0022-538X/84/070192-07$02.00/0
Copyright©) 1984, American SocietyforMicrobiology
Characterization of
a
Temperature-Sensitive Defect of
Enterovirus
70:
Effect of Elevated Temperature
on
In
Vitro Transcription
KIKUKO MIYAMURA,* NAOKAZU TAKEDA, AND SHUDO YAMAZAKI
Central Virus Diagnostic Laboratory, National Institute of Health, Musashimurayama, Tokyo 190-12, Japan
Received 22 December 1983/Accepted 21 March 1984
A crudereplication complex prepared from enterovirus 70-infected cellswasusedtostudy the
temperature-sensitivecharacteristic of the virus. The complex showedatemperaturesensitivity in the in vitro incorporation ofradiolabeled ribonucleoside triphosphate. The endonuclease itself did not account for therestricted RNA synthesisatthenonpermissivetemperature. Analyses of the in vitro products by both gel electrophoresis and sucrose density gradient centrifugation showed that the complex synthesized three typesof viral RNA only when incubated forashortperiod of timeatthe nonpermissivetemperature.When thereplication complexwas
treated with a detergent (deoxycholic acid), incorporation of ribonucleoside triphosphate into RNA at the permissive temperaturewasreduced to the level of thatatthenonpermissive temperature. In addition, the in vitro RNA synthesis by the enterovirus 70 replication complexatthepermissivetemperaturerequiredahigher
concentration of ATP than of other ribonucleoside triphosphates, whereas suchapreference for ATPwasnot
found in the reaction atthe nonpermissive temperature. The results indicatethat the initiation stepof RNA synthesis by the complex is blocked at the nonpermissive temperature. The possible implications ofthese findings arediscussed.
Enterovirus 70(EV70), acute hemorrhagic conjunctivitis
virus, is a naturally occurring temperature-sensitive (ts) virus, which suddenly appeared in 1969 among people in a
pandemic fashion(14, 18, 20). Itis interestingtoinvestigate
the possible mechanism by which the virus appeared so
suddenly and with such a unique pathogenicity as that
causingconjunctivitis.Sofar,onlytwotypesofpicornavirus
are known to be majorcauses ofconjunctivitis, i.e., EV70
and avariant of coxsackievirus A24(19). Since thets nature of the virus is reasonably consideredtofacilitateits growth inconjunctiva, westudiedthemechanism by which the virus
exhibits the tscharacteristic in its growth and attempted to locate the site of the ts lesionin viralreplication.
Inprevious studies (22, 25), the following were found. (i) Novirus-specific RNAwas synthesized in the
EV70-infect-ed cells at anonpermissivetemperature. (ii)Inthe presence
of preformed viral mRNA, synthesis and the subsequent cleavage of virus-specific proteins occurredat a
nonpermis-sive temperature. (iii) Early translation ofviral mRNA at a
nonpermissivetemperaturewasdemonstratedby viral
poly-meraseactivity in theinfected cells. From theseresults, we
concluded that the ts lesion of the virus resided in its transcriptionalstage. Inthe presentstudy,RNAsynthesis of
EV70 at anonpermissive temperature was analyzed in the
cell-free system by using a crude replication complex
pre-paredfromEV70-infected cells.
MATERIALS AND METHODS
Chemicals. BovinepancreaticRNase(type1A), unlabeled
ribonucleoside triphosphates, and deoxycholic acid (DOC)
were purchased from Sigma Chemical Co., St. Louis, Mo. Trichloroaceticacid,sodiumdodecylsulfate(SDS),andTris were from Wako Pure Chemical Industries, Osaka, Japan.
Phosphoenolpyruvateand pyruvate kinasewerefrom
Boeh-ringerMannheimBiochemicals, Mannheim,WestGermany. Dithiothreitolwasfrom SeikagakuKogyo, Tokyo.
[3H]GTP
(10 Ci/mmol),[3H]CTP
(22 Ci/mmol), andEn3Hance
were from New England Nuclear Corp., Boston, Mass. Seakem* Correspondingauthor.
Agarose wasfrom FMC Corp., Marine Colloids Div., Rock-land, Maine. Actinomycin D was from Makor Chemicals,
Jerusalem, Israel.
Cells and viruses. The viruses used were the prototype
J670/71 strain ofEV70 and its warmth-adapted clone
artifi-cially induced by serialpassagesofthe prototypeatelevated temperatures (10 passages at each of 37 and 39°C and
another10 passagesat40°C);theclonegrewbetterat39 than at 33°C, and its optimaltemperature for in vivogrowthwas
38°C. The poliovirus included was the Mahoney strain of
wild-type 1. The viruses were propagated in LLC-MK2 cell monolayers with Eagle minimum essential medium.
Preparation of crude replication complex. The procedure wasessentially the same as thatof Yin and Knight (30). The cells were infectedwith the virus at amultiplicityofca. 40
PFU/cell. The EV70-infected cells were incubated at 33°C,
whereas the cellsinfected with thewarmth-adaptedstrain or
polioviruswereincubated at37°C for5 h.The infected cells werescraped,pelletedby low-speedcentrifugation(600x g)
for 5 min, and stored at -70°C. Approximately 4 x 108 frozen cellsweresuspended in 10 ml of cold buffer (0.01 M
Tris, 0.01 M NaCl, 0.0015 M MgCl2, pH 7.4)to be swollen
for10minonice andwereruptured by15strokes inaglass
Dounce homogenizer. The unbroken cells and the nuclei
wereremoved bycentrifugationat800x gfor5min, and the supernatant fraction was centrifuged at 30,000 x g for 20 min.Thepelletwassuspendedin4mlof buffer(0.05MTris,
0.01 MNaCl, pH 8.0)and storedat -70°C. This
suspension
contained ca. 1 mgofproteinper ml and servedasthe viralreplication complex unlessotherwise indicated.
Assay for polymerase activity. A 50-,ul protein of the
replication complex was mixed with an equal volume ofa standard assay mixture (double concentration of the final reaction mixture) containing the following constituents un-less otherwise indicated: 100 mM Tris buffer (pH 8.0); 40 mM KCl; 10mM
MgCI2;
2.5mMphosphoenolpyruvate; 13 mMdithiothreitol; 10jg
ofactinomycinD perml;50 ,ugof pyruvate kinase per ml; 300 ,uM each of ATP, CTP, andUTP; and 20 FtM
[3H]GTP
(20p.Ci/ml).
The mixtures were incubated in a water bath, and the temperatures were192
on November 10, 2019 by guest
http://jvi.asm.org/
maintained within an error of less than ±0.05°C. After a
desired period, the incubation was terminated by spotting
the mixture onto DEAE-cellulose disks (Whatman DE 81; Whatman, Inc.,Clifton,N.J.), whichwerethensoaked for 5 min at room temperature in a2.5% Na2HPO4 solution. The wash was repeated four more times in Na2HPO4 and twice each in waterand ethanol. Thedried disks werecounted in toluene-based scintillation fluid.
Sucrose density gradient analysis ofin vitro RNA product. The enzyme reactionwasterminated by the addition to the
reactionmixture ofan equal volume ofTNEbuffer (0.01 M
Tris, 0.1 M NaCl, 0.001 M EDTA, pH 7.4) containing 1% SDS.
RNA was extracted three times, each with an equal
volume of a phenol mixture (50 parts phenol,
48
partschloroform, and 2 partsisoamylalcohol,saturated with TNE buffer andcontaining 1 mgof 8-hydroxyquinoline perml) at roomtemperature.The aqueousphasewasadjustedto0.2 M
sodium acetate-70% ethanol, and RNA was precipitated overnight at -20°C. RNA was collected by centrifugation
(10,000 x g, 15 min), dried in vacuo, dissolved in acetate
buffer (0.1 M sodium acetate [pH 5.3], 0.1% SDS, 0.1 M
NaCl, 0.001 M EDTA), and cast on 12 ml ofa 5 to 25% (wt/wt) linear sucrose density gradient in acetate buffer.
Centrifugation wascarriedout at200C for15 hat50,000x g in a Hitachi RPS 40T rotor. Thegradient was collected in
0.5-mlfractions from the bottom. Eachfractionwasdivided
into two equal portions. One portion of each fraction was
diluted with10volumes of 2x SSC(lx SSC is 0.15 M NaCl
plus0.015 M sodium citrate, pH 7.4) and treated with 40
p.g
of bovinepancreatic RNase A per mlat37°Cfor30 min. The other portion remained untreated. RNA from each portion wasprecipitatedwith5%trichloroacetic acid in the presenceofbovine serum albumin and filtered through a glass-fiber
7
6
-2
A..
filter (WhatmanGF/C)before counting with a liquid scintilla-tion counter.
Agarose gel electrophoresis. Electrophoresis was carried outunder nondenatured conditions asdescribed by Hewlett et al. (13). Agarosewasmelted in Ebuffer (40 mM Tris, 10 mM acetate, 1 mM EDTA, pH 7.3) containing 10% (vol/vol) glycerol. Agarose gel (1%, wt/vol) wascast in a horizontal slab gel apparatus (130 by 130 by 1 mm) and prerun in E buffer at 70 V for 1 h. The ethanol precipitates of RNA
species werewashed with95% ethanol, driedin vacuo,and
dissolved in 10 ,ul of 0.1 M Tris(pH 7.5) containing0.15 M NaCl and0.5%SDS. Thesamples which weremixedwith 5 ,ulof 60% (wt/vol) sucrose containing 0.2% (wt/vol)
bromo-phenolbluewereeachlayeredontothegel under therunning buffer. Electrophoresis wascarried out at 75 Vfor 4.5 h at room temperature. Forfluorography,gel slabs were
impreg-natedwithEn3Hance,dried in vacuo,and exposed toSakura
X-ray filmfor 4daysat -70°C.
RESULTS
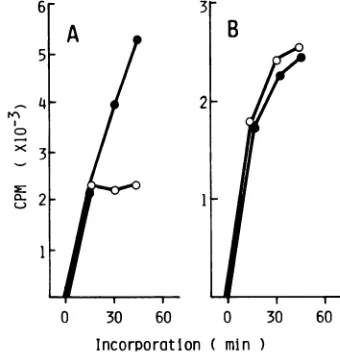
Temperature dependency of viral RNA synthesis. EV70 does not synthesize virus-specific RNA in infected cells at 39°C. The effects of temperature on viral RNAsynthesis in
thecell-free system were studied from the kinetics of
incor-porationof[3H]GMP intoRNAproducts(Fig. 1). The EV70 warmth-adapted strain and the Mahoney strain of poliovirus type 1 were included in the experiment as controls; both
viruses can replicateat 39°Cin vivo.
The incorporation of
[3H]GMP
by the polymerasecom-plexextractedfromJ670/71-infected cellswas almostlinear
at 33°C for 60min. At 37 and 39°C, the reactionduring the
first 15 min proceeded similarly to that at 33°C but was
severely impaired thereafter. The incorporation entirely
ceasedat 39°Cafter incubation for 30 min.Thus,the pattern
0 30 60 90 120 0 30 60 90 120 0 30 60 90 120
incorporation
( min )FIG. 1. Effect of temperature on [3H]GMP incorporation. LLC-MK2 cell monolayers were infected with strain J670/71, its warmth-adaptedstrain, or the Mahoney strainofwild poliovirus1andincubated for 5 h. The infected cells were collected and disrupted to make the replication complex. Portionsof reaction mixture (100,ul)containing50,udof each of thereplication complexand thestandard assaymixture wasincubatedat 33, 37, or39°C.Samplesweretaken at 15-minintervals,and the totalradioactivity incorporatedinto the RNAproductwas
determined. Theradioactivity ofthecomplexat0hwasconsideredtobenonspecific bindingandwassubtractedfrom each value.(A)J670/71, (B)warmth-adapted strain, (C) poliovirus. Incorporationat33(0), 37(A), and39°C (0).
on November 10, 2019 by guest
http://jvi.asm.org/
[image:2.612.95.524.424.673.2]of the incorporation curve at each temperature seemed to faithfully mimic thekinetics of RNA synthesis as well as of viral growth in the infected cells (21, 25). In viral RNA synthesis by the replication complex preparedfrom warmth-adapted and poliovirus-infected cells, the linear incorpo-ration curves of [3H]GMP which were similar to that of the J670/71 complex were obtained at 330C. The inhibition of the incorporation at both 37 and 39°C, however, was much less than that of the J670/71 complex. Actually, the reaction proceeded progressively even at 390C, although the incorpo-ration was markedly reduced.
Since the incorporation by the EV70 complex entirely ceased at 39°C after incubation for 30 min as described above, the following experiment was carried out to deter-minewhether the replication complex contained a thermola-bile component or whether a cellular endonuclease was preferentially activated at 39°C, resulting in the digestion of RNA synthesized during the reaction. The complex pre-pared from J670/71-infected cells was heated for 60 min at 39°C. The heated complex was then mixed with a reaction mixture and assayed for the heat-resistant transcriptional activity from [3H]GMP incorporation for 30 min at the permissive temperature. The complexes of the warmth-adapted clone and the poliovirus were also tested as con-trols. When incubated at 390C for 60 min, the J670/71 complex retained40% of the [3H]GMPincorporation of the unheated control. At the same time, incorporation by the complexof the warmth-adapted clone also was reduced to a similar extent (to 38% of the unheated control) and that of the poliovirus was even more severely impaired (to29% of the unheated control) after heating at 39°C for 60 min (data notshown). Thus, the temperature defect of EV70 replica-tion could not simply be explained by the thermolability of itspolymerase or by activation of the cellularendonucleases at the elevated temperature.
Analysis of RNA products of the replication complex. It has beendemonstrated that crude replicationcomplexes derived from thepoliovirus-infected cells synthesize three speciesof viralRNA, i.e., single-stranded RNA, replicative intermedi-ate, and replicative form (RF) (12). The products of the EV70 replication complex synthesized in the cell-free sys-tem at both permissive and nonpermissive temperatures were analyzed by agarose gel electrophoresis. Figure 2 shows the comparative gel patterns of RNA products syn-thesized in 30, 60, and 120min at both 33 and 39°C; for the product at 33°C, all three species of viral RNA are demon-strated. The longer the incubation period was, the higher
was theyield. The products at390C also gave clear RF bands with similar intensities to those at 33°C. However, the replicative intermediate or 35 S RNA wasbarelydiscernible
in the products incubated at 39°C. The results were also
confirmed by acrylamide gelelectrophoresis of the products (data not shown).
Tocharacterize the in vitro RNA products more quantita-tively, sucrose gradient analysesof the productssynthesized in vitroat both permissive and nonpermissive temperatures were carried out(Fig. 3). In the products produced at 330C,
increased amounts of viral RNA ofboth 35S and 20Sspecies weredemonstrated, confirming the results shown by agarose gel electrophoresis. The products at 39°C also contained both 35S and 20S RNA. However, the amounts of RNA collected from 60- and 120-min incubations were quite
similar to that produced from 30-minincubations,indicating
that synthesis of all RNA species stopped shortly after the initiation ofincubation.
Effect of DOC on polymerase. The treatment ofthe
cyto-33tC
39'C30' 60' 120' 30' 60' 120'
-RI
___ _mm Em --RF
-35S
FIG. 2. Agarose gel electrophoresis of the products of the J670/71replication complex synthesized at permissive and nonper-missive temperatures. Portions (1 ml) of the reaction mixture containing 500 ,ul of the J670/71 replication complex and 10 ,uM [3H]GTP (30
VtCi/ml)
wereincubated at 33 or 39°C. The reaction was terminated after various periods of incubation by the addition of TNEbuffer containing 1% SDS, andRNAwasextracted from each reaction mixture. Electrophoresis was performed on1%agarosegel slabs for4.5 h at 75 V, andthegel was fluorographed. The viral RNAinthegel waslocated by coelectrophoresis of in vivo labeled viral RNA. The temperature and the incubation period (in minutes) foreach productareshown inthefigure.plasmic membrane fraction of the infected cells with a
detergent such as DOC orNonidet P-40
effectively
dissoci-ates the virus-induced
RNA-polymerase
complex
(9). It has been suggested that in vitro viral RNAsynthesis by
adetergent-solubilized complex
might
belimitedtoelongation
of the RNA chains already started (3, 10), lacking theinitiation of transcription. Therefore, further evidence was soughtbyanalyzing the effect ofanelevated temperature on the reaction by the complex treated with a
detergent.
Theaddition of 0.25% DOC to the
replication
complex
of theJ670/71strain, followed
by
heating
for 3 minat33°C,
reduced the incorporation ofGMP at33°C
to the level of incorpo-ration at 39°C, thusentirely
abolishing
thepreferential
incorporation ofGMP at apermissive
temperature(Fig. 4).
The result indicates that a certain functional factor orstructure required for complete RNA
synthesis
atapermis-sive temperature is
destroyed
by
thedetergent.
Atthe sametime, the result suggests that the
elongation
process is notprevented at an elevated temperature.
Requirement of ATP for in vitro transcription. In the
procaryotic
DNA-dependent
RNApolymerase
system, the two events during transcription, initiation andsubsequent
chain elongation,havebeendifferentiatedby
kineticanalysis
of the initial
velocity
of the reaction at different substrate concentrations (1).Thedifferentiation has beenbasedonthe
facts that initiation ofin vitrotranscription
demonstrates astringent preferenceforpurine ribonucleoside
triphosphates
(rNTPs) at a high concentration and, in contrast, that thesubsequent RNA chain elongation
requires
low levels ofpurinerNTPs. Consequently, the KmforATP atinitiation is much higher than that for the same nucleoside
during
elongation. The sameprinciple
has been used tostudy
in vitro RNAsynthesis in thevesicular stomatitis virus system (29) and the rotavirus system(24).
In the presentstudy,
weI..X,
on November 10, 2019 by guest
http://jvi.asm.org/
[image:3.612.355.513.70.264.2]rf 3
0
X
0--I Z:
xL
_...
3
2
C
28S 18S
D
5 10 15 20 5 10 15 20
Fraction Fraction
FIG. 3. Sucrosedensity gradient analysis of products of the J670/71 replication complex. The reaction mixtures in 1-mlvolumeseach containing 500 ,ul of the J670/71 replication complex and 10 ,uM[3H]GTP (20 ,uCi/ml)wereincubatedat33 (A,B) and39°C (C, D) for various periodsof time.RNA wasextracted and fractionatedon a15 to30%sucrosegradient. One half ofeachfractionwastreatedwith40,ugof RNaseper ml (BandD), and allfractions were determined for acid-insoluble radioactivity. Sedimentation was from right to left. Arrows indicate theposition of 18S and 28SrRNApeaks. Products of incubation for30min( ),60mi ( ), and 120mi ( ).
alsousedkineticanalysistodetermine whether theinitiation eventis involved in the reaction in theEV70system atboth
permissive and nonpermissivetemperatures.
The dependence of the rate of reaction on the rNTP
concentration was examined for each of four rNTPsduring
the first 30 min of incubation, and the apparent K,, was
determined fromdouble-reciprocal plots. The ATP
concen-tration required for in vitro transcription by the EV70
replication complex showedbiphasic double-reciprocal plots (Fig. 5). At a low rNTP concentration, the apparent Km
valueswerealmostidentical forallfourrNTPs, i.e., 9 to 12 FiM. There was, however, a sharp increase in the rate of transcriptionwhen the concentration of ATPwasincreased to ca. 20 ,uM or more in the reaction mixture, whereas no
changewasobserved in the apparentKmfor the other three rNTPs at any concentration tested. Consequently, the Km
forATPattheseconcentrations wasapproximately fourfold
greater, i.e., 43 ,uM, than thatforthe other three rNTPs or
for ATP at concentrations less that 20 ,uM. The results indicate thatanincreasedfrequency of initiationwas expect-ed tooccur at such high ATPconcentrations.
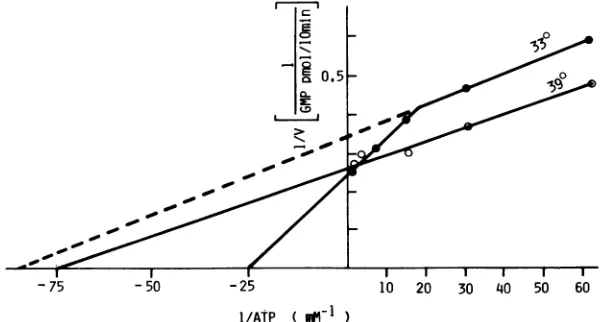
To examine the effect ofan elevated temperature on the
reaction, the apparent Km at 39°C was detertmined. The reaction was carried outfor 10 min, duringwhichperiod of
timeidenticalratesof rNTPincorporation wereobservedat 33 and 39°C (Fig. 1). No increase in the velocity of the reaction was observed at 39°C, even at a higher ATP
concentration (Fig. 6). The result indicates that an
ATP-dependent process, probably initiation, of viral RNA
tran-scription was inhibited atthe nonpermissive temperature.
DISCUSSION
The results reported here show that the transcription in thecell-freesystembyacrudereplication complex prepared
-CL)
B
0 30 60 0 30 60
IncorPoration
( min )FIG. 4. Effect ofDOCtreatment onJ670/71replication complex. The J670-71complexwas mixed with anequalvolume ofTris buffer (pH 8.0) containing 5% DOCand then heatedfor3min at33'C.The DOC-treated complex was mixed with an equal volume of the standardassaymixture,andincorporation of
[3H]GMP
at33(-)and 39°C(0)
wasdetermined asdescribedin thelegendforFig. 1. (A) Untreatedcomplex, (B) DOC-treatedcomplex.on November 10, 2019 by guest
http://jvi.asm.org/
[image:4.612.91.524.71.372.2] [image:4.612.349.519.492.669.2]II I I
-100 -50 50 100
l/rNTP (
r-l1
)FIG. 5. Effectof the rNTPconcentrationon the EV70 RNApolymerase activity.The 100-,u reaction mixtureseach contained50,u1 ofthe EV70 replication complex and the fourrNTPs. [3H]GTPwas added at 10 ,uM(820cpm/pmol), except when GTPdependence was to be measured. ForGTP dependence,[3H]CTPwasused at 10,uM (870cpm/pmol). OtherrNTPs wereincludedat150p.Mexcept XTPwhichwas varied(5to 500p.M). The reaction mixtures wereincubated at33'C for30min.Theradioactivity incorporated intothe RNA product was assayed and expressed as a double-reciprocal plot ofthe rate ofreaction (picomoles of [3H]GMP incorporated/30min) versus the XTP concentration.TheapparentKm values obtainedfor each XTP were asfollows:ATPreactions(0),K,, = 42p.MinthereactionathigherATP concentrations than20p.Mand12p.MatlowerATPconcentrations than20,uM; GTP reactions(A),K,, = 11 FiM;CTPreactions (A), K,, =10 ,uM;UTPreactions (0),K,, = 9 AM.
from EV70-infected cells mimicsfaithfully the in vivo tran-scription, confirming our previous findings that the ts event of the virus ingrowth resides in its transcription. Thus, the system has proved to be useful in studying the ts nature of thevirus.
To analyze the transcriptional events, four identifiable steps have been considered: binding of polymerase to a template, initiation of transcription, elongation of the RNA strand, and its termination. Although itis well known that thepolymerase of picornavirus synthesizes its RNA in a cell-free system, little has been reportedon the involvement of theinitiationstep in itstranscription. Recently, Takegamiet al. (26) statedthat acrudemembrane fraction isolated from
poliovirus-infected cells synthesizes a nucleotidyl
protein,
,a-p
-Ia- 0
i.e., the 5' termini of a nascent RNA strand. In this report, wealso presentedexperimental evidence which supports the
involvement of the initiation step in transcription by the
crude replication complex prepared from EV70-infected
cells.
First, from analyses ofthe in vitro products by sucrose
gradient centrifugation andby gelelectrophoresis, all types
ofviral RNAwerefound to be continuously synthesizedat the same rate during the test period, suggesting de novo initiation of RNAtranscription. Second, another support for
involvement of the initiation was provided by the kinetic study onthe
requirement
ofATPin thereaction. The studywas based on the fact that the two eventsduring
transcrip-tion, invitro initiationandsubsequent chain elongation,are
-75 -25 10 20 30 40 50 60
l/ATP ( mm-l )
FIG. 6. Effect of elevatedtemperature onapparentK,, forATPof the EV70RNApolymeraseactivity.Thereactionmixtures in 100-p.l quantitieseachcontaining50p.loftheEV70replication complex,10p.M
[3H]GTP
(728cpm/pmol),150p.Meach ofCTPandUTP,andvarious concentrations(5to500p.M)
ofATPwereincubatedfor 10 minat33 and39°C.Theapparent K,, valuesweredeterminedasdescribed in the legendforFig. 5.Reaction at33°C(0),K,,=40p.Min thereactionat ahigherATPconcentration and 12p.Mat alower ATPconcentration;reactionat39°C(0),K,,, = 13 p.M.
on November 10, 2019 by guest
http://jvi.asm.org/
[image:5.612.97.526.75.269.2] [image:5.612.163.468.521.682.2]successfullydifferentiatedbytheirrequirementsfor different ATP concentrations, such as those in a procaryotic
DNA-dependent RNApolymerase system(1)and in the vesicular stomatitis virus system (29). In our experiments, the Km for ATP at ahigher ATP concentration in in vitro EV70 tran-scription was nearly fourfold greater than that for the other three rNTPs, suggesting the occurrence of the initiation of
transcription on the template. Interestingly, at lower ATP
concentrations, the Km was reduced to an almost identical
leveltothatforthe other rNTPs.Thisbiphasicpatternofthe
transcriptionrateondifferent levels of ATP concentration is
quitesimilartothatreported byTestaand Banerjee(29)with
the preinitiated complexof vesicularstomatitis virus. Such
resemblance inthepatternofthetranscriptionratebetween
the two complexes was consistent with the fact that the EV70 replication complex consisted largely ofendogenous template-binding polymerase preformedinthe infectedcells, comparableto that in the
preinitiated complex
ofvesicularstomatitis virus.
The results of the
comparative experiments
ontheeffectsof different temperatures strongly indicated the lack of
initiation oftranscriptionatthe
nonpermissive
temperature. In contrast tothe continuoussynthesis
ofthree RNAmole-cules atthe permissive temperature, RNA
synthesis
at thenonpermissivetemperatureceased inashortperiod of time, andthepreferential requirementfor ATP in the reactionwas not observed at the nonpermissive temperature. Further-more, basedon the
suggestion
ofotherinvestigators
(3, 10,17) that the detergent-treated complex supports only chain
elongation, identicalRNAsynthesisatbothtemperaturesby
theDOC-treatedcomplexprepared from EV70-infectedcells
indicates that the
elongation
process is notprevented
at anelevated temperature.
Fromthese results, in vitro RNA synthesis bythe EV70
complex at the
nonpermissive
temperature could beex-plained as follows. During a short period of time after incubation, the RNA chain elongation,
having
alreadystart-ed, continuedtoproceed, and nascent
single-stranded
RNAdissociated from thetemplate,
producing
35SRNA. Howev-er,thesubsequentlyelongating
strand couldnotbe released from the template, possibly due to base pairing with thetemplate bysome yet unknown mechanism.Thus, all
repli-cative intermediates turned to RF, having no newgrowingstrand onit, resultingin thecompleteblockofRNA synthe-sis. Many in vivo and in vitro studies of
picornavirus
replication have shownthat single-stranded RNA synthesisis extremely susceptible to such factors as an elevated temperature (4), detergents (10, 16, 17), guanidine (28), hydroxybenzimidazol (7), and the antibody directed against
thehost cellfactor(5, 8). In most cases,
synthesis
ofRFwas not affected. Thus block ofsingle-stranded RNA synthesisand accumulation of RF in viral transcription, as seen in
EV70 replication at the nonpermissive temperature, might be a phenomenon rather frequently observed under the
condition in which RNA synthesis of picornavirus is im-paired.
Recent advances in studies on theinitiationstepof
picor-navirus RNA replication in vitro have been focused on the roles of the two factors which are associated with de novo
synthesisof RNAreplicase.Oneis a hostfactor(s)which has been shown to be necessary for the initiation of RNA
synthesis by the purified poliovirus replicase (2, 6, 8). The
factor was found to bind to viral polymerase in vitro (5). Thus it might be possible to inhibit the transcription if the
interactionof the hostfactorwith another component of the
EV70-replication complex is susceptible to an elevated tem-perature.
Another factor which is considered to play a role in
initiation is a small, virus-coded protein, VPg, covalently bound to the 5' termini of picornavirus RNA (11, 15).
Nomoto et al. (23) proposed the following model in which VPgfunctionsas aprimer forRNA synthesis. DuringRNA
strandelongation,theRNA-linked VPg remains attachedto
thepolymerase, preventing the nascentstrandfrom anneal-ing to the complementarytemplate RNA. When the
RNA-linked VPg dissociates from polymerase after the initiation ofRNA synthesis, the nascent strand may anneal to
tem-plate RNA, generating double-stranded RNA, RF. Accord-ing to this model, the effect ofan elevatedtemperature on EV70 transcription may be explained as follows. If the linkage between VPg and polymerase is susceptible to an
elevated temperature, VPg
might
dissociate from polymer-ase during elongation at a nonpermissive temperature,re-sultingin thegeneration ofRF.Another
possibility might
be adefectin thecleavage oftheprecursorpeptide
ofVPgat anelevated temperature.
Takegami
et al. (26, 27) recently presenteda model in which VPg is present in the initiation complex as part ofa precursorpolypeptide
and is cleavedfrom it after the initiation ofRNA synthesis. Thefailure of this cleavage at an elevated temperature could inhibit the
formation ofafunctional form of VPg.
Further clarification is required for a conformational as well as afunctional relationship of VPgand thepolymerase of EV70 with other components present in the initiation complex, i.e., functional templates andhostfactor(s).
ACKNOWLEDGMENTS
We aregratefultoT.Yamashita, Daiichi College of Pharmaceuti-calSciences, for his useful advice forexperimental procedures.
This workwassupportedinpartbyaGrant-in-Aidfor Scientific Research from the Ministry of Education, Science and Culture, Japan.
LITERATURECITED
1. Anthony,D.D., C.W.Wu,and D. A.Goldthwait. 1969. Studies with the ribonucleic acid polymerase. II. Kinetic aspects of initiation andpolymerization.Biochemistry8:246-256. 2. Baron,M.H.,and D.Baltimore. 1982. Purification and
proper-ties ofahost cell protein requiredfor poliovirus replicationin vitro.J. Biol. Chem. 257:12351-12358.
3. Baron,M.H.,and D. Baltimore. 1982. Invitrocopyingofviral positive strand RNA by poliovirus replicase. J. Biol. Chem. *257:12359-12366.
4. Cooper,P.D.,D.Stantek,and D. F.Summers. 1970.Synthesis of double-stranded RNA by poliovirus temperature-sensitive
mutants. Virology40:971-977.
5. Dasgupta,A. 1983.Antibodytohostfactorprecipitates
poliovi-rusRNApolymerase frompolio-infectedHeLa cells. Virology 128:252-259.
6. Dasgupta, A.,P.Zabel,and D. Baltimore.1980. Dependenceof theactivityofthepoliovirus replicaseon ahost cellprotein.Cell 19:423-429.
7. Dmitrieva, T.M.,andV. I. Agol. 1974. Selective inhibition of thesynthesis ofsingle-strandedRNAofencephalomyocarditis virus by 2-(a-hydroxybenzyl) benzimidazole in cell-free
sys-tems.Arch. Gesamte Virusforsch. 45:17-26.
8. Dmitrieva, T. M., M. V. Shcheglova, and V. I. Agol. 1979. Inhibition of activity of encephalomyocarditis virus-induced RNA polymerase by antibodies against cellular components. Virology92:271-277.
9. Ehrenfeld, E., J.V.Maizel,andD. F.Summers. 1970. Soluble RNApolymerase complexfrompoliovirus-infectedHeLacells. Virology40:840-846.
10. Etchison, D.,and E.Ehrenfeld. 1981.Comparisonofreplication complexessynthesizing poliovirusRNA. Virology111:33-46. 11. Flanegan, J. B.,R. F.Pettersson,V.Ambros,M.J. Hewlett,and
D.Baltimore. 1977. Covalent linkage ofaprotein to adefined nucleotide sequenceatthe5'-terminus of virionandreplicative
on November 10, 2019 by guest
http://jvi.asm.org/
intermediate RNAsof poliovirus. Proc. Natl. Acad. Sci. U.S.A. 74:961-965.
12. Girard, M. 1969. In vitro synthesis of poliovirus ribonucleic acid: roleofthereplicative intermediate. J. Virol. 3:376-384. 13. Hewlett, M. J., S. Rozenblatt, V. Ambros, and D. Baltimore.
1977. Separation and quantitation of intracellular forms of poliovirus RNA by agarose gel electrophoresis. Biochemistry 16:2763-2767.
14. Kono, R., A. Sasagawa, K. Ishii, S. Sugiura, M. Ochi, H. Matsumiya, Y. Uchida, K. Kameyama, M. Kaneko, and N. Sakurai. 1972. Pandemic of new type ofconjunctivitis. Lancet i:1191-1194.
15. Lee, Y. F., A. Nomoto, B. M. Detjen, and E. Wimmer. 1977. A protein covalently linked to poliovirus genome RNA. Proc. Natl. Acad. Sci. U.S.A.74:59-63.
16. McDonnel, J. P., and L. Levintow. 1970. Kinetics ofappearance oftheproducts ofpoliovirus-inducedRNApolymerase. Virolo-gy 42:999-1006.
17. Meyer, J., R. E. Lundquist, and J. V. Maizel, Jr. 1978. Structur-al studies of the RNAcomponent of thepoliovirus replication complex. II.Characterization byelectronmicroscopy and auto-radiography. Virology 85:445-455.
18. Mirkovic, R. R., R. Kono, M. Yin-Murphy, R. Sohier, N. J. Schmidt, and J. L. Melnick. 1973. Enterovirus type 70: the etiologic agentofpandemicacute haemorrhagicconjunctivitis. Bull.W.H.O. 49:341-346.
19. Mirkovic, R. R., N. J. Schmidt, M. Yin-Murphy, and J. L. Melnick. 1974. Enterovirus etiology of the 1970 Singapore epidemic of acuteconjunctivitis. Intervirology4:119-127. 20. Miyamura, K., A. Sasagawa, E. Tajiri, and R. Kono. 1976.
Growth characteristics of acute hemorrhagic conjunctivitis (AHC)virusinmonkeykidneycells. 11.Temperaturesensitivity oftheisolates obtainedatvariousepidemicareas.Intervirology
7:192-200.
21. Miyamura, K., S. Yamazaki, E. Tajiri, and R. Kono. 1974. Growth characteristics of acute hemorrhagic conjunctivitis (AHC) virus in monkey kidney cells. I. Effectof temperature on viral growth. Intervirology4:279-286.
22. Miyamura, K., S. Yamazaki, E. Tajiri, and R. Kono. 1978. Growth characteristics of acute hemorrhagic conjunctivitis (AHC) virusinmonkeykidneycells. III.ViralRNAsynthesisat permissive and nonpermissive temperatures. Intervirology 9:206-213.
23. Nomoto, A., B. Detjen, R.Pozzatti, and E. Wimmer. 1977. The location of the polio genome protein in viral RNAs and its implication forRNA synthesis.Nature (London) 268:208-213. 24. Spencer, E., and M. L. Arias. 1981. In vitro transcription
catalyzed by heat-treated humanrotavirus. J.Virol.40:1-10. 25. Takeda, N., K. Miyamura, R. Kono, and S. Yamazaki. 1982.
Characterization ofatemperature-sensitive defect of enterovi-rustype 70. J.Virol. 44:98-106.
26. Takegami, T., R. J. Kuhn, C. W. Anderson, and E. Wimmer. 1983. Membrane-dependenturidylylation of thegenome-linked protein VPg of poliovirus. Proc. Natl. Acad. Sci. U.S.A. 80:7447-7451.
27. Takegami, T., B. L. Semler, C. W. Anderson, and E. Wimmer. 1983.Membranefractionsactiveinpoliovirus RNAreplication contain VPg precursorpolypeptides. Virology 128:33-47. 28. Tershak, D. R. 1982. Inhibition of poliovirus polymerase by
guanidineinvitro. J. Virol. 41:313-318.
29. Testa, D., and A. K. Banerjee. 1979. Initiation of RNAsynthesis in vitro byvesicular stomatitis virus. J. Biol.Chem. 254:2053-2058.
30. Yin, F. H., and E. Knight, Jr. 1972. In vivo and in vitro synthesis of human rhinovirustype 2ribonucleic acid.J. Virol. 10:93-98.
![FIG.1.determined.was(B)replicationadapted Effect of temperature on [3H]GMP incorporation](https://thumb-us.123doks.com/thumbv2/123dok_us/1426607.95311/2.612.95.524.424.673/fig-determined-was-replicationadapted-effect-temperature-gmp-incorporation.webp)
![FIG. 2.J670/71TNEforcontainingterminatedmissiveviralreactionRNAslabs[3H]GTP Agarose gel electrophoresis of the products of the replication complex synthesized at permissive and nonper- temperatures.Portions(1 ml) of the reaction mixture 500 ,ul of the J670](https://thumb-us.123doks.com/thumbv2/123dok_us/1426607.95311/3.612.355.513.70.264/tneforcontainingterminatedmissiveviralreactionrnaslabs-electrophoresis-replication-synthesized-permissive-temperatures-portions-reaction.webp)