Copyright © 1998, American Society for Microbiology
Genotype-Specific Complementation of Hepatitis Delta Virus
RNA Replication by Hepatitis Delta Antigen
JOHN L. CASEY*ANDJOHN L. GERIN
Division of Molecular Virology and Immunology, Georgetown University Medical Center, Rockville, Maryland 20852
Received 23 July 1997/Accepted 16 December 1997
Characterizations of genetic variations among hepatitis delta virus (HDV) isolates have focused principally on phylogenetic analysis of sequences, which vary by 30 to 40% among three genotypes and about 10 to 15% among isolates of the same genotype. The significance of the sequence differences has been unclear but could be responsible for pathogenic variations associated with the different genotypes. Studies of the mechanisms of HDV replication have been limited to cDNA clones from HDV genotype I, which is the most common. To perform a comparative analysis of HDV RNA replication in genotypes I and III, we have obtained a full-length cDNA clone from an HDV genotype III isolate. In transfected Huh-7 cells, the functional roles of the two forms of the viral protein, hepatitis delta antigen (HDAg), in HDV RNA replication are similar for both genotypes I and III; the short form is required for RNA replication, while the long form inhibits replication. For both genotypes, HDAg was able to support replication of RNAs of the same genotype that were mutated so as to be defective for HDAg production. Surprisingly, however, neither genotype I nor genotype III HDAg was able to support replication of such mutated RNAs of the other genotype. The inability of genotype III HDAg to support replication of genotype I RNA could have been due to a weak interaction between the RNA and HDAg. The clear genotype-specific activity of HDAg in supporting HDV RNA replication confirms the original categorization of HDV sequences in three genotypes and further suggests that these should be referred to as types (i.e., HDV-I and HDV-III) rather than genotypes.
Hepatitis delta virus (HDV) is a subviral human pathogen that increases disease severity in those infected with its helper, hepatitis B virus (29). The viral particle is composed of the single-stranded circular 1.7-kb HDV RNA, two forms of the viral protein, hepatitis delta antigen (HDAg), and hepatitis B surface antigen (2). HDAg binds specifically to HDV RNA (12, 20), and the short form, HDAg-S, is required for viral RNA replication (15). The mechanism of RNA replication is not known in detail, although several essential functional ele-ments of HDAg, including the regions involved in HDAg dimerization (amino acids 12 to 60 [[30, 36]) and RNA binding (amino acids 89 to 145 [18, 20]), have been identified.
Analysis of HDV isolates from around the world has indi-cated that there are at least three phylogenetically distinct genotypes with different geographic distributions and associ-ated disease patterns. Genotype I is the most widespread geo-graphically (26, 31), having been identified in isolates from North America, Europe, Africa, east and west Asia, and the South Pacific, and is associated with a broad spectrum of chronic disease (26). Genotype II has been found only in east Asia and may be responsible for some of the milder forms of HDV disease in that region (33, 34). Genotype III is exclu-sively found in northern South America, where HDV infection is associated with particularly severe disease (7, 10, 25). Se-quence divergence among the genotypes is as high as 40% over the entire RNA genome and 35% for the amino acid sequence of HDAg (7).
Thus far, the characterization of HDV genotypes has been based on phylogenetic analysis of sequences, and studies of HDV replication have been limited to clones of HDV geno-type I isolates, most of which are closely related. In this report,
we investigate replication of HDV genotype I and III clones in cultured cells to determine whether there might be functional as well as genetic differences between HDV genotypes.
MATERIALS AND METHODS
Plasmids for HDV genome expression.The genotype III expression plasmids
pHDVzIII(1) and pHDVzIII(2) were created from cloned PCR amplification products from the isolate Peru-1 (7). Briefly, amplification products spanning nucleotides (nt) 307 to 1290 and 1078 to 424 of the circular 1,677-nt genome were cloned into the plasmid vector pT7Blue (Novagen) to yield clones pNV1-2 and pNV3-1, respectively. The 1-kb PstI fragment from pNV1-1 was cloned into the
PstI site of pNV3-1 to yield the monomeric clone pNV3/1. The 1.7-kb KpnI
fragment from pNV3/1 was cloned between the KpnI sites of clone pNV1-3, a sister clone to pNV1-2, to yield pPeru-131.5. From clone pNV1-3, the 466-bp
SmaI fragment encompassing nt 522 to 988 of the Peru-1 genome was cloned into
the SmaI site of pGEM-3Zf(1) (Promega) to yield clone p293. The 1,677-bp
Esp3 I fragment from pPeru-131.5 (the Esp3 I site is at position 795 in the Peru-1 isolate) was cloned into the Esp3 I site of p293 to yield pPeru-131.2, which contained a duplication of both the genomic and antigenomic ribozyme autocleavage domains and encoded the short form of HDAg. The EcoRI-XbaI fragment from pPeru-131.2 was then inserted into the same sites of the plasmids pCMV2 and pCMV3 (8) to yield the expression plasmids pHDVzIII(1) and pHDVzIII(2), which direct the synthesis of antigenomic and genomic HDV RNA, respectively, in transfected cells.
Constructs for the expression of HDV RNA defective for HDAg production, HDAg(2), were created by site-directed mutagenesis with PCR primers con-taining the following mutations: for genotype I, an A was inserted between positions 1577 and 1578 (genotype I nucleotide numbering according to refer-ence 32), thereby introducing a stop codon and frameshift at codon 7; for genotype III, an A was inserted between positions 1579 and 1580 (genotype III nucleotide numbering according to reference 7), thereby introducing a stop codon and frameshift at codon 6. PCR-amplified fragments containing these mutations were inserted between the SstII and BglII sites of pCMV2-DC131.2 and pCMV3-DC131.2 (8) for genotype I, and between the Bsu36 I and XhoI sites of pHDVzIII(1) and pHDVzIII(2) for genotype III. The sequences of the PCR-amplified mutated segments were verified after cloning. Plasmids pHDVzI(2)Ag(2) and pHDVzIII(2)Ag(2) were constructed to produce genomic genotype I and genotype III RNA, respectively, in transfected cells, and pHDVzI(1)Ag(2) and pHDVzIII(1)Ag(2) were constructed to produce anti-genomic genotype I and genotype III RNA, respectively, in transfected cells.
Plasmid pHDVzIII(1)D-NR was constructed to express nonreplicating geno-type III antigenomic RNA in transfected cells. It is similar to the nonreplicating genotype I HDV antigenomic RNA expression construct pCMV3-DC-D131(A) * Corresponding author. Mailing address: Division of Molecular
Virology and Immunology, Georgetown University Medical Center, 5640 Fishers Lane, Rockville, MD 20852. Phone: (301) 881-2676. Fax: (301) 881-0810. E-mail: [email protected].
2806
on November 9, 2019 by guest
http://jvi.asm.org/
the same fragment in pHDAg-SzIII (see below) to yield pHDVzIII(1)D-NR, in which genotype III positions 1505 to 1677 and 1 to 90 have been deleted.
Plasmids for HDAg expression.The genotype I HDAg expression plasmid
pCMV-AgS, which encodes the short form of HDAg, is described elsewhere (28). For clarity, we refer to this construct here as pHDAg-SzI. The expression construct pHDAg-LzI, which encodes the long form of genotype I HDAg, was created by substituting the PstI-SalI fragment of the plasmid pGDC-1M2 (4), which contains the nucleotide C at position 1012, for the same fragment in pHDAg-SzI. The genotype III expression plasmid pHDAg-SzIII was obtained by substituting the 872-bp SstI-ApaI fragment from pPeru-131.2 for the same frag-ment in pHDAg-SzI. Plasmid pHDAg-SzIII contains positions 779 to 1651 from the Peru-1 isolate and includes the coding sequence for HDAg and the polyad-enylation signal. Expression plasmid pHDAg-LzIII, which encodes the long form of genotype III HDAg, was obtained by substituting the 310-bp PstI-ApaI frag-ment from pNV1-3, which contains the nucleotide C at position 1014, for the same fragment in pHDAg-SzIII.
Expression plasmids for HDAg-S from additional isolates were obtained by PCR amplification. Genotype I clones were obtained by ligating PCR-amplified fragments into the Ecl136 II site of plasmid pCMV3 (8). PCR amplification was performed with primers 5414 (26) and 6657 (59-CAGCAGTCTCCTCTTTACA GA-39; nt 1658 to 1638); conditions were as reported previously (6) except that the extension step was increased to 2 min and Pfu polymerase was used. Geno-type I isolates were from Italian patients I27 and I43, who were chronically infected with HDV (26), and U02, an American patient with chronic HDV infection (7). For genotype III, HDAg-coding sequences were amplified with primers 5414 and 7676 (59-GCTCCTTCCTCCTTAGGAGAGATAAG-39; nt 1650 to 1625). Fragments digested with Bsu36I (position 1635) and SmaI (posi-tion 988) were substituted for the same fragment in pHDAg-LzIII. Genotype III isolates were Peru 5 and 13, from Peruvian soldiers who experienced acute HDV infection while stationed at different base camps in the Amazon jungle (10), and P20 and P21, from two natives of the Amazon basin in Peru (provided by M. Sjo¨gren and A. Colichon). Clones were analyzed by restriction digestion and sequencing to determine the correct orientation relative to the cytomegalovirus promoter and to determine which form of HDAg (short or long) was encoded.
Chimeric HDAg expression plasmids. The chimeric constructs
pHDAg-SzIIIzIIIzI and pHDAg-SzIzIzIII were obtained by switching 0.7-kb NgoA IV frag-ments between pHDAg-SzI and pHDAg-SzIII. The NgoA IV site, located at position 1161 in the genotype I isolate, is identical in both isolates. Clones were screened by restriction digestion for the correct orientation. Plasmid pHDAgzIIIzIzI contained genotype III sequences from the beginning of the HDAg sequence up to the ApaI site in genotype I (nt 1380); it was created by a two-step procedure. Genotype III sequences between nt 1380 and 215 were amplified with primers 59-TTTCCTGCCTCGGGCCCTCTTCGCC-39and 59-T TTGCTGCCGATGGGCCCTCGGACCGGGGTC-39and cloned between the
ApaI sites of the genotype I plasmid pGDC-1 to yield pG{Peru/DC-1} (Apa);
clones were checked by restriction digestion for proper orientation. The primers used were specific for genotype III sequences, but internal ApaI sites were added; the ApaI site in the primer for the 1380 region is located such that the coding sequence is uninterrupted through this site when the genotype I and genotype III sequences are combined. The 601-bp BbsI-Esp3 I fragment from pG{Peru/DC-1}(Apa) was inserted between the same sites in plasmid pHDAg-SzIII to yield pHDAg-SzIIIzIzI. Plasmid pHDAg-SzIIIzIzIII, containing genotype I HDAg-S se-quences between the ApaI and NgoA IV sites and genotype III sese-quences elsewhere, was created by substituting the 708-bp NgoA IV segment from pHDAg-SzIII for the same fragment in pHDAg-SzIIIzIzI. Plasmid pHDAg-SzIzIIIzI, containing genotype III HDAg-S sequences between the ApaI and NgoA IV sites and genotype I sequences elsewhere, was created by a three-step pro-cedure. The region between nt 1380 and 883 was amplified with primers 59-GC GAAGAGGGCCCGGCAGGAAACCATG-39 and 5414 (26); the primer around 1380 contains an internal ApaI site which is located such that the coding sequence is uninterrupted through this site when genotype I and genotype III sequences are combined. The ApaI-PstI fragment of the PCR amplification product was substituted for the same fragment in plasmid pHDAg-SzIIIzIzI to give pHDAgP/C(Apa-Pst). The NgoA IV fragment of pHDAgP/C(Apa-Pst) was re-placed by the same fragment of pHDAg-SzI to give pHDAgP/C(Apa-Ngo); the
ApaI fragment of this plasmid was substituted for the same fragment in
pHDAg-SzI to yield pHDAg-SzIzIIIzI. Plasmid pHDAg-SzIzIIIzIII was created by substi-tuting the NgoA IV fragment of pHDAg-SzIII for that in pHDAg-SzIzIIIzI.
Transfections.Huh-7 cells were plated in six-well (35-mm-diameter) dishes
and transfected with 5mg of DNA per well by the calcium phosphate method as described previously (6). Transfections included the plasmid pSEAP2Control (Clontech), which was used to correct for transfection variability by monitoring
electroblotted onto Nytran (Boehringer Mannheim) membranes and fixed by UV illumination (Stratalinker; Stratagene, La Jolla, Calif.). Complete and even transfer of RNA samples to the membrane was verified by inspection of ethidium bromide-stained RNAs under UV illumination. Hybridization was for 16 to 36 h at 60°C in buffer containing 50% formamide, 1 M NaCl, 10% dextran sulfate, 1% sodium dodecyl sulfate (SDS), 0.5 mg of sheared salmon sperm DNA per ml, and hybridization probe (106cpm/ml; 53108cpm/mg). Blots were washed with 0.13 SSC (13SSC is 0.15 M NaCl plus 0.015 M sodium citrate) at 65°C and then exposed to film or quantified by radioanalytic imaging (InstantImager; Packard Instruments, Meriden, Conn.). Hybridization probes were monomeric genotype I or genotype III RNAs, as indicated, except where noted otherwise. Genotype I probes were used to detect genotype I RNAs, and genotype III probes were used to detect genotype III RNAs. Under the stringent hybridization and wash-ing conditions used, homologous hybridization is favored over heterologous hybridization because the sequences of genotypes I and III are only 60% iden-tical overall, and the longest contiguous segment of sequence identity is 35 nt. However, it is possible to detect genotype III RNA with a genotype I RNA probe, and vice versa. For purposes of quantitation, hybridization signal inten-sities were corrected for transfection efficiency by cotransfection of a reporter expression construct for secreted alkaline phosphatase (14).
Immunoblot analysis.Cell lysates were obtained by treatment with 2% SDS–
0.2 M Tris-HCl (pH 7.5)–1 mM EDTA and analyzed for hepatitis delta antigen by SDS-polyacrylamide gel electrophoresis in 12% acrylamide gels and immu-noblotting with human monoclonal anti-HD T1/39 as described elsewhere (27).
RESULTS
A replicating HDV genotype III cDNA clone. Analyses of HDV replication in cultured cells have thus far been limited to cDNA clones of HDV genotype I isolates. Two of the clones used (6, 15, 22) were derived from animals infected in related passage series and differed by about 1% (16, 32), a third (11, 23) was obtained from a genotype I isolate (24) that is about 10% different in nucleic acid sequence over the entire genome. Sequence variations among the genotypes, on the other hand, are as high as 40% (7), and inspection of sequence differences within some functional elements (such as the RNA editing site and the C terminus of the long form of HDAg) suggests that there could be important functional differences among the genotypes (3, 9). To compare the replication of genotypes I and III, we obtained a full-length cDNA clone from the pro-totype genotype III isolate Peru-1 (7). Expression constructs in which the cytomegalovirus promoter directs the synthesis of HDV RNA containing duplications of the ribozyme elements and encoding the short form of HDAg were created (Fig. 1). These constructs are similar to those used for analysis of HDV genotype I replication (6).
Following transfection of Huh-7 cells, the genotype III
ex-pression construct pHDVzIII(1) yielded replicating HDV
ge-notype III RNA, as indicated by the accumulation of both genomic (Fig. 2A) and antigenomic RNA (not shown) between 6 and 12 days posttransfection, even though only antigenomic RNA synthesis is directed by the cytomegalovirus promoter. The results shown are representative of numerous similar transfections in which either genomic or antigenomic RNA synthesis was directed by the transfected plasmids. In these experiments, the level of HDV RNA was determined by ra-dioanalytic imaging to be about two- to threefold lower for the transfected genotype III expression construct than for a similar genotype I expression construct. In addition to HDV RNA expression, HDAg was also detected in the transfected cells. Although only the short form of HDAg is encoded in the transfected constructs, both forms of HDAg are produced
on November 9, 2019 by guest
http://jvi.asm.org/
on November 9, 2019 by guest
http://jvi.asm.org/
(Fig. 2B) because of an RNA editing event that occurs during viral RNA replication (6, 21). Comparison of the relative levels of genotype I and III HDAg expression is difficult because of antigenic and structural variations. Curiously, genotype III HDAg-S migrated faster in the gel than did genotype I HDAg (Fig. 2C). Genotype III HDAg-L also migrated faster than its genotype I counterpart, but the difference in the migration rates was not as great. The different migration pattern was observed in numerous experiments in which either replicating HDV expression constructs or HDAg expression constructs were transfected (not shown).
Genotype-specific support of RNA replication by HDAg.In cultured cells, HDV RNA replication requires expression of the short form of HDAg (16). Thus, transfection of HDV cDNA constructs deficient in HDAg expression yields replicat-ing HDV RNA only if an expression construct for HDAg is also cotransfected and if the defect which abrogates HDAg expression leaves intact structural elements required for RNA replication (1, 15, 17, 35). To examine the ability of HDAg to support replication of genotype I and genotype III HDV RNAs, constructs for intracellular expression of RNAs defec-tive for HDAg synthesis were created (Fig. 1B). The defect was created by inserting a thymidine nucleotide in the cDNA at the beginning of the seventh and sixth codons, respectively, of the wild-type genotype I and genotype III expression constructs.
This insertion created a stop codon as well as a frameshift. A single-base insertion was used in order to minimize potential effects on the RNA secondary structure that could also affect the ability of the RNA to replicate (35).
Transfection of Huh-7 cells with HDAg(2) RNA expression
constructs but without cotransfected expression plasmids for HDAg yielded no evidence of HDV RNA replication, as de-termined by the inability to detect opposite-sense transcripts 6 days posttransfection (Fig. 3). As previously shown for similar
genotype I HDAg(2) expression constructs, RNA replication
was observed upon cotransfection of an expression construct for genotype I HDAg (Fig. 3). Antigenomic RNA was readily detectable 6 days after cotransfection of the defective genomic
RNA expression construct pHDVzI(2)Ag(2) with the HDAg
expression construct pHDAg-SzI, and genomic HDV was
ob-served after cotransfection of the defective antigenomic RNA
expression construct pHDVzI(1)Ag(2) with the same
geno-type I HDAg expression construct. The same result was ob-tained for genotype III: cotransfection of genotype III
HDAg(2) constructs with a genotype III HDAg expression
construct yielded high levels of replication of genotype III
HDAg(2) RNAs (Fig. 3). However, genotype III HDAg did
not support appreciable replication of genotype I HDAg(2)
RNAs, and genotype I HDAg supported replication of
geno-type III HDAg(2) RNAs at only about 5% of the level
ob-served with genotype III HDAg (Fig. 3). Thus, the ability of
HDAg to support replication of HDAg(2) RNA was genotype
specific.
[image:4.612.82.255.459.654.2]HDAg clones from different HDV isolates.The results pre-sented in Fig. 2 and 3 were obtained with clones derived from the prototype isolates of HDV genotypes I and III. To deter-mine whether the observed effects might be due to genotype-specific differences rather than a peculiarity of the two isolates used, additional HDAg expression clones were obtained by cloning PCR-amplified cDNAs. Three clones were obtained from additional unrelated genotype I isolates and four from
[image:4.612.307.546.572.658.2]FIG. 2. RNA replication and HDAg production following transfection with clones of genotype I and genotype III HDV. Huh-7 cells were transfected with pCMV3-DC131.2 (left) or pHDVzIII(1) (right), which express 1.2 unit-length genotype I and genotype III antigenomic RNA, respectively. Cells were har-vested and RNA and proteins were isolated on days 6 (lanes 1 and 4), 9 (lanes 2 and 5), and 12 (lanes 3 and 6). (A) Northern blot analysis of genomic HDV RNA; (B) immunoblot analysis; (C) second run of samples from lanes 3 and 6 of panel B in adjacent lanes in the same gel.
FIG. 3. Genotype-specific support of RNA replication by HDAg-S of geno-types I and III. Huh-7 cells were transfected with indicated constructs for the expression of genomic (left) or antigenomic (right) HDV RNA defective for HDAg production. Cells were also cotransfected with the plasmid vector pCMV3 (lanes2) or plasmids pHDAg-SzI (lanes I) and pHDAg-SzIII (lanes III), which express genotype I and III HDAg, respectively. RNAs were isolated 6 days posttransfection and analyzed by Northern blotting. Hybridization probes de-tected antigenomic RNA (left) or genomic RNA (right).
expression constructs for HDAg.
on November 9, 2019 by guest
http://jvi.asm.org/
additional unrelated genotype III isolates. Within a 357-bp region (nt 908 to 1265) corresponding to the C-terminal half of the HDAg-coding region, extents of sequence divergence among these genotype I and genotype III isolates were from 6 to 9% and from 4 to 8%, respectively (4, 10, 26, 31). Upon cotransfection of Huh-7 cells, all four genotype I HDAg clones
supported replication of HDAg(2) genotype I RNA, and all
five genotype III HDAg clones supported replication of
HDAg(2) genotype III RNA (Fig. 4). However, none of the
HDAg clones was able to support replication of HDAg(2)
RNA of the other genotype at more than 5% of the level supported by the prototype HDAg clones of the same geno-type (Fig. 4). These results were verified in three separate transfection experiments. Variations in transfection efficiency, as determined by analysis of a cotransfected reporter gene (14), were less than twofold and could not account for the different levels of RNA replication.
Interestingly, the level of RNA replication supported varied considerably among the HDAg clones, particularly for those from genotype I. Isolates I27 and I43 were 5-fold less active and 10-fold more active, respectively, than the HDAg clone from the prototype (Fig. 4). Variations in the level of HDAg expression were less than 25%, as determined by immunoblot analysis (not shown), and do not likely explain the differences in the levels of genotype I RNA replication supported by the genotype I HDAg clones. The low activity of the I27 clone was not likely due to deleterious mutations introduced by misin-corporation during PCR amplification prior to cloning, be-cause a second clone from a separate PCR amplification pro-duced the same result. Isolates U02 and I27 are more closely related to the prototype (6.5 and 7.3% divergence, respec-tively) than is isolate I43 (9% divergence), suggesting that genetic distance alone does not account for the different levels of genotype I RNA replication supported. For the genotype I
HDAg clones, the relative levels of activity in supporting RNA replication were similar for genotype I and genotype III RNA, although the levels of genotype III RNA replication supported were much lower than those supported by any of the genotype III HDAg clones. There was much less variation among the level of RNA replication supported by genotype III HDAg clones from different isolates.
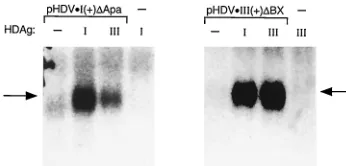
Stabilization of HDV RNA by HDAg.HDAg has been shown to bind specifically to HDV RNA, and this binding is required for HDV RNA replication (19). In transfected cells, the inter-action between HDAg and HDV RNA has also been shown to stabilize HDV RNAs that contain large deletions which pre-serve the ability of the RNA to form the characteristic un-branched rod structure but abolish the replication activity of the RNA (17). To assess whether inefficient interactions be-tween RNAs and HDAg of different genotypes could be re-sponsible for the diminished ability to support replication,
Huh-7 cells were cotransfected with pHDAg-SzI or
pHDAg-SzIII and either pHDVzI(1)DApa or pHDVzIII(1)DBX. The
latter two constructs direct the synthesis of antigenomic RNAs containing internal deletions that preserve the ability of the RNA to form the predicted unbranched rod structure but prevent RNA replication even in the presence of HDAg. Six days posttransfection, RNAs were harvested and analyzed by Northern blotting for levels of antigenomic transcripts
tran-scribed from pHDVzI(1)DApa or pHDVzIII(1)DBX. The
hy-bridization probes did not contain HDAg coding sequences and did not detect appreciable RNA in cells transfected with
either pHDAg-SzI or pHDAg-SzIII alone (Fig. 5).
Both the genotype I and genotype III RNAs accumulated to
higher levels in cells cotransfected with either pHDAg-SzI or
pHDAg-SzIII than in cells cotransfected with a control plasmid
[image:5.612.339.512.69.152.2]that did not encode HDAg (Fig. 5). This accumulation was likely due to stabilization of the RNA, as previously suggested (17), and not due to RNA replication, because genomic RNAs were not detectable (not shown). Genotype III RNA was ap-parently stabilized equally by both genotype I and III HDAg. However, genotype I RNA was stabilized three- to fourfold more effectively by genotype I HDAg than by genotype III HDAg (Fig. 5). This difference was not as great as that be-tween the abilities of these two HDAg species to support RNA replication of genotype I RNA (Fig. 2). These data suggest that the low level of replication of genotype I RNA supported by genotype III antigen might be due, in part, to weak interactions between this RNA and this antigen, but that additional factors are also likely to be important. Conversely, because genotype I HDAg stabilized genotype III RNA about as well as did ge-notype III HDAg, it seems unlikely that weak RNA-HDAg
[image:5.612.53.289.69.221.2]FIG. 4. Genotype-specific support of RNA replication is exhibited by HDAg clones from multiple genotype I and genotype III isolates. Huh-7 cells were transfected with plasmids pHDVzI(1)Ag(2) (top) and pHDVzIII(1)Ag(2) (bottom), which express antigenomic HDV RNA defective for HDAg produc-tion. Cells were also cotransfected with expression constructs for HDAg clones from different isolates (lanes 1 to 4, genotype I; lanes 5 to 9, genotype III) or with the plasmid vector pCMV3 (lanes 10). Cotransfected clones: lanes 1, genotype I prototype clone pHDAg-SzI; lanes 2, isolate U02 (7); lanes 3, isolate I27 (26); lanes 4, isolate I43; lanes 5, genotype III prototype clone pHDAg-SzIII; lanes 6, isolate Peru 5 (10); lanes 7, isolate Peru 13 (10); lanes 8, isolate P20; lanes 9, isolate P21. RNAs were isolated 7 days posttransfection and analyzed by North-ern blotting for HDV genomic RNA. Numbers beneath lanes indicate RNA levels quantified by radioanalytic imaging and normalized for variations in trans-fection efficiency by evaluating expression of a cotransfected reporter gene (14). Values shown are relative to the prototype (lane 1 in the upper panel; lane 5 in the lower panel), which is assigned a value of 100, and are the averages of two transfections.
FIG. 5. Stabilization of HDV RNA by HDAg. Huh-7 cells were transfected with the indicated expression constructs for HDV antigenomic RNA. Cells were also cotransfected with the plasmid vector pCMV3 (lanes 2) or plasmids pHDAg-SzI (lanes I) and pHDAg-SzIII (lanes III), which express genotype I and III HDAg, respectively. RNAs were isolated 6 days posttransfection and ana-lyzed for HDV antigenomic RNA by Northern blotting; the hybridization probes did not include HDAg-encoding sequences transcribed by the HDAg expression constructs.
on November 9, 2019 by guest
http://jvi.asm.org/
interactions can explain the low level of replication of genotype III RNA supported by genotype I HDAg.
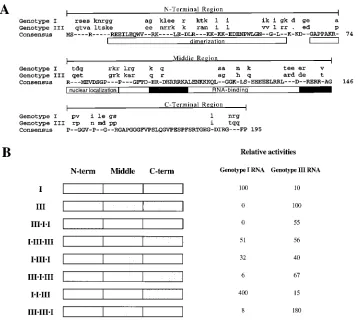
Chimeric HDAg constructs. Comparison of the predicted amino acid sequences of genotype I and III HDAg indicates both conserved and variable regions (Fig. 6A). Studies of ge-notype I HDAg have identified several functional elements, including a dimerization domain (30, 36), nuclear localization signals (36), and a bipartite RNA-binding domain containing two arginine-rich motifs (19). To attempt to identify regions and possible functional elements responsible for the genotype-specific support of HDV RNA replication, chimeric HDAg constructs containing mixtures of type I and type III sequences were created and analyzed for the ability to support replication of type I and type III RNAs. The HDAg coding region was divided into three approximately equal-size regions, which were exchanged between the genotype I and III clones to create a series of six chimeric HDAg expression constructs (Fig. 6B). The amino-terminal third included the dimerization domain, the middle third included the RNA-binding region, and the C-terminal third included a glycine-proline-rich region of unknown function. Amino acid sequence identities were 60, 70, and 78% for the N-terminal, middle, and C-terminal
segments, respectively. Huh-7 cells were cotransfected with
these chimeric HDAg expression constructs and the HDAg(2)
RNA expression construct pHDVzI(1)Ag(2) or pHDVzIII(1)
Ag(2), each of which directs expression of HDAg(2)
antige-nomic RNA. Seven days posttransfection, RNA was harvested and analyzed for the presence of HDV genomic RNA by Northern blot hybridization. RNA levels were quantified by radioanalytic imaging and are presented relative to that ob-served for cotransfection with the prototype HDAg constructs (Fig. 6).
Analysis of replication supported by the chimeric constructs indicated that multiple regions of type I and type III antigens play important roles in supporting replication. The most dra-matic effect was attributed to the N-terminal segment of ge-notype I HDAg, which was required for replication of geno-type I RNA. None of the HDAg constructs containing the genotype III N-terminal segment supported high levels of rep-lication of genotype I RNA; these constructs did, however, support replication of genotype III RNA. Interestingly, the N-terminal region of genotype III HDAg was not as critical for replication of genotype III RNA. Substitution of the genotype
III N-terminal region (i.e., IzIIIzIII) reduced replication of
ge-FIG. 6. Support of RNA replication by chimeric genotype I-genotype III HDAg expression constructs. (A) Comparison of HDAg-S sequences of genotype I and III prototype isolates. Uppercase letters for consensus indicate agreement between genotype I and III clones. Dashes in the consensus indicate positions not in agreement; for each isolate, the identities of these positions are indicated in lowercase. Dots indicate gaps introduced to produce an optimal alignment. Lines above sequences indicate boundaries used in the creation of the chimeric constructs (B). Functional domains identified in other studies are indicated in open boxes beneath sequences. The black boxes within the RNA-binding domain indicate arginine-rich motifs (19). (B) Left, schematic representation of HDAg chimeric constructs. Open boxes indicate genotype I sequences, shaded boxes indicate genotype III sequences. Right, relative levels of RNA replication supported by chimeric HDAg constructs. Huh-7 cells were transfected with pHDVzI(1)Ag(2) (genotype I RNA) or pHDVzIII(1)Ag(2) (genotype III RNA) and with the indicated HDAg expression constructs. RNAs were isolated 7 days posttransfection and analyzed by Northern blotting for the appropriate genomic HDV RNA. RNA levels were quantified by radioanalytic imaging and normalized for expression of a cotransfected reporter gene (14). Values indicated are relative to the levels of expression obtained with the homologous prototype HDAg clone, as described for Fig. 4. Quantitations presented are averages from two independent transfections and did not vary by more than 10% in separate experiments. Relative activities that were less than 1% of the prototypes are shown as 0.
on November 9, 2019 by guest
http://jvi.asm.org/
[image:6.612.123.481.69.389.2]notype III RNA to 56% of the prototype level, compared with no detectable replication of genotype I RNA with the
comple-mentary construct IIIzIzI (Fig. 6). As suggested by the results of
the RNA stabilization experiment, substitution of the genotype I RNA-binding region with that from type III led to a substan-tial decrease in replication of type I RNA (32% of the
proto-type level for construct IzIIIzI [Fig. 6]) but did not eliminate
activity altogether. The complementary substitution within ge-notype III HDAg reduced replication of gege-notype III RNA, but to a lesser extent (67% of the prototype level for construct
IIIzIzIII). Overall, replication of genotype I RNA was more
sensitive to substitutions within genotype I HDAg compared to the sensitivity of genotype III RNA replication to substitutions within genotype III HDAg. Curiously, the C-terminal regions of both HDAg species not only exhibited no genotype speci-ficity but increased replication upon grafting onto the other
HDAg. Compared with the prototypes, construct IzIzIII
sup-ported fourfold-higher replication of genotype I RNA and
construct IIIzIIIzI produced twofold-higher levels of genotype
III RNA.
Effects of HDAg on replication of wild-type HDV.The in-ability of HDAg from genotypes I and III to fully support
replication of HDAg(2) RNA from the other genotype raised
the question of what effects might occur on replication of wild-type HDV. Huh-7 cells were cotransfected with HDV expression constructs for genotype I and genotype III and expression constructs for the short form of HDAg from both genotypes; for comparison, expression constructs for the long form of HDAg were also cotransfected independently. For wild-type genotype I, cotransfection of an expression construct for HDAg-S(I) had little effect on RNA replication, and as expected (15), coexpression of the long form of HDAg sup-pressed replication (Fig. 7). Surprisingly, coexpression of the short form of genotype III HDAg strongly inhibited genotype I RNA replication; remarkably, this inhibition was even greater than that due to the long form of genotype I HDAg (Fig. 7). This inhibition was not limited to the prototype genotype I isolate; RNA replication of another genotype I isolate (clone L [5]; GenBank accession no. L22066) was also strongly inhibited by coexpression of genotype III HDAg (4). The strongest sup-pression of genotype I RNA replication was observed with the long form of genotype III HDAg. Genotype III RNA replica-tion was less affected by the presence of genotype I HDAg.
Cotransfection of pHDAg-SzI with pHDVzIII(1) resulted in a
two- to threefold inhibition of replication, while pHDAg-SzIII
increased replication of the wild-type genotype III RNA.
How-ever, genotype I HDAg-L strongly inhibited replication of ge-notype III RNA, as did gege-notype III HDAg-L.
DISCUSSION
Previous studies of HDV replication have been limited to three different clones of HDV genotype I, two of which are nearly identical. In this report, we find that the roles of the short and long forms of HDAg in viral RNA replication are similar for HDV genotypes I and III, despite sequence varia-tions of about 40%. The short form of HDAg is required for RNA replication (Fig. 3), while the long form inhibits RNA replication (Fig. 7). Moreover, for both genotypes, HDAg can bind HDV RNA of the same type in cells, as evidenced by the stabilization of the RNA in the presence of HDAg (Fig. 5). It is notable that the inhibitory activities of HDAg-L are similar for the two types (Fig. 7), even though the sequence of the C-terminal extension responsible for this inhibitory activity (13) is very different in genotypes I and III (7, 10, 31).
Despite these similarities, however, we describe important functional distinctions between HDV genotypes I and III that further confirm the original classification of sequences within these phylogenetically distinct clades as belonging to unique genotypes (7, 26, 31). The observation that HDAg clones from genotype I and III isolates cannot fully support replication of HDV RNA from the other genotype (Fig. 2, 4, and 6) strongly suggests that the support of replication by HDAg is genotype specific. Although the wide variation of genotype I RNA rep-lication supported by genotype I HDAg clones raises the for-mal possibility that there are also isolate-specific effects (within a genotype), it seems unlikely that the genotype-specific effects observed are simply an extension of an isolate-specific effect which is related to genetic similarities. There was no correla-tion between sequence divergence and the amount of replica-tion supported by the different genotype I HDAg clones. The relative levels of genotype I RNA replication supported by HDAg clones from isolates U02, I27, and I43 were 1.0, 0.2, and 11, respectively (Fig. 4), while the sequence similarities be-tween these isolates and the prototype were 6.5, 7.3, and 9.6%, respectively (4, 26, 31). Moreover, notwithstanding possible isolate-specific effects, the genotype-specific effect was clear: all HDAg clones supported replication of the prototype RNA from the same genotype substantially better than the prototype RNA of the other genotype, regardless of their activities on the RNA of the same genotype (Fig. 4).
The genotype-specific activity of HDAg-S from genotypes I and III in supporting HDV RNA replication (Fig. 3, 4, and 6) suggests that these clades may best be referred to as types rather than genotypes (i.e., HDV-I and HDV-III), the distinc-tion being that the descripdistinc-tion of the uniqueness of the differ-ent sequence groups is not limited to genetic analysis but includes functional activity essential to the virus life cycle. In this regard, it is notable that the capsid and polymerase pro-teins of hepatitis B virus and woodchuck hepatitis virus, which infect different hosts, have been shown to be functionally in-terchangeable (37). The different electrophoretic mobilities of genotype I and III HDAg, which could be due to differences in posttranslational modifications (e.g., phosphorylation) or structural variations, further confirm the differences between these genotypes and may be related to the different functional activities of the two species. Because the regions of sequence variation between HDAg of HDV-I and HDV-III also vary in genotype II (7), it seems likely that the genotype-specific sup-port of replication will also apply to HDV genotype II. Whether such is the case can readily be determined after full-length clones of HDV genotype II are available.
FIG. 7. Inhibition of genotype I RNA replication by genotype III HDAg. Huh-7 cells were transfected with pCMV3-DC131.2 (left) or pHDVzIII(1) (right), which express wild-type 1.2 unit-length genotype I and genotype III antigenomic RNA, respectively. Cells were also cotransfected with the indicated HDAg expression constructs or with the plasmid vector pCMV3 (lanes None). RNAs were isolated 6 days posttransfection and analyzed by Northern blotting for HDV antigenomic RNA.
on November 9, 2019 by guest
http://jvi.asm.org/
pair than between the homologous RNA and HDAg species. Thus, the inability of type III HDAg to support type I RNA replication could be due, in part, to a weak interaction between this HDAg-RNA pair. However, this explanation does not appear to apply to the inability of type I HDAg to support type III RNA replication, because type I HDAg apparently binds to and stabilizes type III RNA (Fig. 5). Perhaps the specific mech-anisms by which the short form of HDAg facilitates RNA replication are not the same for genotypes I and III.
The nonreciprocal nature of activities of type I and III HDAg in stabilizing type I and type III RNAs is curious. This stabilization is most likely due to binding between HDAg and HDV RNA, as previously reported (17). Both type I and type III RNAs used in the experiments presented in Fig. 5 are predicted to form unbranched rod structures with extensive base pairing, and most of the sequences within the arginine-rich motifs are highly conserved between types I and III (Fig. 6). At least for genotype I, these features have been shown to be required for binding between HDAg and HDV RNA (12, 19). However, the specific determinants of binding have not been identified in either the RNA or HDAg. Further compar-ative analysis of the binding activities of the RNAs and HDAgs of types I and III is likely to prove instructive for understanding these determinants (on both HDAg and HDV RNA) and their roles in HDV RNA replication.
The broad range (about 20-fold) of replication supported by HDAg from different genotype I isolates was remarkably greater than that observed among genotype III isolates, par-ticularly as the extents of sequence variation in a segment including the C-terminal half of the antigen coding region were similar among the two groups of isolates. As discussed above, it is possible that variations in the level of HDV genotype I prototype RNA replication supported by the different geno-type I HDAg clones were due to isolate-specific interactions between HDAg and HDV RNA. Alternatively, the wide range of replication supported by the different type I isolates could be due to differences in the inherent abilities of HDAg from these isolates to support replication. Indeed, viremia varies considerably among different patients (26), and it is tempting to speculate that the broad spectrum of disease associated with type I HDV infection (26) is due, in part, to variations in the ability of HDAg in different isolates to support high levels of RNA replication. Analyses of the abilities of HDAg clones to support replication of HDV RNAs from additional isolates, particularly those with divergent sequences and/or disease and viremia patterns, will be useful in examining these possibilities. The strong inhibition of genotype I RNA replication by genotype III HDAg could have implications for our under-standing of both the molecular biology and the epidemiology of this agent, as well as for the development of novel thera-peutic approaches. Because genotype III HDAg-S bound ge-notype I RNA only weakly (Fig. 5), it seems unlikely that the mechanism of inhibition would involve sequestration of the genotype I RNA by genotype III HDAg. An alternative pos-sibility involves direct interaction between HDAg-S of geno-types I and III such that the replication activity of genotype I HDAg is inhibited. In this event, the lack of inhibition of genotype III RNA replication by genotype I HDAg could be
tivity could be used to develop therapies to inhibit genotype I HDV replication in infected patients.
ACKNOWLEDGMENTS
We are grateful to M. Sjo¨gren and A. Colichon for providing HDV isolates from Peru. We thank Thomas Brown for excellent technical assistance.
This work was supported by NIAID contract NO1-AI-45179.
REFERENCES
1. Beard, M. R., T. B. MacNaughton, and E. J. Gowans. 1996. Identification and characterization of a hepatitis delta virus RNA transcriptional promoter. J. Virol. 70:4986–4995.
2. Bonino, F., B. Hoyer, E. Ford, J. W. Shih, R. H. Purcell, and J. L. Gerin. 1981. The delta agent: HBsAg particles with delta antigen and RNA in the serum of an HBV carrier. Hepatology 1:127–131.
3. Casey, J. L. 1996. Hepatitis delta virus: genetics and pathogenesis. Clin. Lab. Med. 16:451–464.
4. Casey, J. L. 1997. Unpublished results.
5. Casey, J. L., K. F. Bergmann, T. L. Brown, and J. L. Gerin. 1993. Determi-nants of RNA editing in hepatitis delta virus. Prog. Clin. Biol. Res. 382:5–11. 6. Casey, J. L., K. F. Bergmann, T. L. Brown, and J. L. Gerin. 1992. Structural requirements for RNA editing in hepatitis delta virus: evidence for a uridine-to-cytidine editing mechanism. Proc. Natl. Acad. Sci. USA 89:7149–7153. 7. Casey, J. L., T. L. Brown, E. J. Colan, F. S. Wignall, and J. L. Gerin. 1993.
A genotype of hepatitis D virus that occurs in northern South America. Proc. Natl. Acad. Sci. USA 90:9016–9020.
8. Casey, J. L., and J. L. Gerin. 1995. Hepatitis D virus RNA editing: specific modification of adenosine in the antigenomic RNA. J. Virol. 69:7593–7600. 9. Casey, J. L., and J. L. Gerin. 1995. Hepatitis delta virus: RNA editing and genotype variations, p. 111–124. In G. Dinter-Gottlieb (ed.), The unique hepatitis delta virus. R. G. Landes Company, Austin, Tex.
10. Casey, J. L., G. A. Niro, R. E. Engle, A. Vega, H. Gomez, M. McCarthy, D. M.
Watts, K. C. Hyams, and J. L. Gerin.1996. Hepatitis B virus (HBV)/hepatitis
D virus (HDV) coinfection in outbreaks of acute hepatitis in the Peruvian Amazon Basin: the roles of HDV genotype III and HBV genotype F. J. Infect. Dis. 174:920–926.
11. Chang, M. F., C. Y. Sun, C. J. Chen, and S. C. Chang. 1993. Functional motifs of delta antigen essential for RNA binding and replication of hepatitis delta virus. J. Virol. 67:2529–2536.
12. Chao, M., S. Y. Hsieh, and J. Taylor. 1991. The antigen of hepatitis delta virus: examination of in vitro RNA-binding specificity. J. Virol. 65:4057– 4062.
13. Chao, M., S. Y. Hsieh, and J. Taylor. 1990. Role of two forms of hepatitis delta virus antigen: evidence for a mechanism of self-limiting genome rep-lication. J. Virol. 64:5066–5069.
14. Cullen, B. R., and M. H. Malim. 1992. Secreted placental alkaline phospha-tase as a eukaryotic reporter gene. Methods Enzymol. 216:362–368. 15. Kuo, M. Y., M. Chao, and J. Taylor. 1989. Initiation of replication of the
human hepatitis delta virus genome from cloned DNA: role of delta antigen. J. Virol. 63:1945–1950.
16. Kuo, M. Y., L. Sharmeen, G. G. Dinter, and J. Taylor. 1988. Characterization of self-cleaving RNA sequences on the genome and antigenome of human hepatitis delta virus. J. Virol. 62:4439–4444.
17. Lazinski, D. W., and J. M. Taylor. 1994. Expression of hepatitis delta virus RNA deletions: cis and trans requirements for self-cleavage, ligation, and RNA packaging. J. Virol. 68:2879–2888.
18. Lazinski, D. W., and J. M. Taylor. 1993. Relating structure to function in the hepatitis delta virus antigen. J. Virol. 67:2672–2680.
19. Lee, C. Z., J. H. Lin, M. Chao, K. McKnight, and M. M. Lai. 1993. RNA-binding activity of hepatitis delta antigen involves two arginine-rich motifs and is required for hepatitis delta virus RNA replication. J. Virol. 67:2221– 2227.
20. Lin, J. H., M. F. Chang, S. C. Baker, S. Govindarajan, and M. M. Lai. 1990. Characterization of hepatitis delta antigen: specific binding to hepatitis delta virus RNA. J. Virol. 64:4051–4058.
21. Luo, G. X., M. Chao, S. Y. Hsieh, C. Sureau, K. Nishikura, and J. Taylor. 1990. A specific base transition occurs on replicating hepatitis delta virus RNA. J. Virol. 64:1021–1027.
22. MacNaughton, T. B., E. J. Gowans, A. R. Jilbert, and C. J. Burrell. 1990.
on November 9, 2019 by guest
http://jvi.asm.org/
Hepatitis delta virus RNA, protein synthesis and associated cytotoxicity in a stably transfected cell line. Virology 177:692–698.
23. MacNaughton, T. B., Y. J. Wang, and M. M. Lai. 1993. Replication of hepatitis delta virus RNA: effect of mutations of the autocatalytic cleavage sites. J. Virol. 67:2228–2234.
24. Makino, S., M. F. Chang, C. K. Shieh, T. Kamahora, D. M. Vannier, S.
Govindarajan, and M. M. Lai.1987. Molecular cloning and sequencing of a
human hepatitis delta (delta) virus RNA. Nature 329:343–346.
25. Maynard, J. E., S. C. Hadler, and H. A. Fields. 1987. Delta hepatitis in the Americas: an overview. Prog. Clin. Biol. Res. 234:493–505.
26. Niro, G. A., A. Smedile, A. Andriulli, M. Rizzetto, J. L. Gerin, and J. L.
Casey.1997. Predominance of hepatitis delta virus genotype I among
chron-ically infected Italian patients. Hepatology 25:728–734.
27. Pohl, C., B. M. Baroudy, K. F. Bergmann, P. J. Cote, R. H. Purcell, J.
Hoofnagle, and J. L. Gerin.1987. A human monoclonal antibody that
rec-ognizes viral polypeptides and in vitro translation products of the genome of the hepatitis D virus. J. Infect. Dis. 156:622–629.
28. Polson, A. G., H. L. Ley III, B. L. Bass, and J. L. Casey. Hepatitis delta virus RNA editing is highly specific for the amber/W site, and is suppressed by hepatitis delta antigen. Mol. Cell. Biol., in press.
29. Rizzetto, M. 1983. The delta agent. Hepatology 3:729–737.
30. Rozzelle, J. J., J. G. Wang, D. S. Wagner, B. W. Erickson, and S. M. Lemon. 1995. Self-association of a synthetic peptide from the N terminus of the hepatitis delta virus protein into an immunoreactive alpha-helical multimer.
Proc. Natl. Acad. Sci. USA 92:382–386.
31. Shakil, A. O., S. Hadziyannis, J. H. Hoofnagle, A. M. DiBisceglie, J. L. Gerin,
and J. L. Casey.1997. Geographic distribution and genetic variability of
hepatitis delta virus genotype I. Virology 234:160–167.
32. Wang, K. S., Q. L. Choo, A. J. Weiner, J. H. Ou, R. C. Najarian, R. M.
Thayer, G. T. Mullenbach, K. J. Denniston, J. L. Gerin, and M. Houghton.
1986. Structure, sequence and expression of the hepatitis delta viral genome. Nature 323:508–514.
33. Wu, J. C., C. M. Chen, I. J. Sheen, S. D. Lee, H. M. Tzeng, and K. B. Choo. 1995. Evidence of transmission of hepatitis D virus to spouses from sequence analysis of the viral genome. Hepatology 22:1656–1660.
34. Wu, J. C., K. B. Choo, C. M. Chen, T. Z. Chen, T. I. Huo, and S. D. Lee. 1995. Genotyping of hepatitis D virus by restriction-fragment length polymorphism and relation to outcome of hepatitis D. Lancet 346:939–941.
35. Wu, T. T., H. J. Netter, D. W. Lazinski, and J. M. Taylor. 1997. Effects of nucleotide changes on the ability of hepatitis delta virus to transcribe, pro-cess, and accumulate unit-length, circular RNA. J. Virol. 71:5408–5414. 36. Xia, Y. P., C. T. Yeh, J. H. Ou, and M. M. Lai. 1992. Characterization of
nuclear targeting signal of hepatitis delta antigen: nuclear transport as a protein complex. J. Virol. 66:914–921.
37. Zierman, R., and D. Ganem. 1996. Homologous and heterologous comple-mentation of HBV and WHV capsid and polymerase functions in RNA encapsidation. Virology 219:350–356.