Abstract
Previous studies have investigated the relationship between blood type and pancreatic cancer, but little research has been done to determine how blood type correlates with the development of colon cancer. The fucosyltransferase2 (FUT2) gene is involved in the
with decreased expression of the inflammation marker IL-10. Our findings suggested that the rs516246 FUT2 variant alters bacterial composition and inflammation of the colorectal tract. Further studies are needed in order to analyze additional FUT2 SNPs and confirm their role in colorectal adenomas and cancer.
Introduction
Colorectal cancer is one of the leading causes of cancer death worldwide (Torre et al., 2015) and the third leading cause of cancer death in the United States, in both females and males (American Cancer Society, 2014). This disease usually progresses slowly, and the transition from normal epithelium to adenocarcinoma (Figure 1) can sometimes take 20 years (American Cancer Society, 2014). One of the stages of colorectal cancer is the adenoma (Lao et al., 2011), which was the focus of this study.
Figure 1. The progression of colorectal cancer from normal gut epithelium to polyps/adenomas, to cancer. Source: (Lao et al., 2011)
Colorectal cancer can be hereditary, but it can also affect individuals with no family history of the disease, as in sporadic colorectal cancer (Haghighi et al., 2009). Risk factors for sporadic colorectal cancer include lifestyle choices such as physical inactivity, smoking (Torre et al., 2015) and high meat intake (Roberts-Thomson et al., 1999). The impact that these
that are present (Roberts-Thomson et al., 1999). One genetic polymorphism that could affect the impact of environmental factors is the rs516246 SNP in the fucosyltransferase 2 (FUT2) gene, which is involved in the production of ABH blood group antigens (Jaff, 2010).
of secretion of ABH antigens would change the composition of the protective mucus layer that lines the lumen of the gut (Tong et al., 2014). Therefore, non-secretors may be at a higher risk for colorectal cancer, as research has indicated that the gut microbiome may play a highly significant role in the development of this disease (Louis et al., 2014). The rs516246 SNP in the FUT2 gene is a good indicator of the secretor status of an individual. It can be inferred that those with the wildtype cytosine (C) at the site of this SNP in both alleles have the secretor status, while those with the variant thymine (T) at this same site in both alleles have the non-secretor status (Okunola et al., 2015). Those who are heterozygous and have a C at the rs516246 SNP in one allele and a T at this SNP in the other allele most likely experience some secretion of blood group antigens.
We hypothesized that the rs516246 SNP is associated with the risk of developing colorectal adenomas and that this association may be modified by bacteria, which utilize the antigens produced by FUT2. In addition, the differences in the levels of secretion of these antigens into the mucin of the gut may affect levels of inflammation caused by pathogenic bacteria in the gut.
In this study we examined the relationship between the FUT2 C>T polymorphism (rs516246), microbial gut composition, levels of gene expression of inflammatory markers (Interleukin 10, Toll-Like Receptor 4, Mucin 2) and the presence of colorectal adenomas.
Methods
Study Population
Study V (DHSV). The study was approved by the UNC School of Medicine IRB. Rectal biopsies and blood samples used in this study were obtained during the procedure. After
collection, blood was centrifuged in order to separate the plasma, buffy coat and red blood cells. The buffy coat containing the white blood cells was collected and stored at -80°C until use in the current study.
DNA Extraction from Buffy Coat
DNA Extraction was performed using the Gentra Puregene Blood Kit (Qiagen) according to the kit protocol. Buffy coat samples were thawed at 37 °C, after which residual red blood cells were lysed with Red Blood Cell Lysis solution. The white blood cells were then pelleted by centrifugation and lysed with Cell Lysis solution in order to isolate genomic DNA. The solution was treated with RNase A Solution to achieve RNA-free DNA. Proteins were separated from the DNA with Protein Precipitation Solution, and isopropanol was added to each sample to coalesce the DNA strands. The DNA was then pelleted, dried and later resuspended in DNA Hydration Solution. The concentration of DNA obtained from each sample was measured using a
NanoDrop Spectrophotometer, and the DNA was stored at -4 °C.
Allelic Discrimination Assay
Bacterial DNA Extraction and Illumina Sequencing
Both human and bacterial DNA were extracted from colorectal biopsy samples using a Qiagen DNeasy Blood and Tissue Kit with a modified kit protocol that included lysozyme and bead-beating (McCoy et al., 2013).
The Illumina library was then created using two different PCR reactions. First-step PCR (PCR1) was performed using primers designed to amplify the V2 region of the 16S bacterial rRNA gene. For PCR1, one reaction was performed for each sample using Phusion High-Fidelity Master Mix (Life Technologies, Carlsbad, CA. PCR1 product was diluted 20-fold for use as a template for second-step PCR (PCR2).
PCR2 primers contained an Illumina index barcode sequence, Illumina adapter sequence and a tag sequence. There were two sets of PCR2 primers, and each PCR2 reaction received one
of each, resulting in a dual-indexed product. One reaction was performed for each sample using Phusion High-Fidelity Master Mix.
PCR product was visualized on an E-Gel 96 (Life Technologies, Carlsbad, CA) to check all samples for amplification. All samples with positive amplification were included in the library. All samples were normalized to 25 ng/µl using the SequalPrep Normalization Kit (Life Technologies, Carlsbad, CA). The library was cleaned using AxyPrep Mag Beads (Fadrosh, 2014). Bacterial sequences were filtered for quality control and processed in Qiime (Caporaso, 2010).
Expression of Inflammatory Markers
Human RNA was extracted from colorectal biopsy samples using a Qiagen RNeasy Mini Kit. The RNA was loaded into a chip and run on the Agilent 2100 bioanalyzer using the Agilent 6000 Nano Kit, in order to determine the concentration and fragment size of the RNA in each sample. The RNA for each sample was then diluted to a concentration of 500 ng per 10 μl. A
Promega RQ1 RNase-Free DNase Kit was used to digest any DNA contaminants in the RNA samples. Reverse transcription PCR was performed using an Invitrogen Cloned AMV First-Strand cDNA Synthesis Kit in order to create cDNA from the RNA samples. The cDNA was used as a template for real time PCR, along with a Qiagen RT² qPCR Primer Assay, which included primers specific to the genes of interest. The Primer Assay was mixed with template
cDNA and iTaqTM Universal SYBR® Green and run on a Bio-Rad CFX384 Real Time PCR
Detection System (Kang et al., 2013).
Comparisons of general characteristics of the study population, genotype distribution,
bacterial abundance and gene expression between adenoma cases and controls with no adenomas were assessed by t-tests for continuous variables and chi squared test for categorical variables. Multivariate analysis of bacteria data such as cluster analysis and non-multidimensional scaling was performed using PRIMER-7 software as previously described (Shen et al., 2010).
Multivariate analysis permits the examination of relationships among multiple variables at the same time. P-values <0.05 were deemed significant after adjustment for multiple comparisons.
Results
The purpose of the experiments described above was to determine if there is a correlation between the rs516246 SNP, the types of bacteria present in the gut, the level of inflammation in the gut and the presence of colorectal adenomas. The bacterial profile of the gut was determined through the use of Illumina sequencing of extracted bacterial DNA, and the level of
inflammation of the gut was determined by measuring the level of expression of certain genes involved in inflammation. The results of these experiments, along with the general
characteristics of the study population are given below.
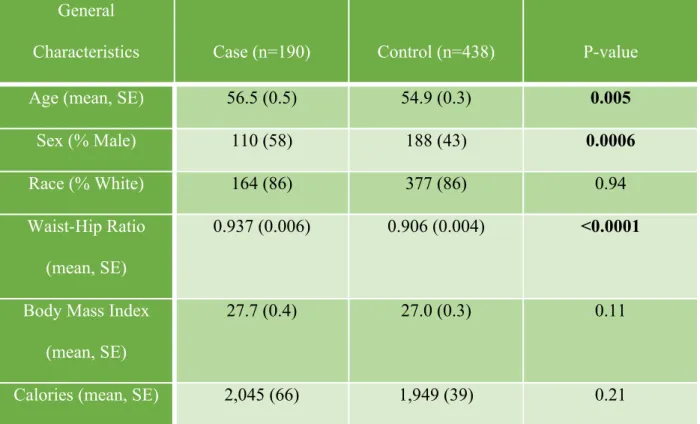
Table 1. Descriptive Characteristics of Study Participants General
Characteristics Case (n=190) Control (n=438) P-value
Age (mean, SE) 56.5 (0.5) 54.9 (0.3) 0.005
Sex (% Male) 110 (58) 188 (43) 0.0006
Race (% White) 164 (86) 377 (86) 0.94
Waist-Hip Ratio
(mean, SE)
0.937 (0.006) 0.906 (0.004) <0.0001
Body Mass Index
(mean, SE)
27.7 (0.4) 27.0 (0.3) 0.11
Calories (mean, SE) 2,045 (66) 1,949 (39) 0.21
The genotype distributions of FUT2 were in Hardy-Weinberg equilibrium, as determined by the equation p² + 2pq + q² = 1, but there was no significant overall association between FUT2 genotype and presence of colorectal adenomas. The distribution of the CC, CT and TT
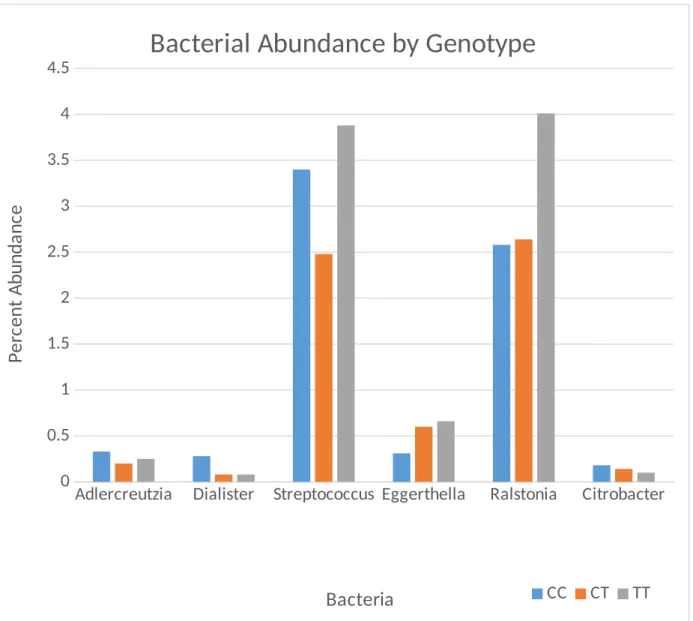
the CC genotype. The genera of bacteria analyzed are divided into two graphs based on the scale of the axis that best fits the data.
Adlercreutzia Dialister Streptococcus Eggerthella Ralstonia Citrobacter 0
0.5 1 1.5 2 2.5 3 3.5 4
4.5
Bacterial Abundance by Genotype
CC CT TT Bacteria
P
e
rc
e
n
t
A
b
u
n
d
a
n
ce
Ruminococcus Blautia 0
5 10 15 20 25
Bacterial Abundance by Genotype
CC CT TT
Bacteria
P
e
rc
e
n
t
A
b
u
n
d
a
n
ce
Figure 3b. The distribution of two genera of bacteria by FUT2 genotype. There was a significant reduction in Ruminococcus (p=0.02) from CC to TT.
There was no statistically significant relationship between the Mucin 2 gene (MUC2) and FUT2 genotype.
Table 2. Gene Expression Levels of Inflammatory Markers in Relation to FUT2 Genotype Inflammatory
Marker
Genotype Case Control P-Value
IL-10 CC (wildtype) 0.79 0.63 0.27
CT 0.64 0.70 0.10
TT 0.46 0.55 0.46
T allele 0.55 0.66 0.06
MUC2 CC (wildtype) 1.24 1.20 0.67
CT 1.02 1.19 0.49
TT 1.02 1.16 0.40
T allele 1.02 1.19 0.45
TLR4 CC (wildtype) 1.03 1.0 0.65
CT 1.19 1.03 0.15
TT 1.03 0.87 0.05
T allele 1.13 0.95 0.03
Discussion
In this study we determined the relationship between a FUT2 polymorphism (rs516246), risk of colorectal adenomas, bacterial composition of the colorectal tract and inflammatory markers. Although previous studies had found a correlation between blood type and pancreatic cancer (Wolpin et al., 2009) and non-secretor status and oral cancer (Ensinck et al., 2013), we did not find any overall association between the rs516246 FUT2 SNP and the risk of developing colorectal adenomas. However, we found that the homozygous variant genotype (TT) is
Previous studies reported significantly lower levels of Ruminococcus and Dialister in the stool of patients with colorectal cancer compared to healthy individuals (Weir et al., 2013) and high levels of Eggerthella in the gut of colorectal cancer patients (Wang et al., 2012). Therefore, the association between the reduced abundance of the two former genera, the increased
abundance of Eggerthella and the variant FUT2 genotype could imply that the rs516246 FUT2 SNP affects the risk of developing colorectal cancer by way of altering the bacterial profile of the gut. Although it is not completely understood how bacteria in the colorectal tract interact with their human host, it is known that variations in the composition of the gut microbiome are present in many diseases (Wu et al., 2015).
Not only does FUT2 affect the composition of the gut microbiome, but it also seems to affect the function of these bacteria in the mucosal layer of the colorectal tract (Tong et al., 2014). These functional changes have been shown to be accompanied by inflammation of the intestinal walls (Tong et al., 2014). In this study we found a link between inflammation and the presence of colorectal adenomas. Compared to controls, cases with the variant T allele displayed lower expression levels of the anti-inflammatory gene IL-10 and higher expression levels of the TLR4 gene, which produces a receptor that triggers the innate immunity response when it is bound to a pathogen, than those with the homozygous wildtype genotype (CC). This may suggest that a complex relationship exists between the gut microbiota, intestinal inflammation, FUT2 and colorectal adenomas.
Although no overall association was found in this study between the rs516246 FUT2 SNP and the risk of developing colorectal adenomas, that does not discount FUT2 as being an important factor in the progression of colorectal cancer, because this SNP is one of many
were not under time constraints, we would have liked to genotype more SNPs within the FUT2 gene.
In summary, our findings suggest that the rs516246 FUT2 variant alters bacterial composition and inflammation of the colorectal tract. Further studies are needed in order to analyze additional FUT2 SNPs and confirm their role in colorectal adenomas and cancer.
Acknowledgements
I would like to thank Dr. Temitope Keku for giving me the opportunity to complete this study, for developing the hypothesis and for her continued support and input throughout the process. I would like to thank Amber McCoy for teaching me all protocols and for completing the bacterial DNA extractions and Illumina sequencing. I would like to thank Winifred Okunlola for extracting and genotyping DNA from almost a third of the samples used in this study. I would like to thank Dr. Joe Galanko for completing the data analysis for this study. I would like to thank Félix Araújo-Pérez and the University of Maryland Institute for Genome Sciences for determining cytokine gene expression in this study. I would like to thank Dr. Blaire Steinwand for serving as my Biology Faculty Sponsor. I wish to acknowledge the Center of
References
American Cancer Society (2014). Colorectal Cancer Facts and Figures 2014-2016. Atlanta: American Cancer Society.
Caporaso J, J Kuczynski, et al. (2010). QIIME allows analysis of high-throughput community sequencing data. Nature Methods, 7(5): 335-336.
Chaudhuri A, N Wilson, et al. (2013) Host genetic background impacts modulation of the TLR4 pathway by RON in tissue-associated macrophages. Immunology and Cell Biology, 91(7): 451-460.
Ensinck M, M Valles, et al. (2013). Expression of the FUT2 gene and CD44 marker in patients with oral lesions. Immunología, 32(4): 123-128.
Fadrosh D, B Ma, et al. (2014). An improved dual-indexing approach for multiplexed 16S rRNA gene sequencing on the Illumina MiSeq platform. Microbiome, 2(1): 6.
Haghighi MM, M Vahedi, et al. (2009). Comparison of survival between patients with hereditary non polyposis colorectal cancer (HNPCC) and sporadic colorectal cancer. Asian Pacific Journal of Cancer Prevention, 10(3): 497-500.
Iyer S, G Cheng (2012). Role of Interleukin 10 Transcriptional Regulation in Inflammation and Autoimmune Disease. Critical Reviews in Immunology, 32(1): 23-63.
Kang M, P Edmundson, et al. (2013). Association of plasma endotoxin, inflammatory cytokines and risk of colorectal adenomas. BMC Cancer, 13: 91.
Lao V, W Grady (2011). Epigenetics and Colorectal Cancer. Nature Reviews Gastroenterology and Hepatology, 8(12): 686-700.
Livak K (1999). Allelic discrimination using fluorogenic probes and the 5’ nuclease assay. Genetic Analysis: Biomolecular Engineering, 14(5-6): 143-149.
Louis P, G Hold, H Flint (2014). The gut microbiota, bacterial metabolites and colorectal cancer. Nature Reviews Microbiology, 12(10): 661-672.
Mäkivuokko H, S Lahtinen, et al. (2012). Association between the ABO blood group and the human intestinal microbiota composition. BMC Microbiology, 12, 94.
McCoy A, F Araújo-Pérez, et al. (2013). Fusobacterium Is Associated with Colorectal Adenomas. PLoS One, 8, e53653.
Okunola W, A McCoy, et al. (2015). Association of FUT2 gene variant, gut bacteria and colorectal adenomas. Cancer Research, 75, 4586.
Roberts-Thomson IC, WJ Butler, et al. (1999). Meat, metabolic genotypes and risk for colorectal cancer. European Journal of Cancer Prevention, 8(3): 207-211.
Shen XJ, JF Rawls, et al. (2010). Molecular characterization of mucosal adherent bacteria and associations with colorectal adenomas. Gut Microbes,1(3):138-147.
Tong M, I McHardy, et al. (2014). Reprogramming of gut microbiome energy metabolism by the FUT2 Crohn’s disease risk polymorphism. International Society for Microbial Ecology Journal, 8, 2193-206.
Ürün Y, N Yildirim, et al. (2012). ABO and Rh blood groups and risk of colorectal adenocarcinoma. Asian Pacific Journal of Cancer Prevention, 13(12): 6097-6100.
Wacklin P, Mäkivuokko H, et al. (2011). Secretor phenotype (FUT2 gene) is strongly associated with the composition of bifidobacteria in the human intestine. PLOS ONE, 6, e20113.
Wang T, G Cai, et al. (2012). Structural segregation of gut microbiota between colorectal cancer patients and healthy volunteers. The International Society for Microbial Ecology Journal, 6, 320-329.
Weir T, D Manter, et al. (2013). Stool Microbiome and Metabolome Differences between Colorectal Cancer Patients and Healthy Adults. PLOS ONE, 8, e70803.
Wolpin BM, AT Chan, et al. (2009). ABO blood group and the risk of pancreatic cancer. Journal of the National Cancer Institute, 101(6): 424-431.
Wu H, V Tremaroli, et al. (2015). Linking Microbiota to Human Diseases: A Systems Biology Perspective. Trends in Endocrinology & Metabolism, 26(12): 758-770.