0022-538X/95/$04.0010
Copyrightq1995, American Society for Microbiology
The Highly Conserved Aspartic Acid Residue between
Hypervariable Regions 1 and 2 of Human Immunodeficiency
Virus Type 1 gp120 Is Important for Early Stages of
Virus Replication
WEI-KUNG WANG, MAX ESSEX,ANDTUN-HOU LEE*
Department of Cancer Biology, Harvard School of Public Health, Boston, Massachusetts 02115
Received 5 August 1994/Accepted 28 September 1994
Between hypervariable regions V1 and V2 of human immunodeficiency virus type 1 (HIV-1) gp120 lies a cluster of relatively conserved residues. The contribution of nine charged residues in this region to virus infectivity was evaluated by single-amino-acid substitutions in an infectious provirus clone. Three of the HIV-1 mutants studied had slower growth kinetics than the wild-type virus. The delay was most pronounced in a mutant with an alanine substituted for an aspartic acid residue at position 180. This aspartic acid is conserved by all HIV-1 isolates with known nucleotide sequences. Substitutions with three other residues at this position, including a negatively charged glutamic acid, all affected virus infectivity. The defect identified in these mutants suggests that this aspartic acid residue is involved in the early stages of HIV-1 replication.
Human immunodeficiency virus type 1 (HIV-1) is the etio-logic agent of AIDS (2, 8, 28, 30). HIV-1 isolates display prominent genomic heterogeneity, particularly in the region that encodes for envelope glycoprotein gp120 (3, 9, 25, 29, 31, 36). Variation in gp120 is clustered in five discrete hypervari-able regions, V1 to V5, which are interspersed between five conserved regions, C1 to C5 (23, 25). Despite the sequence divergence, there are 18 highly conserved cysteine residues in the mature gp120 (25). Leonard et al. have determined the locations of nine disulfide bonds in a prototypic gp120 and have proposed a secondary structure that groups gp120 resi-dues into five major cysteine loops (19).
The second major cysteine loop has a complex double-loop structure with two disulfide bonds involving cysteine residue pairs 119 and 205 and 126 and 196, near the base of the loop, and a third disulfide bond involving residues 131 and 157 (Fig. 1). Between residues 119 and 205, there are about 90 amino acid residues, which account for approximately one-fifth of the gp120 coding sequence. Functionally, residues within the sec-ond major cysteine loop are reported to interact with those in V3 and other regions to influence syncytium formation (1). However, several observations suggest that the second major cysteine loop of gp120 is structurally independent from the rest of the molecule. First, the second major cysteine loop can be removed without affecting expression and processing of gp120 or the interaction between gp120 and the transmembrane subunit gp41 (37). Second, the entire second major cysteine loop is dispensable for CD4 binding. This suggests that this loop is not tightly linked to the C1, C2, C3, and C4 regions, which account for approximately three-fifths of the gp120 sequence (4, 15, 17, 20, 26, 27, 33). Third, deletion of the second major cysteine loop does not abolish binding of several broadly neutralizing human monoclonal gp120 antibodies that are effective at blocking gp120-CD4 interactions to gp120 (24, 37). Residues that are involved in the binding of these mono-clonal antibodies have also been mapped to C1, C2, C3, and C4
(34, 35), again suggesting that this loop is not tightly linked to the rest of gp120. Last, the second major cysteine loop alone is sufficient to present some conformational epitopes of gp120 when fused to the surface glycoprotein of murine leukemia virus (12).
Hypervariable regions V1 and V2 are located within the second major cysteine loop (19, 25). These two hypervariable regions contain residues that are highly divergent among various HIV-1 isolates. Insertions and deletions, which are often linked to the creation or removal of potential N-linked glycosylation sites, are frequently found in V1 and V2 (25). The highly divergent V1 and V2 contrast sharply with a cluster of relatively conserved residues located between V1 and V2 and flanked by a glycine at position 152 and an aspartic acid at position 185 (Fig. 1 and Table 1). Within this relatively conserved region there are nine charged residues, a number disproportionately higher than that found in most other re-gions of gp120 (25). In this study, the relative importance of these charged residues in HIV-1 infectivity was investigated.
Using site-directed mutagenesis, nine charged amino acid residues in the region flanked by V1 and V2 were each replaced with an alanine in a previously described infectious provirus clone, HXB2 (6, 16, 18). Corresponding to the position of each alanine substitution, mutant proviruses were designated 153EA, 155KA, 166RA, 168KA, 171KA/172EA, 178KA, 180DA, and 185DA. To study the infectivity of these
mutants, 4mg of mutant provirus DNA or wild-type DNA was
transfected into 3 3 106 to 5 3 106 COS-7 cells, using a
previously described DEAE-dextran method (14, 38). Cell-free virions from culture supernatants of COS-7 transfectants were collected 72 h posttransfection. Equal amounts of mutant and wild-type viruses, as measured by reverse transcriptase (RT)
activity (39), were used to infect CD41 SupT1 cells.
Virus-infected cultures were monitored every 4 days for RT activity. As shown in Fig. 2, the growth kinetics of these mutants followed three patterns. Mutant viruses 166RA, 168KA, 171KA/172EA, 178KA, and 185DA had growth kinetics that were comparable to or slightly faster than those of the wild-type virus (Fig. 2A). The growth kinetics of mutant viruses 153EA and 155KA lagged slightly behind those of the
wild-* Corresponding author. Mailing address: Department of Cancer Biology, Harvard School of Public Health, 665 Huntington Ave., Boston, MA 02115. Phone: (617) 432-1302. Fax: (617) 739-8348.
538
on November 9, 2019 by guest
http://jvi.asm.org/
type virus (Fig. 2B). The substitution introduced to the aspartic acid residue at position 180 had the most significant effect on growth kinetics (Fig. 2C). These results demonstrate that alanine substitutions for most of the nine conserved charged residues were well tolerated and that these residues are unlikely to have critical roles in virus infectivity.
Among the nine charged residues studied, the aspartic acid at position 180 was the most invariable among all HIV-1 isolates of different genetic clades (25) (Table 1). Amino acid substitutions with a positively charged lysine residue, nated 180DK, and a noncharged asparagine residue, desig-nated 180DN, were introduced to the aspartic acid at position 180 to evaluate the requirement of a negatively charged residue in this position. As shown in Fig. 3A, the growth kinetics of mutant virus 180DN were similar to those of mutant 180DA and were significantly slower than those of the wild-type virus. In the culture infected by mutant 180DK, RT activity was not detected during the entire follow-up period, indicating that the positively charged lysine residue at this position was not tolerated. To further test whether a negatively charged residue other than aspartic acid could confer the
wild-type phenotype, substitution with a glutamic acid residue, 180DE, was also carried out. The growth kinetics of mutant 180DE were similar to those for 180DA and 180DN (Fig. 3A), suggesting that the aspartic acid residue at this position and not the charge itself is critical for virus infectivity.
To investigate whether a defect in expression or processing of the envelope protein could account for the delay in growth kinetics for mutant viruses with substitutions introduced to position 180, lysates from COS-7 transfected cells were
[image:2.612.316.555.70.561.2]pre-FIG. 1. Schematic drawing of the second major cysteine loop of gp120. The loop is formed by two disulfide bonds involving cysteine pairs 119 and 205 and 126 and 196, near the base of the loop, and a third disulfide bond involving cysteine residues 131 and 157. Between hypervariable regions V1 and V2 is a cluster of relatively conserved residues flanked by a glycine at position 152 and an aspartic acid at position 185. The nine charged residues located within this relatively conserved region are indicated by dots.
FIG. 2. Growth kinetics of mutant and wild-type (WT) viruses in CD41 SupT1 cells. RT activity in culture supernatants was monitored for wild-type virus and mutant viruses 166RA, 168KA, 171KA/172EA, 178KA, and 185DA (A), wild-type virus and mutant viruses 153EA and 155KA (B), and wild-type virus and mutant virus 180DA (C). Mock, cultures infected by cell-free super-natants of mock-transfected cultures.
TABLE 1. Degree of conservation of nine charged residues in the region flanked by V1 and V2 of HIV-1 gp120
Residue
No. of isolatesa
from the indicated clade:
A (n54)
B (n537)
C (n55)
D (n55)
E (n51)
O (n53)
E-153 4 37 5 2 1 2
K-155 4 32 4 5 0 2
R-166 4 31 4 2 0 1
K-168 4 34 2 5 1 3
K-171 4 33 5 0 1 1
E-172 0 28 0 1 0 0
K-178 1 29 4 1 1 0
D-180 4 37 5 5 1 3
D-185 2 30 3 5 0 0
a
Number of isolates having the identical residues, at a given position, as in the consensus B sequence. n, number of isolates in each clade as listed in reference 25.
on November 9, 2019 by guest
http://jvi.asm.org/
[image:2.612.104.250.72.247.2] [image:2.612.58.297.584.709.2]pared 72 h posttransfection for Western immunoblot analysis (18, 39). As shown in Fig. 3B, both gp160 and gp120 were readily detected by a reference sheep anti-gp120 serum, indi-cating that expression and processing of the envelope protein were not drastically affected by the substitutions. Western blot analysis was also conducted on lysates of virus pellets derived from cell-free supernatants of COS-7 transfectants (18, 39). As
shown in Fig. 3C, the amount of gp120 relative to p24 in the lysates of mutant viruses was not significantly different from that of the wild-type virus. This finding indicates that the delay in growth kinetics of the mutant viruses could not be attributed to defects in incorporation of gp120 into virions or association between gp120 and gp41.
An alternative possibility is that the delay in growth kinetics of the mutant viruses is the result of a defect in early stages of virus replication. To address this possibility, a previously described complementation assay was adapted (10). The abil-ities of mutant viruses to complement an env-defective and
rev-truncated provirus, dBgldrev (39), which is a modification
of pHXBDenvCAT (10), were measured. In this one-round
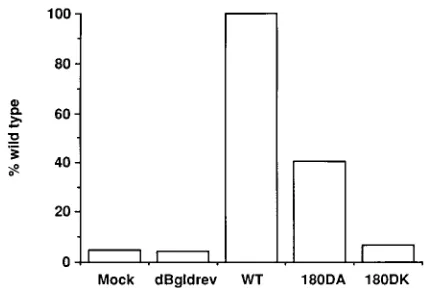
replication assay, target cells infected by mutant viruses with a defect in the early stages of virus replication are expected to have lower levels of chloramphenicol acetyltransferase (CAT) activity than those infected by wild-type virus. The envelope gene of mutants 180DA and 180DK and that of the wild-type virus were constructed in an envelope expression vector pSVIII (10). Mutant viruses 180DN and 180DE had growth kinetics similar to those of mutant virus 180DA, and their envelope genes were not included in this analysis. Each pSVIII-derived envelope expression plasmid was cotransfected with dBgldrev in COS-7 cells. Cell-free virions with mutant or wild-type envelope proteins were harvested from COS-7 supernatants 72 h posttransfection, normalized for RT activity, and used to infect SupT1 cells. CAT activity in the SupT1 cell lysates was analyzed 48 h postinfection. As shown in Fig. 4, both mutants exhibited a reduction in CAT gene expression relative to that of the wild type, indicating that they had a defect in the early stages of virus replication. Consistent with the growth kinetics shown in Fig. 3A, CAT activity for mutant 180DK was significantly less than that for mutant 180DA.
Four of the nine charged residues studied here were previ-ously analyzed (32). The arginine residue at position 166 was replaced by a leucine residue and designated 166 R/L. The glutamic acid residue at position 153 and the glycine residue at position 152 were simultaneously substituted with a methio-nine residue and a serine residue, respectively, and designated 152/153 GE/SM. The lysine residue at position 168 was substi-tuted with a leucine residue and designated 168 K/L. The aspartic acid residue at position 180 and the leucine residue at position 179 were simultaneously substituted with a leucine
[image:3.612.327.541.70.214.2]FIG. 3. (A) Growth kinetics of wild-type virus and 180 mutant viruses. RT activity in culture supernatants of CD41SupT1 cells was monitored for mutants 180DA, 180DE, 180DN, and 180DK. (B) Western blot analysis of envelope proteins expressed in COS-7 cells transfected with proviral DNA of wild-type (WT) or 180 mutant viruses, using a reference sheep anti-gp120 serum. gp160 and gp120 are indicated. (C) Western blot analysis of envelope proteins incorporated into virions of wild-type and 180 mutant viruses. Viral lysates prepared from wild-type, 180DA, 180DE, 180DN, and 180DK viruses were reacted with pooled HIV-1-positive sera. The specificity of gp120 was also verified by a reference sheep anti-gp120 serum (data not shown). Lane MW, molecular weight markers.
FIG. 4. CAT activity of virions with mutant envelope proteins. Cell-free virions derived from COS-7 cells cotransfected with a defective provirus clone, dBgldrev, and an envelope expression vector were used to infect SupT1 cells. CAT activity in SupT1 cells infected by viruses containing wild-type (WT), 180DA, or 180DK envelope proteins was expressed as the percent conversion of chloramphenicol relative to that of the wild type.
on November 9, 2019 by guest
http://jvi.asm.org/
to that of the wild-type envelope but had a significant defect in syncytium induction. It is unclear whether the phenotypes observed for mutants 152/153 GE/SM and 179/180 LD/DL were caused by substitutions to either or both residues. How-ever, our findings indicate that the glutamic acid at position 153 did not have a drastic effect on gp120 incorporation into the virus (data not shown). Substituting three other amino acid residues for the aspartic acid residue at position 180, including a negatively charged glutamic acid, was not well tolerated by the virus. The importance of this aspartic acid residue in virus infectivity is consistent with the high degree of conservation of this residue by all HIV-1 isolates across genetic clades (25).
The initial interaction between HIV-1 and most target cells is believed to involve high-affinity binding between gp120 and CD4, the primary receptor of HIV (5, 13, 21, 22). This interaction is followed by the so-called post-CD4-binding events, which include a series of incompletely characterized processes, possibly involving different regions of gp120, gp41, and undefined cellular proteins. The charged residues studied here are located within a region that is not known to be involved in CD4 binding. It is likely that the aspartic acid residue at position 180 is critical for post-CD4-binding steps. Mechanistically, it is conceivable that this aspartic residue may interact with other residues within the second major cysteine loop or residues in other regions of gp120 or gp41 to confer the conformation required for virus entry. This interpretation was not supported by our finding that expression of mutant enve-lope proteins and their incorporation into virions were not significantly altered. In addition, the recognition of 180 mutant proteins by 1.5e, a previously described conformation-depen-dent human gp120 monoclonal antibody, was not affected (data not shown). However, indirect involvement of the aspar-tic acid residue at position 180 in the early steps of virus entry cannot be completely ruled out. Alternatively, this aspartic acid residue plays a direct role during the process of virus entry. This interpretation is compatible with previous reports that several neutralizing monoclonal antibodies appear to target residues overlapping with this aspartic acid residue (7, 11, 32). Taken together, this aspartic acid residue should be considered as a potential target for future antiviral design.
We thank C. N. Lee, X. F. Yu, and Z. Matsuda for helpful discussions; R. C. Gallo for HXB2; J. G. Sodroski for kindly providing pSVIII and pHXBDenvCAT, from which pBgldrev was derived; and E. Conway for editorial assistance. Sheep anti-gp120 serum was obtained from the AIDS Research and Reference Reagent Program, National Institutes of Health, Bethesda, Md.
This work was supported in part by Public Health Service grants CA-39805 and HL-33774 from the National Institutes of Health and 17-90-C-0151 from the U.S. Army. W.-K. Wang is supported by training grant 5 D43 TW00004 from the Fogarty International Center, National Institutes of Health.
and R. A. Weiss.1984. The CD4 (T4) antigen is an essential component of the receptor for the AIDS retrovirus. Nature (London) 312:763–767.
6. Fisher, A. G., E. Collalti, L. Ratner, R. C. Gallo, and F. Wong-Staal. 1985. A molecular clone of HTLV-III with biological activity. Nature (London) 316:262–265.
7. Fung, M. S., C. R. Sun, W. L. Gordon, R. S. Liou, T. W. Chang, W. N. Sun, E. S. Daar, and D. D. Ho.1992. Identification and characterization of a neutralization site within the second variable region of human immunode-ficiency virus type 1 gp120. J. Virol. 66:848–856.
8. Gallo, R. C., S. Z. Salahuddin, M. Popovic, G. M. Shearer, M. Kaplan, B. F. Haynes, T. J. Palker, R. Redfield, J. Oleske, B. Safai, G. White, P. Foster, and P. D. Markham.1984. Frequent detection and isolation of cytopathic retroviruses (HTLV-III) from patients with AIDS and at risk for AIDS. Science 224:500–503.
9. Hahn, B. H., M. A. Gonda, G. M. Shaw, M. Popovic, J. A. Hoxie, R. C. Gallo, and F. Wong-Staal. 1985. Genomic diversity of the acquired immune deficiency syndrome virus HTLV-III: different viruses exhibit greatest diver-gence in their envelope genes. Proc. Natl. Acad. Sci. USA 82:4813–4817. 10. Helseth, E., M. Kowalski, D. Gabuzda, U. Olshevsky, W. Haseltine, and J.
Sodroski.1990. Rapid complementation assays measuring replicative poten-tial of human immunodeficiency virus type 1 envelope glycoprotein mutants. J. Virol. 64:2416–2420.
11. Ho, D. D., M. S. Fung, Y. Z. Cao, X. L. Li, C. Sun, T. W. Chang, and N. C. Sun.1991. Another discontinuous epitope on glycoprotein gp120 that is important in human immunodeficiency virus type 1 neutralization is identi-fied by a monoclonal antibody. Proc. Natl. Acad. Sci. USA 88:8949–8952. 12. Kayman, S. C., Z. Wu, K. Revesz, H. Chen, R. Kopelman, and A. Pinter.
1994. Presentation of native epitopes in the V1/V2 and V3 regions of human immunodeficiency virus type 1 gp120 by fusion glycoproteins containing isolated gp120 domains. J. Virol. 68:400–410.
13. Klatzmann, D., E. Champagne, S. Chamaret, J. Gruest, D. Guetard, T. Hercend, and L. Montagnier.1984. T-lymphocyte T4 molecule behaves as the receptor for human retrovirus LAV. Nature (London) 312:767–768. 14. Kornfeld, H., N. Riedel, G. A. Viglianti, V. Hirsch, and J. I. Mullins. 1987.
Cloning of HTLV-4 and its relation to simian and human immunodeficiency viruses. Nature (London) 326:610–613.
15. Kowalski, M., J. Potz, L. Basiripour, T. Dorfman, W. C. Goh, E. Terwilliger, A. Dayton, C. Rosen, W. Haseltine, and J. Sodroski.1987. Functional regions of the envelope glycoprotein of human immunodeficiency virus type 1. Science 237:1351–1355.
16. Kunkel, T. A. 1985. Rapid and efficient site-specific mutagenesis without phenotypic selection. Proc. Natl. Acad. Sci. USA 82:488–492.
17. Lasky, L. A., G. Nakamura, D. H. Smith, C. Fennie, C. Shimasaki, E. Patzer, P. Berman, T. Gregory, and D. J. Capon.1987. Delineation of a region of the human immunodeficiency virus type 1 gp120 glycoprotein critical for inter-action with the CD4 receptor. Cell 50:975–985.
18. Lee, W.-R., X.-F. Yu, W.-J. Syu, M. Essex, and T.-H. Lee. 1992. Mutational analysis of conserved N-linked glycosylation sites of human immunodefi-ciency virus type 1 gp41. J. Virol. 66:1799–1803.
19. Leonard, C. K., M. W. Spellman, L. Riddle, R. J. Harris, J. N. Thomas, and T. J. Gregory.1990. Assignment of intrachain disulfide bonds and charac-terization of potential glycosylation sites of the type 1 recombinant human immunodeficiency virus envelope glycoprotein (gp120) expressed in Chinese hamster ovary cells. J. Biol. Chem. 265:10373–10382.
20. Linsley, P. S., J. A. Ledbetter, E. Kinney-Thomas, and S.-L. Hu. 1988. Effects of anti-gp120 monoclonal antibodies on CD4 receptor binding by the env protein of human immunodeficiency virus type 1. J. Virol. 62:3695–3702. 21. McDougal, J. S., M. S. Kennedy, J. M. Sligh, S. P. Cort, A. Mawle, and J. K.
Nicholson.1986. Binding of HTLV-III/LAV to T41T cells by a complex of the 110K viral protein and the T4 molecule. Science 231:382–385. 22. McDougal, J. S., J. K. Nicholson, G. D. Cross, S. P. Cort, M. S. Kennedy, and
A. C. Mawle.1986. Binding of the human retrovirus HTLV-III/LAV/ARV/ HIV to the CD4 (T4) molecule: conformation dependence, epitope map-ping, antibody inhibition, and potential for idiotypic mimicry. J. Immunol. 137:2937–2944.
on November 9, 2019 by guest
http://jvi.asm.org/
23. Modrow, S., B. E. Hahn, G. M. Shaw, R. C. Gallo, F. Wong-Staal, and H. Wolf.1987. Computer-assisted analysis of envelope protein sequences of seven human immunodeficiency virus isolates: prediction of antigenic epitopes in conserved and variable regions. J. Virol. 61:570–578. 24. Moore, J. P., Q. J. Sattentau, H. Yoshiyama, M. Thali, M. Charles, N.
Sullivan, S.-W. Poon, M. S. Fung, F. Traincard, M. Pinkus, G. Robey, J. E. Robinson, D. D. Ho, and J. Sodroski.1993. Probing the structure of the V2 domain of human immunodeficiency virus type 1 surface glycoprotein gp120 with a panel of eight monoclonal antibodies: human immune response to the V1 and V2 domains. J. Virol. 67:6136–6151.
25. Myers, G., B. Korber, S. Wain-Hobson, R. F. Smith, and G. N. Pavlakis. 1993. Human retroviruses and AIDS 1993: a compilation and analysis of nucleic acid and amino acid sequences. Theoretical Biology and Biophysics, Los Alamos National Laboratory, Los Alamos, N.M.
26. Olshevsky, U., E. Helseth, C. Furman, J. Li, W. Haseltine, and J. Sodroski. 1990. Identification of individual human immunodeficiency virus type 1 gp120 amino acids important for CD4 receptor binding. J. Virol. 64:5701– 5707.
27. Pollard, S. R., M. D. Rosa, J. J. Rosa, and D. C. Wiley. 1992. Truncated variants of gp120 bind CD4 with high affinity and suggest a minimum CD4 binding region. EMBO J. 11:585–591.
28. Popovic, M., M. G. Sarngadharan, E. Read, and R. C. Gallo. 1984. Detection, isolation, and continuous production of cytopathic retroviruses (HTLV-III) from patients with AIDS and pre-AIDS. Science 224:497–500. 29. Ratner, L., R. C. Gallo, and F. Wong-Staal. 1985. HTLV-III, LAV, ARV are
variants of same AIDS virus. Nature (London) 313:636–637. (Letter.) 30. Sarngadharan, M. G., M. Popovic, L. Bruch, J. Schupbach, and R. C. Gallo.
1984. Antibodies reactive with human T-lymphotropic retroviruses (HTLV-III) in the serum of patients with AIDS. Science 224:506–508.
31. Shaw, G. M., B. H. Hahn, S. K. Arya, J. E. Groopman, R. C. Gallo, and F. Wong-Staal. 1984. Molecular characterization of human T-cell leukemia
(lymphotropic) virus type III in the acquired immune deficiency syndrome. Science 226:1165–1171.
32. Sullivan, N., M. Thali, C. Furman, D. D. Ho, and J. Sodroski. 1993. Effect of amino acid changes in the V1/V2 region of the human immunodeficiency virus type 1 gp120 glycoprotein on subunit association, syncytium formation, and recognition by a neutralizing antibody. J. Virol. 67:3674–3679. 33. Syu, W. J., J. H. Huang, M. Essex, and T. H. Lee. 1990. The N-terminal
region of human immunodeficiency virus envelope glycoprotein gp120 contains potential binding sites for CD4. Proc. Natl. Acad. Sci. USA 87:3695–3699.
34. Thali, M., C. Furman, D. D. Ho, J. Robinson, S. Tilley, A. Pinter, and J. Sodroski.1992. Discontinuous, conserved neutralization epitopes overlap-ping the CD4-binding region of human immunodeficiency virus type 1 gp120 envelope glycoprotein. J. Virol. 66:5635–5641.
35. Thali, M., U. Olshevsky, C. Furman, D. Gabuzda, M. Posner, and J. Sodroski.1991. Characterization of a discontinuous human immunodefi-ciency virus type 1 gp120 epitope recognized by a broadly reactive neutral-izing human monoclonal antibody. J. Virol. 65:6188–6193.
36. Wong-Staal, F., G. M. Shaw, B. H. Hahn, S. Z. Salahuddin, M. Popovic, P. Markham, R. Redfield, and R. C. Gallo.1985. Genomic diversity of human T-lymphotropic virus type III (HTLV-III). Science 229:759–762. 37. Wyatt, R., N. Sullivan, M. Thali, H. Repke, D. Ho, J. Robinson, M. Posner,
and J. Sodroski.1993. Functional and immunologic characterization of human immunodeficiency virus type 1 envelope glycoproteins containing deletions of the major variable regions. J. Virol. 67:4557–4565.
38. Yu, X. F., S. Ito, M. Essex, and T. H. Lee. 1988. A naturally immunogenic virion-associated protein specific for HIV-2 and SIV. Nature (London) 335:262–265.
39. Yuan, X., X. Yu, T. H. Lee, and M. Essex. 1993. Mutations in the N-terminal region of human immunodeficiency virus type 1 matrix protein block intracellular transport of the Gag precursor. J. Virol. 67:6387–6394.