA kit
(mil-liequivalents per milliliter).
dViral load was estimated with the Roche Cobas Amplicor HCV test
(inter-national units per milliliter).
TABLE 5. Neutralization of pseudotype viruses by HCV-infected human sera in the presence or absence of complement
Serum sample (1/100 dilution)
% Plaque reduction with the following pseudotype virus and complement:
E1 E2
No
complement Complement complementNo Complement
OP1843 41.6 41.5 31.6 53.2
22-26994 43.7 66.0 37.4 42.8
22-27245 35.9 47.2 11.7 61.0
902 16.1 15.6 7.3 46.5
594 3.4 28.1 23.2 33.7
A1-6 27.5 28.1 28.0 52.3
A2-2 20.6 18.8 32.9 32.6
A3-2 2.1 34.3 8.5 41.9
P8-4 17.2 50.0 29.3 58.1
on November 8, 2019 by guest
http://jvi.asm.org/
[image:6.587.43.284.581.729.2]only in a small number of chimpanzees persistently infected with HCV genotype 1a. Although the NOB assay and our pseudotype virus neutralization assay are different, the latter clearly results in a lower (2.5- to 5-fold) end point. Our obser-vations are compatible with the results of Rosa et al. (36) showing that the ability to neutralize the binding of recombi-nant E2 derived from genotype 1 was evenly distributed among patients infected with genotypes 1, 2, and 3.
Mimotopes of HVR1 of HCV E2 can induce antibodies cross-reacting with a large number of viral variants (34). These antibodies, recognizing the most extensively variable N-termi-nal 27 amino acids of the E2 glycoprotein (45), were shown to be protective against viral infection by ex vivo neutralization prior to injection into chimpanzees (13, 14). A recent study with a virus bearing a deletion of HVR1 from E2 suggests that HVR1 may play a minimal role in the conformational changes of the glycoprotein, as the assembly of infectious virus still continues to occur and wild-type HCV without HVR1 is ap-parently attenuated (16). HVR1 positions are likely accessible at the surface of the E2 glycosylated product and are mainly involved in antigenicity and molecular interactions (31). The conservation of positively charged residues indicates that HVR1 is involved in interactions with negatively charged mol-ecules such as lipids, proteins, or glycosaminoglycans. Binding interactions of HCV E2 through multiple attachment cell sur-face molecules, including glycosaminoglycans, may confer a selective advantage, allowing the virus to easily attach to cells. In the present study, we observed a significant level of pseudotype virus neutralization (⬃80%) by an HVR1 mimo-tope-specific serum (V7R4) at an optimum concentration, even in the absence of serum complement. In contrast, rela-tively weak (⬃50 to 55%) pseudotype virus neutralization was obtained with the two human MAbs at their optimum concen-trations. These experimental findings further suggest that the binding of these two human MAbs can partially inhibit pseudotype virus entry, probably by steric hindrance. Alterna-tively, a relatively weak affinity of the antibodies for E2, com-pared to the much stronger affinity between E2 and host cell surface molecules, would not allow for complete inhibition of pseudotype virus plaque formation. The inhibitory activity; es-pecially that of CBH8C, could be due either to steric hindrance of E2 binding to CD81 or to binding of the antibody to E2 initiating a conformational change in E2 that leads to the prevention of subsequent binding to CD81. CBH8C has been observed to completely block the binding of E2 of HCV of genotypes 1a, 2a, 1b, and 2b, while being unable to bind to preformed E2-CD81 complexes, excluding those comprised of genotype 2a. This result will make it difficult to ascertain the true inhibitory nature of CBH8C as far as epitope specificity. The binding site in soluble form of HCV E2 interacting with CD81 is conformational (15), and recombinant human MAbs against conformational epitopes have been shown to inhibit the interaction of E2 with CD81 (3, 18). However, antibodies aimed at interfering with HCV binding to CD81 may not be completely neutralizing, as other molecules which interact with HCV exist on human cells (32). The nature of HCV E2 or its tertiary structure as a viral ligand may determine the functional relationship of the E2-CD81 interaction (30).
The ability of certain combinations of antibodies to display increased neutralization potency or reduced persistent
frac-tions and an enhanced neutralization rate has been noted before (37). Antibodies directed against different epitopes of the same antigen can bind synergistically. Synergistic binding of cocktails of antiviral antibodies directed to the same virus glycoprotein has been proposed to be responsible for increased neutralization. However, strict proportionalities between bind-ing affinity and either neutralization potency or neutralization rate are often not demonstrable (38). Possible exceptions are viruses, such as HIV, that appear to be neutralized primarily by prevention of attachment to different host cell receptors and coreceptors. In other cases, such as herpes simplex virus (HSV), where neutralization occurs primarily through postat-tachment mechanisms, epitope specificity appears to be impor-tant, given that different glycoproteins and specific domains within them may be crucial in virus penetration. In such a scenario, simultaneous inhibition of crucial determinants for HSV interaction with the cellular receptor could account for the synergistic properties of a cocktail of neutralizing antibod-ies. In the process of HSV penetration, different glycoproteins are believed to be sequentially recruited; this process would appear to produce synergistic interactions between neutraliz-ing antibodies directed to epitopes on different glycoproteins participating in the infectious process. The reason for the ab-sence of a synergistic effect of the antibodies in our pseudotype virus neutralization assay is not clear at this time. Interestingly, human MAb CBH4G displayed slightly higher E2 pseudotype virus reduction when used alone than when used along with MAb CBH8C or V7R4 antiserum, as noted earlier with HIV type 1 (23, 44). This result may have arisen from a number of mechanisms, including the interaction of different ligands with cell surface receptors and the relative affinities of the individ-ual antibodies for the respective antigenic sites. As a result, the binding of a higher-affinity antibody with E2 may induce a conformational change preventing the access of other antibod-ies and negating the cumulative effect. It is also plausible, in the case of complement-activating antibodies, that the anti-genic sites are situated in such a way that IgG antibodies may not build in the configuration which allows two Fc domains to come close enough together to initiate C1q activation of the classical complement pathway (25). In this manner, a combi-nation of two antibodies may antagonize their individual abil-ities to activate complement, leading to a decrease in pseudotype virus neutralization.
HCV may exist in the blood as free virus and virus com-plexed to antibodies. Anti-HCV antibody and HCV have been found in patients with essential mixed cryoglobulinemia (43). Cryoglobulins appear to occur in certain patients with lympho-proliferative, autoimmune, and infectious diseases. The immune response to HCV may include the development of cryoglobu-lins and autoimmunity (reviewed in reference 27), suggesting the possibility for molecular mimicry of the envelope proteins with host cell proteins. Because circulating immune complexes activate and consume complement, hypocomplementemia is the hallmark of immune complex-mediated diseases typified by mixed cryoglobulinemia and membraneous glomerulonephri-tis. Decreased hemolytic activity was detected in 45 (82%) of 55 apparently healthy persons with HCV viremia (21). It is likely that HCV-associated antigens form complexes with an-tibodies, consume complement, and induce decreased hemo-lytic activity in most symptom-free carriers of HCV. Among
on November 8, 2019 by guest
http://jvi.asm.org/
various viruses causing persistent infection, HCV seems to have a particularly strong capacity to induce hypocomple-mentemia. The geometric mean of hemolytic titers is much lower in patients with type C than in those with type B chronic liver disease. Total complement levels were decreased in seven of nine patients with chronic HCV disease, and C4 levels were decreased in six of seven patients (9).
Taken together, our results provide further support for the hypothesis that HCV E2 may play an independent role in virus attachment and/or entry, especially under situations such as those seen with pseudotype virus. Although this conclusion should be extrapolated with care due to the limitations of the particular model system used, the E2 chimeric glycoprotein appears to include epitopes that induce neutralizing antibod-ies, and their titers are enhanced by serum complement. Fur-ther studies on the role of serum complement during acute-phase infection may provide important information regarding HCV-related disease progression and pathogenesis.
ACKNOWLEDGMENTS
We gratefully acknowledge the interest and constructive criticisms of Robert B. Belshe during the entire course of this study. Our special thanks go to Steven K. H. Foung and Kenneth G. Hadlock for pro-viding the human MAbs and to Lin Cowick for preparation of the manuscript.
This research was supported by grants DK58023 and AI45250 from the National Institutes of Health.
REFERENCES
1.Akahane, Y., Y. Miyazaki, S. Naitoh, K. Takeda, F. Tsuda, H. Okamoto, K. Itoh, Y. Miyakawa, and M. Mayumi.1996. Cold activation of complement for monotoring the response to interferon in patients with chronic hepatitis
C. Am. J. Gastroenterol.91:319–327.
2.Allain, J.-P., Y. Dong, A.-M. Vandamme, V. Moulton, and M. Salemi.2000. Evolutionary rate and genetic drift of hepatitis C virus are not correlated with the host immune response: studies of infected donor-recipient clusters.
J. Virol.74:2541–2549.
3.Allander, T., K. Drakenberg, A. Beyene, D. Rosa, S. Abrignani, M. Hough-ton, A. Widell, L. Grillner, and M. A. A. Persson.2000. Recombinant human monoclonal antibodies against different conformational epitopes of the E2 envelope glycoprotein of hepatitis C virus that inhibit its interaction with
CD81. J. Gen. Virol.81:2451–2459.
4.Alter, H. J., R. H. Purcell, J. W. Shih, J. C. Melpolder, M. Houghton, Q. L. Choo, and G. Kuo.1989. Detection of antibody to hepatitis C virus in prospectively followed transfusion recipients with acute and chronic non-A,
non-B hepatitis. N. Engl. J. Med.32:1494–1500.
5.Alter, M. J., D. Kruszon-Moran, O. V. Nainan, G. M. McQuillan, F. Gao, L. A. Moyer, R. A. Kaslow, and H. S. Margolis.1999. The prevalence of hepatitis C virus infection in the United States, 1988 through 1994. N. Engl.
J. Med.341:556–562.
6.Choo, Q. L., G. Kuo, A. J. Weiner, L. R. Overby, D. W. Bradley, and M. Houghton.1989. Isolation of a cDNA clone derived from a blood-borne
non-A, non-B viral hepatitis genome. Science244:359–362.
7.Choo, Q. L., K. H. Richman, J. H. Han, K. Berger, C. Lee, C. Dong, C. Gallegos, D. Coit, R. Medina-Selby, P. J. Barr, et al.1991. Genetic organi-zation and diversity of the hepatitis C virus. Proc. Natl. Acad. Sci. USA
88:2451–2455.
8.Cooper, N. R.1991. Complement evasion strategies of microorganisms.
Im-munol. Today12:327–331.
9.Daoud, M. S., R. A. el-Azhary, L. E. Gibson, M. E. Lutz, and S. Daoud.1996. Chronic hepatitis C, cryoglobulinemia, and cutaneous necrotizing vasculitis. Clinical, pathologic, and immunopathologic study of twelve patients. J. Am.
Acad. Dermatol.34:219–223.
10.Decker, T., and M. L. Lohmann-Matthes.1988. A quick and simple method for the quantitation of lactate dehydrogenase release in measurements of cellular cytotoxicity and tumor necrosis factor (TNF) activity. J. Immunol.
Methods115:61–69.
11.Deleersnyder, V., A. Pillez, C. Wychowski, K. Blight, J. Xu, Y. S. Hahn, C. M. Rice, and J. Dubuisson.1997. Formation of native hepatitis C virus
glyco-protein complexes. J. Virol.71:697–704.
12.Fan, X., H. Solomon, J. E. Poulos, B. A. Neuschwander-Tetri, and A. M. Di Bisceglie.1999. Comparasion of genetic heterogeneity of hepatitis C viral
RNA in liver tissue and serum. Am. J. Gastroenterol.94:1347–1354.
13.Farci, P., H. J. Alter, D. C. Wong, R. H. Miller, S. Govindarajan, R. Engle, M. Shapiro, and R. H. Purcell.1994. Prevention of hepatitis C virus infection in chimpanzees after antibody-mediated in vitro neutralization. Proc. Natl.
Acad. Sci. USA91:7792–7796.
14.Farci, P., A. Shimoda, D. Wong, T. Cabezon, D. DeGioannis, A. Strazzera, Y. Shimizu, M. Shapiro, H. J. Alter, R. H. Purcell.1996. Prevention of hepatitis C virus infection in chimpanzees by hyperimmune serum against the hyper-variable region 1 of the envelope 2 protein. Proc. Natl. Acad. Sci. USA
93:15394–15399.
15.Flint, M., C. Maidens, L. D. Loomis-Price, C. Shotton, J. Dubuisson, P. Monk, A. Higginbottom, S. Levy, and J. A. McKeating.1999. Characteriza-tion of hepatitis C virus E2 glycoprotein interacCharacteriza-tion with a putative cellular
receptor, CD81. J. Virol.73:6235–6244.
16.Forns, X., R. Thimme, S. Govindarajan, S. U. Emerson, R. H. Purcell, and F. V. Chisari.2000. Hepatitis C virus lacking the hypervariable region 1 of the second envelope protein is infectious and causes acute resolving or
persistent infection in chimpanzees. Proc. Natl. Acad. Sci. USA21:13318–
13323.
17.Grakoui, A., C. Wychowski, C. Lin, S. M. Feinstone, and C. M. Rice.1993. Expression and identification of hepatitis C virus polyprotein cleavage
prod-ucts. J. Virol.67:1385–1395.
18.Hadlock, K. G., R. E. Lanford, S. Perkins, J. Rowe, Q. Yang, S. Levy, P. Pileri, S. Abrignani, and S. K. H. Foung.2000. Human monoclonal antibod-ies that inhibit binding of hepatitis C virus E2 protein to CD81 and recognize
conserved conformational epitopes. J. Virol.74:10407–10416.
19.Hijikata, M., N. Kato, Y. Ootsuyama, M. Nakagawa, and K. Shimotohno.
1991. Gene mapping of the putative structural region of the hepaitits C virus
genome by in vitro processing analysis. Proc. Natl. Acad. Sci. USA88:5547–
5551.
20.Ishii, K., D. Rosa, Y. Watanabe, T. Katayama, H. Harada, C. Wyatt, K. Kiyosawa, H. Aizaki, Y. Matsuura, M. Houghton, S. Abrignani, and T. Miyamura.1998. High titers of antibodies inhibiting the binding of envelope to human cells correlate with natural resolution of chronic hepatitis C.
Hepatology28:1117–1120.
21.Itoh, K., H. Tanaka, J-I Shiga, K. Hirakawa, Y. Akahane, F. Tsuda, H. Okamoto, Y. Miyakawa, and M. Mayamu.1994. Hypocomplemnetemia as-sociated with hepatitis C viremia in sera from voluntary blood donors. Am. J.
Gastroenterol.89:2019–2024.
22.Kato, N., H. Sekiya, Y. Ootsuyama, T. Nakazawa, M. Hijikata, S. Ohkoshi, and K. Shimotohno.1993. Humoral immune response to hypervariable re-gion 1 of the putative envelope glycoprotein (gp70) of hepatitis C virus.
J. Virol.67:3923–3930.
23.Kennedy, M. S., S. Orloff, C. C. Ibegbu, C. D. Odell, P. J. Maddon, and J. S. McDougal.1991. Analysis of synergism/antagonism between HIV-1 anti-body-positive human sera and soluble CD4 in blocking HIV-1 binding and
infectivity. AIDS Res. Hum. Retrovir.7:975–981.
24.Kenny-Walsh, E.1999. Clinical outcomes after hepatitis C infection from contaminated anti-D immune globulin. Irish Hepatology Research Group.
N. Engl. J. Med.340:1228–1233.
25.Lachmann, P. J., and A. Davies.1997. Complement and immunity. Immunol.
Rev.159:69–77.
26.Lagging, L. M., K. Meyer, R. J. Owens, and R. Ray.1998. Functional role of hepatitis C virus chimeric glycoproteins in the infectivity of pseudotyped
virus. J. Virol.72:3539–3546.
27.McMurray, R. W., and K. Elbourne.1997. Hepatitis C virus infection and
autoimmunity. Semin. Arthritis Rheum.26:689–701.
28.Meyer, K., Basu, A., and R. Ray.2000. Functional features of hepatitis C virus glycoproteins for pseudotype virus entry into mammalian cells.
Virol-ogy276:214–226.
29.Okamoto, H., M. Kojima, S. Okada, H. Yoshizawa, H. Iizuka, T. Tanaka, E. E. Muchmore, D. A. Peterson, Y. Ito, and S. Mishiro.1992. Genetic drift of hepatitis C virus during an 8.2-year infection in a chimpanzee: variability
and stability. Virology190:894–899.
30.Owsianka A., R. F. Clayton, L. D. Loomis-Price, J. A. McKeating, J. A., and A. H. Patel.2001. Functional analysis of hepatitis C virus E2 glycoproteins and virus-like particles reveals structural dissimilarities between different
forms of E2. J. Gen. Virol.82:1877–1883.
31.Penin, F., C. Combet, G. Germanidis, P.-O. Frainais, G. Deleage, and J.-M. Pawlotsky.2001. Conservation of the conformation and positive charges of hepatitis C virus E2 envelope glycoprotein hypervariable region 1 points to
a role in cell attachment. J. Virol.75:5703–5710.
32.Petracca, R., F. Falugi, G. Galli, N. Norais, D. Rosa, S. Campagnoli, V. Burgio, E. Di Stasio, B. Giardina, M. Houghton, S. Abrignani, and G. Grandi.2000. Structure-function analysis of hepatitis C virus envelope-CD81
binding. J. Virol.74:4824–4830.
33.Pileri, P., Y. Uematsu, S. Campagnoli, G. Galli, F. Falugi, R. Petracca, A. J. Weiner, M. Houghton, D. Rosa, G. Grandi, and S. Abrignani.1998. Binding
of hepatitis C virus to CD81. Science282:938–941.
34.Puntoriero, G., A. Meola, A. Lahm, S. Zucchelli, B. B. Ercole, R. Tafi, M. Pezzanera, M. U. Mondelli, R. Cortese, A. Tramontano, G. Galfre, and A. Nicosia. 1998. Towards a solution for hepatitis C virus hypervariability:
on November 8, 2019 by guest
http://jvi.asm.org/
mimotopes of the hypervariable region 1 can induce antibodies
cross-react-ing with a large number of viral variants. EMBO J.17:3521–3533.
35.Ralston, R., K. Thudium, K. Berger, C. Kuo, B. Gervase, J. Hall, M. Selby, G. Kuo, M. Houghton, and Q. Choo.1993. Characterization of hepatitis C virus envelope glycoprotein complexes expressed by recombinant vaccinia
viruses. J. Virol.67:6753–6761.
36.Rosa, D., S. Campagnoli, C. Moretto, E. Guenzi, L. Cousens, M. Chin, C. Dong, A. J. Weiner, J. Y. Lau, Q. L. Choo, D. Chien, P. Pileri, M. Houghton, and S. Abrignani.1996. A quantitative test to estimate neutralizing antibod-ies to the hepatitis C virus: cytofluorimetric assessment of envelope
glyco-protein 2 binding to target cells. Proc. Natl. Acad. Sci. USA93:1759–1763.
37.Sanna, P. P., F. Ramiro-Ibanez, and A. De Logu.2000. Synergistic
interac-tions of antibodies in rate of virus neutralization. Virology270:386–396.
38.Schofield, D. J., J. R. Stephenson, and N. J. Dimmock.1997.High and low efficiency neutralization epitopes on the haemagglutinin of type A influenza
virus. J. Gen. Virol.78:2441–2446.
39.Shimizu, Y. K., A. Iwamoto, H. Minako, R. H. Purcell, and H. Yoshikura.
1992. Evidence for in vitro replication of hepatitis C virus genome in a
human T-cell line. Proc. Natl. Acad. Sci. USA89:5477–5481.
40.Shimizu, Y. K., H. Igarashi, T. Kiyohara, T. Cabezon, P. Farci, R. H. Purcell, and H. Yoshikura.1996. A hyperimmune serum against a synthetic peptide corresponding to the hypervariable region 1 of hepatitis C virus can prevent
viral infection in cell cultures. Virology223:409–412.
41.Shindo, M., A. M. DiBisceglie, T. Akatsuka, T.-L. Fong, L. Scaglione, M. Donets, J. H. Hoofnagle, and S. M. Feinstone.1994. The physical state of the negative strand of hepatitis C virus RNA in serum of patients with chronic
hepatitis C. Proc. Natl. Acad. Sci. USA91:8719–8723.
42.Takaki, A., M. Wiese., G. Maertens, E. Depla., U. Seifert., A. Liebetrau, J. L. Miller, M. P. Manns, and B. Rehermann.2000. Cellular immune responses persist and humoral responses decrease two decades after recovery from a
single-source outbreak of hepatitis C. Nat. Med.6:578–582.
43.Tanaka, K., T. Aiyama, J. Imai, Y. Morishita, T. Fukatsu, and S. Kakumu.
1995. Serum cryoglobulin and chronic hepatitis C virus disease among
Jap-anese patients. Am. J. Gastroenterol.90:1847–1852.
44.Warrier-Vijh, S., A. Pinter, W. J. Honnen, and S. A. Tilley.1996. Synergistic neutralization of human immunodeficiency virus type 1 by a chimpanzee monoclonal antibody against the V2 domain of gp120 in combination with monoclonal antibodies against the V3 loop and the CD4-binding site. J.
Vi-rol.70:4466–4473.
45.Weiner, A. J., H. M. Geysen, C. Christopherson, J. E. Hall, T. J. Mason, G. Saracco, F. Bonino, K. Crawford, C. D. Marion, K. A. Crawford, M. Bru-netto, P. J. Barr, T., Miyamura, J. McHutchinson, and M. Houghton.1992. Evidence for immune selection of hepatitis C virus (HCV) putative envelope glycoprotein variants: potential role in chronic HCV infections. Proc. Natl.
Acad. Sci. USA89:3468–3472.
46.Widell, A., S. Shev, S. Mansson, Y. Y. Zhang, U. Foberg, G. Norkrans, A. Fryden, O. Weiland, J. Kurkus, and E. Nordenfelt.1994. Genotyping of hepatitis C virus isolates by a modified polymerase chain reaction assay using
type specific primers: epidemiological applications. J. Med. Virol.44:272–
279.
47.Wunschmann, S., J. D. Medh, D. Klinzmann, W. N. Schmidt, and J. T. Stapleton.2000. Characterization of hepatitis C virus (HCV) and HCV E2 interactions with CD81 and the low-density lipoprotein receptor. J. Virol.
74:10055–10062.
48.Zibert, A., S. Eckart, and M. Roggendorf.1995. Antibodies in human sera specific to hypervariable region 1 of hepatitis C virus can block viral
attach-ment. Virology208:653–661.
49.Zibert, A., W. Kraas, H. Meisel, G. Jung, and M. Roggendorf.1997. Epitope mapping of antibodies directed against hypervariable region 1 in acute
self-limiting and chronic infections due to hepatitis C virus. J. Virol.71:4123–
4127.
50.Zibert, A., H. Meisel, W. Kraas, A. Schulz, G. Jung, and M. Roggendorf.
1997. Early antibody response against hypervariable region 1 is associated
with acute self-limiting infections of hepatitis C virus. Hepatology25:1245–
1249.