0095-1137/96/$04.00
1
0
Copyright
q
1996, American Society for Microbiology
Differentiation of Spotted Fever Group Rickettsiae by Sequencing and
Analysis of Restriction Fragment Length Polymorphism of
PCR-Amplified DNA of the Gene Encoding the Protein rOmpA
VERONIQUE ROUX, PIERRE-EDOUARD FOURNIER,
ANDDIDIER RAOULT*
Unite´ des Rickettsies, Faculte´ de Me´decine, Centre National de la Recherche Scientifique EPJ 0054,
13385 Marseille, France
Received 18 January 1996/Returned for modification 7 May 1996/Accepted 28 May 1996
Currently, the genotypic identification of the spotted fever group (SFG) rickettsiae is based on restriction
fragment length polymorphism analysis of PCR-amplified genes coding for the enzyme citrate synthase and the
surface proteins rOmpA and rOmpB. A set of useful restriction endonucleases was found following comparison
of Rickettsia rickettsii and R. prowazekii sequences. However, by using three PCR amplifications and four enzyme
digestions with this set, it was impossible to differentiate between all of the known serotypes of the SFG
rickettsiae. We amplified by PCR and sequenced using an automated laser fluorescent DNA sequencer a
fragment of the gene encoding the protein rOmpA from 21 serotypes of the SFG rickettsiae. A 632-bp
amplification product was obtained for most of the strains, although no product could be obtained by using R.
akari, R. australis, R. helvetica, and R. bellii DNAs. We found a characteristic sequence for all strains studied
except the two isolates of R. massiliae, isolates GS and Mtu1. Using the software package BISANCE, we
determined the restriction map of this fragment and identified five potentially useful endonucleases, RsaI, AluI,
PstI, XbaI, and AvaII. We confirmed the computer analysis-derived profiles by PCR-restriction fragment length
polymorphism analysis. The combination of the profiles obtained after digestion of the PCR product by RsaI
and PstI allowed for the differentiation of 16 strains. The use of AluI and XbaI allowed for the characterization
of R. parkeri and strain HA-91, respectively. R. africae and strain S were differentiated by AvaII digestion. Thus,
using a single PCR amplification, we were able to differentiate all of the SFG rickettsiae whose ompA gene was
amplified by PCR.
The bacteria included in the order Rickettsiales were first
described as short rods and gram-negative bacilli which
re-tained basic fuchsin when they were sre-tained by the method of
Gimenez (21) and which grew in association with eukaryotic
cells (42). Recently, Rickettsia tsutsugamushi, on the basis of
16S rRNA gene sequence analysis, has been excluded from the
genus Rickettsia (39), which presently comprises two groups,
the spotted fever group (SFG) and the typhus group. The
typhus group contains only three species, R. prowazekii, R.
typhi, and R. canada; however, the development of a new cell
microculture isolation technique (the shell vial technique) (28)
has recently led to an increase in the number of recognized
SFG rickettsiae, and at present there are 14 validated species
(R. conorii, R. rickettsii, R. rhipicephali, R. montana, R. japonica,
R. parkeri, R. africae, R. massiliae, R. sibirica, R. bellii, R. akari,
R. australis, R. helvetica, and R. felis [24, 38]), and 12 as yet
unrecognized species (“R. slovaca,” Thai tick typhus rickettsia,
Israeli tick typhus rickettsia, strain HA-91, Astrakhan fever
rickettsia, strain S, strain Bar29, strain MC16, AB bacterium
[43], Flinders Island rickettsia [7], and “R. amblyommii” [34])
have been described. The isolation of SFG rickettsiae from
their arthropod hosts, ticks, is difficult because of the presence
of other less fastidious bacteria associated with the arthropod
and especially because SFG rickettsiae are susceptible to most
antibiotics, which therefore cannot be included in any isolation
or enrichment cultures. The official criterion for the
identifi-cation of the SFG rickettsiae is mouse serotyping (33), and the
antigenic determinant of this serotyping is constituted by two
major envelope proteins of high molecular weight called
rOmpA and rOmpB. However, cross-reactivity between
differ-ent species exists, and moreover, this method requires antisera
and antigens of all of the described serotypes. PCR-based
methods for the identification of SFG rickettsiae are clearly
attractive because they not only circumvent the need for
cul-ture but they also possibly offer more sensitive and specific
alternatives. In 1991 Regnery et al. (35) proposed a method,
based on PCR-restriction fragment length polymorphism
(RFLP) analysis of the gene encoding the rOmpA protein,
which allowed for the differentiation of the nine SFG
rickett-siae studied. In 1994 Eremeeva et al. (19) used a combination
of this approach with a method based on PCR-RFLP analysis
of a fragment of the gene encoding the protein rOmpB to
identify all SFG strains except R. africae and R. parkeri.
A 632-bp amplification product of the ompA gene was
ob-tained for most of the strains and was sequenced. We
deter-mined the restriction map for each sequence and chose
endo-nucleases which allowed the identification of all of the strains
studied except R. akari, R. australis, R. helvetica, and R. bellii,
for which no PCR amplifications were obtained.
MATERIALS AND METHODS
Rickettsial strains.The strains studied are listed in Table 1.
(i) Recognized species.Four strains were obtained from the American Type Culture Collection (ATCC): R. conorii Moroccan (ATCC VR-141), R. conorii Seven (Malish; ATCC VR-613), R. rickettsii R (Bitteroot; ATCC VR-891), and
R. akari VR-148. R. rhipicephali, R. africae, and R. australis were supplied by
G. A. Dasch (Naval Medical Research Institute, Bethesda, Md.). Four strains were a gift from D. H. Walker (University of Texas, Galveston): R. montana, R.
* Corresponding author. Mailing address: Unite
´ des Rickettsies,
Faculte
´ de Me
´decine, Centre National de la Recherche Scientifique
EPJ 0054, 27 Blvd. Jean Moulin, 13385 Marseille, France. Phone: (33)
91-38-55-17. Fax: (33) 91-83-03-90. Electronic mail address: RAOULT
@CITI2.FR.
2058
on May 15, 2020 by guest
http://jcm.asm.org/
japonica, R. parkeri, and R. bellii. R. massiliae Mtu1 and GS were isolated in our
laboratory from ticks found in France and in the province of Fokis in central Greece, respectively. R. sibirica was obtained from the Gamaleya Research Institute Collection. R. helvetica was donated by W. Burgdorfer (Rocky Moun-tain Laboratory, Hamilton, Mont.).
(ii) Unrecognized species.Three strains were a gift from G. A. Dasch: “R.
slovaca,” Thai tick typhus rickettsia, and Israeli tick typhus rickettsia. Indian tick
typhus rickettsia, R. conorii M1, and strain S were obtained from the Gamaleya Research Institute Collection. Strain HA-91 and Astrakhan fever rickettsia were isolated in our laboratory from a tick collected from the Alashian region of the Inner Mongolia autonomous region and from a tick collected in the Astrakhan region of Russia, respectively. Strains Bar29 and MC16 were isolated in the laboratory from ticks collected in Spain and Morocco, respectively.
Rickettsial cultivation and nucleic acid preparation.The rickettsial strains were propagated on Vero cell monolayers (ATCC CRL 1587) in Eagle’s minimal essential medium (MEM; Seromed, Berlin, Germany) supplemented with 4% fetal calf serum (Seromed) and 2 mM glutamine at 328C. Harvesting was done when the degree of infection estimated by Gimenez (21) staining was high (3 to 5 days). Rickettsial cultures were centrifuged (12,0003g for 10 min),
resus-pended in MEM, and stored at2708C.
The bacterial walls of the rickettsiae were lysed during overnight incubation in a mixture of 1% sodium dodecyl sulfate (SDS) and proteinase K (0.2 mg/ml) in an Eppendorf tube at 378C. A phenol extraction and then a phenol-chloroform extraction were carried out after recovery of the aqueous phase. The DNA was precipitated by the addition of 2 volumes of chilled ethanol. The tube was stored overnight at2208C. The pellet of DNA obtained after centrifugation (12,0003
g for 10 min) was washed with cold 70% ethanol (12,0003g for 5 min). The
pellet was then dried and resuspended in sterile distilled water.
PCR amplification and sequencing method.We used the primers Rr 190.70p described by Regnery et al. (35) and primer 190-701, which was chosen from a
conserved region after alignment of the known sequences of R. rickettsii and R.
conorii Malish. The sequencing primers were the same as those used for the PCR
amplification and primers 190-267 and 190-488, which were chosen from con-served regions after alignment of the sequences determined for all strains stud-ied. The primers are described in Table 2.
For PCR amplification one primer was labelled at the 59end with biotin during oligonucleotide synthetis (Eurogentec, Seraing, Belgium), whereas the other was unmodified. Two microliters of the DNA preparation was amplified in a 100-ml reaction mixture containing 10 pmol of each primer; 200mM (each) dATP, dCTP, dGTP, and dTTP (Boehringer Mannheim Biochemicals, Indianapolis, Ind.); 1.25 U of AmpliTaq DNA polymerase (Perkin-Elmer Cetus, Norwalk, Conn.); and 6ml of a 25 mM solution of MgCl2(Perkin-Elmer) in 13Taq buffer
(Perkin-Elmer).
[image:2.612.59.560.81.467.2]Amplification was carried in a DNA thermal cycler (PTC 200; MJ Research Inc., San Francisco, Calif.) under the following conditions: an initial 3 min of denaturation at 958C was followed by 35 cycles of denaturation for 20 s at 958C,
TABLE 1. Rickettsial strains studied
Rickettsia Strain Source Geographical origin Human disease Reference(s)
R. conorii
Moroccan
Unknown
Morocco
Mediteranean spotted fever
12
R. conorii
Seven (Malish)
Unknown
South Africa
Mediteranean spotted fever
None
R. conorii
M1
Rhipicephalus sanguineus
Georgia, former USSR
23
R. conorii
Indian Tick Typhus
rickettsia
Rhipicephalus sanguineus
India
31
Astrakan fever
rickettsia
A-167
Rhipicephalus pumilio
Astrakhan region,
former USSR
Astrakhan fever
18, 40
Israeli tick
typhus
rickettsia
ISTT CDC1
Human
Israel
Israeli spotted fever
22
R. sibirica
246
Dermacentor nuttali
Former USSR
Siberian tick typhus
12
Strain HA-91
HA-91
Haemophysalis asiaticum
Inner Mongolia
45
R. parkeri
Maculatum 20
Amblyomma maculatum
Mississippi
11
“R. africae”
ESF-5
Amblyomma variegatum
Shulu Province,
Ethiopia
30
Strain S
S
Rhipicephalus sanguineus
Armenia, former
USSR
17
“R. slovaca”
13-B
Dermacentor marginalis
Slovakia
36
Thai tick
typhus
rickettsia
TT-118
Ixodes or Rhipicephalus
sp.
Thailand
37
R. rickettsii
R (Bitterroot)
Dermacentor andersoni
Montana
Rocky Mountain spotted
fever
11
R. japonica
YM
Human
Japan
Oriental spotted fever
41
R. massiliae
Mtu 1
Rhipicephalus turanicus
Camargue, France
9
R. massiliae
GS
Rhipicephalus sanguineus
Greece
6
Strain Bar 29
Bar 29
Rhipicephalus sanguineus
Spain
8
R. rhipicephali
3-7-6
Rhipicephalus sanguineus
Mississippi
15
R. montana
ATCC VR-611
Dermacentor variabilis or
Dermacentor andersoni
Ohio
10
Strain MC16
MC16
Hyaloma marginatum
Morocco
8
R. australis
Phillips
Human
Queensland, Australia
Queensland tick typhus
5
R. akari
MK (Kaplan)
Human
New York, N.Y.
Rickettsialpox
25
R. helvetica
C9P9
Ixodes ricinus
Switzerland
14
R. bellii
369L42-1
Dermacentor variabilis
Ohio
32
TABLE 2. Oligonucleotide primers
Primer Nucleotide sequence (59–39) ompA positions
Rr 190.70p
aATG-GCG-AAT-ATT-TCT-CCA-AAA
70–90
190-264
bCGT-TAT-CTC-ATT-CCA-ATT-AT
264–245
190-485
GCA-AAA-GCT-TAA-CTT-TAA-A
485–503
190-701
a,bGTT-CCG-TTA-ATG-GCA-GCA-TCT
701–681
aThe primer was used for PCR and the sequencing reaction. bThe primer was located on the complementary strand of DNA.
V
OL. 34, 1996
DIFFERENTIATION OF SPOTTED FEVER GROUP RICKETTSIAE
2059
on May 15, 2020 by guest
http://jcm.asm.org/
[image:2.612.316.556.651.710.2]annealing for 30 s at 468C, and extension for 1 min at 638C. The amplification was completed by holding for 7 min at 728C to allow complete extension of the PCR products. The success of the amplification was confirmed by resolution of the products by electrophoresis on a 1% agarose gel. The sizes of the PCR ampli-fication products were determined by comparison with a molecular weight stan-dard (marker VI; Boehringer) under UV light following ethidium bromide stain-ing.
The PCR products were purified by using streptavidin-coated magnetic beads (Dynabead M-280; Dynal Inc., Great Neck, N.Y.). Twenty microliters of beads was used for 40ml of amplified DNA product. The beads were washed three times in 13BW buffer (5 mM Tris-HCl [pH 7.5], 0.5 mM EDTA, 1 M NaCl) and were resuspended in 40ml of 23BW buffer. Then, the beads and the DNA were mixed in a 1.5-ml Eppendorf tube, and the mixture was incubated for 30 min with mixing at ambient temperature. The beads and bound DNA were sequestered with a Dynal MPC magnet, the supernatant was discarded, and the beads were washed twice with 13BW buffer. The bound DNA was denatured by adding 8
ml of 0.1 N NaOH. After 5 min of incubation at 378C the supernatant containing the complementary DNA strand was removed and neutralized with 4ml of 0.2 N HCl and 1ml of 1 M Tris-HCl (pH 7.4). The DNA strand that was left bound to the beads was washed once with 50ml of 0.1 N NaOH, once with 40ml of 13BW
buffer, and once with 50ml of water and was suspended in 10ml of distilled water. The Autoread sequencing kit (Pharmacia Biotech Norden, Sollentuna, Sweden) was used to carry out the sequencing reaction. Ten microliters of the bead suspension or the eluted DNA strand solution was used for each sequencing reaction, and the fluorescent primers were labeled at the 59end with fluorescein isothiocyanate (Eurogentec). The following reagents were mixed in a 1.5-ml Eppendorf tube: 10ml of template DNA, 2ml of the selected fluorescent primer (3 pmol/ml), and 2ml of annealing buffer. The tube was incubated at 688C for 10 min and was then left at ambient temperature for 10 min. One microliter of extension buffer, 3ml of dimethyl sulfoxide, and 2ml of T7 DNA polymerase (2.5 U/ml) were added to a tube that had been held at 378C, and immediately, 4.5ml of this mixture was pipetted into each of the four prewarmed sequencing mix-tures. Incubation was continued for 5 min, and 5ml of stop solution was added to each reaction mixture.
Sequencing reactions were resolved on 6% polyacrylamide gels (Ready Mix Gel; A.L.F. grade; Pharmacia) and electrophoresis was performed in the A.L.F. DNA Sequencer (Pharmacia) in 13TBE buffer (pH 8; 44.5 mM Tris-HCl, 44.5 mM boric acid, 1 mM EDTA).
Data analysis.The sequences of the gene coding for the rOmpA protein were aligned by using the multisequence alignment program CLUSTAL within the FIG. 1. Alignment of the nucleotide sequences of the rOmpA protein gene amplified by PCR (primers Rr 190.70p and 190-701). The endonuclease restriction sites are underlined (AluI, AGCT; AvaII, GGWCC; PstI, CTGCAG; RsaI, GTAC; and XbaI, TCTAGA). Homology is indicated by dots. Abbreviations: SEV, R. conorii Seven; MOR, R. conorii Moroccan; M1, R. conorii M1; IND, R. conorii Indian tick typhus rickettsia; AST, Astrakhan fever rickettsia; ISR, Israeli tick typhus rickettsia; SIB, R. sibirica; HA, strain HA-91; PAR, R. parkeri; S, strain S; AFR, R. africae; RR, R. rickettsii; THA, Thai tick typhus rickettsia; SLO, “R. slovaca”; JAP, R. japonica; RHI, R. rhipicephali; MC, strain MC16; MON, R. montana; MAS, R. massiliae; BAR, strain Bar29.
on May 15, 2020 by guest
http://jcm.asm.org/
BISANCE environment (16). The restriction maps of the determined sequences were obtained by using the same program. The percentages of similarity were determined by using the DNASIS software package (Hitachi Software Engineer-ing America, Ltd., Brisbane, Calif.).
PCR-RFLP of the rickettsia strains amplification product.Unmodified prim-ers Rr 190.70p and 190-701 were used to amplify DNA. Enzymatic digestion was performed by incubating 23.5ml of the amplified products with 2.8ml of enzyme buffer and 20 U of endonuclease. Digestion was continued for 3 h at 378C with
PstI, RsaI, and AluI, and the digested products were separated on 8%
polyacryl-amide gels (100 V for 4 h), which were stained with ethidium bromide and examined with a UV transilluminator. Digestion was continued overnight at 378C with XbaI and at 508C with AvaII, and the digested products were separated on a 1% agarose gel with the Mupid-2 minigel electrophoresis system (Eurogentec) for 30 min at 100 V. We used DNA molecular weight markers V and VI (Boehringer Mannheim).
Nucleotide sequence accession numbers.The nucleotide sequence data re-ported in this paper will appear in GenBank under the following accession numbers: U43795 for R. japonica, U43799 for R. massiliae, U43793 for strain GS, U43801 for R. montana, U43803 for R. rhipicephali, U43792 for strain Bar29, U43800 for strain MC16, U43804 for R. rickettsii, U43808 for “R. slovaca,” U43809 for Thai tick typhus rickettsia, U43807 for R. sibirica, U43796 for HA-91, U43802 for R. parkeri, U43790 for R. africae, U43805 for strain S, U43797 for Israeli tick typhus rickettsia, U43791 for Astrakhan fever rickettsia, U45244 for
R. conorii Moroccan, U43794 for R. conorii (Indian tick typhus rickettsia),
U43806 for R. conorii Malish, and U43798 for R. conorii M1.
RESULTS
PCR amplification.
For PCR amplification we used one of
the primers described by Regnery et al. (35), Rr 190.70p, and
a consensus primer from the R. conorii Seven and R. rickettsii
sequences which hybridized to the ompA gene close to the 5
9
end of the tandemly repeat units, primer 190-701. It was
pos-sible to obtain amplification with all of the strains studied
except R. bellii, R. akari, R. australis, and R. helvetica even if the
annealing temperature was reduced to below 46
8
C. The length
of the fragments obtained for most of the studied strains was
632 bp. For three strains the product was smaller because of
deletions of 156, 24, and 3 bp for R. conorii M1, R. conorii
Moroccan, and R. montana, respectively. For R. conorii Indian
tick typhus rickettsia, an insertion of 3 bp was noted, and so the
fragment was 635 bp (Fig. 1).
Sequence analysis.
For all PCR products both DNA strands
were sequenced twice. A specific sequence was obtained for all
of the rickettsial strains tested except the two isolates of the
species R. massiliae, isolates GS and Mtu1. The strains shared
between 99.3 and 67.1% sequence similarity. Following
multi-ple alignment of the sequences, several insertions and
dele-tions were noted. For example, the insertion of the codon TTT
(nucleotides 191 to 193) was found in only three strains of R.
conorii (strains Seven, Moroccan, and M1). The deletion of
codon AAT (nucleotides 250 to 252) was found in the four
strains of R. conorii (strains Seven, Moroccan, and M1 and the
Indian tick typhus rickettsia). We also noted mutations
char-acteristic of a group of rickettsial strains, for example, the
transition T/C in R. rhipicephali, strain MC16, R. montana, R.
massiliae, and strain Bar29 (nucleotide 340) or the transition
C/T in R. sibirica, strain HA-91, R. parkeri, strain S, and R.
africae (nucleotide 435).
The sequence of R. rickettsii was identical to the one
pub-lished by Anderson et al. (3). Recently, the sequence of the
PCR product amplified from R. japonica with the primer pair
Rr 190.70p and Rr 190.602n was published (44). It was
iden-tical to the one that we determined.
[image:4.612.59.296.52.649.2]Restriction maps.
Using a software package of the French
service BISANCE, the suitabilities of a large number of
endo-nucleases could be assessed. After preliminary analysis of the
data we selected five enzymes which allowed for the
identifi-cation of all of the species studied: PstI, RsaI, AluI, XbaI, and
AvaII. For PstI we obtained a specific profile with seven strains
(Moroccan, M1, Indian tick typhus rickettsia, Israeli tick
ty-phus rickettsia, “R. slovaca,” R. massiliae, and R. montana)
(Fig. 2). We obtained the same profiles for Astrakhan fever
rickettsia, R. sibirica, HA-91, R. parkeri, R. africae, and strain S;
seven R. rickettsii and Thai tick typhus rickettsia; R. japonica
and MC16; and Bar29 and R. rhipicephali. For RsaI we noted
a specific profile with eight strains (Moroccan, M1, Indian tick
FIG. 1—Continued.
V
OL. 34, 1996
DIFFERENTIATION OF SPOTTED FEVER GROUP RICKETTSIAE
2061
on May 15, 2020 by guest
http://jcm.asm.org/
typhus rickettsia, Thai tick typhus rickettsia, R. japonica, R.
rhipicephali, MC16, and R. montana) (Fig. 3). We obtained the
same profiles for R. conorii Seven, Astrakhan fever rickettsia,
and Israeli tick typhus rickettsia; HA-91, R. parkeri, R. africae,
and strain S; R. sibirica, R. rickettsii and R. slovaca; and R.
massiliae and Bar29. For AluI we obtained a specific profile for
eight strains (Moroccan, M1, Indian tick typhus rickettsia,
As-trakhan fever rickettsia, R. sibirica, R. parkeri, R. rickettsii, and
R. montana) (Fig. 4). We obtained the same profile for R.
conorii Seven, Israeli tick typhus rickettsia strain HA-91, R.
africae, strain S, Thai tick typhus rickettsia, and “R. slovaca”;
and R. japonica, R. massiliae, Bar29, R. rhipicephali, and MC16.
If we combined the profiles obtained with these three
endo-nucleases it was possible to identify all strains except HA-91, R.
africae, and strain S. The characterization of HA-91 was
ob-tained with the XbaI profile, and the differentiation of R.
afri-cae and strain S was obtained by comparison of the AvaII
profiles (Fig. 5).
PCR-RFLP analysis of the amplification products of the
rickettsial strains.
We tested the five endonucleases chosen
and found the profiles hypothesized by analysis of the obtained
sequences with the software package of the French service
BISANCE (Fig. 6 and 7). For XbaI digestion, it was impossible
to obtain total digestion with the PCR-amplified product from
strain HA-91, but the digested fragments obtained were those
expected.
DISCUSSION
SFG rickettsiae are obligate intracellular (sometimes
in-tranuclear) bacteria which are transmitted via arthropod
vec-tors, in whom they are maintained by transovarial transmission
(13). They have a worldwide distribution that is determined by
the geographic distribution of infected ticks and mites. The
introduction and the development of improved culture and
identification methods have allowed for the characterization of
more and more isolates of SFG rickettsiae over the past few
years, and this in turn has led to the identification of new
species and the reevaluation of the distribution of the
recog-nized species. Some of the SFG rickettsiae are pathogenic for
humans and are responsible for tick-borne spotted fevers in
which the principal symptoms consist of fever, headache, and
eruption (R. conorii, R. rickettsii, R. akari, R. australis, R.
sibirica, R. japonica, Israli tick typhus rickettsia, Astrakhan
fever rickettsia, R. africae. Flinders Island spotted fever isolate,
and R. felis). Although other strains have only been isolated
from ticks, it is unwise to consider them nonpathogenic for
humans; R. africae was isolated from a patient in Zimbabwe
(26, 27) some 40 years after the characterization of the same
serotype in one tick in Ethiopia (30).
[image:5.612.61.552.65.358.2]Although PCR with specific primers is recognized as a
sen-sitive method for the detection of bacterial DNA and
PCR-RFLP analysis has proven to be a rapid method for their
identification, few SFG rickettsiae genes have been sequenced
to date. Those which have been sequenced include the genes
encoding two surface proteins, rOmpA and rOmpB. Their
significance is evidenced by their surface location (1),
immu-nogenicity for humans and mice (1, 20), and the fact that the
rOmpA protein functions as a protective immunogen in guinea
pigs (29). The rOmpA protein seems to be specific to the SFG
rickettsiae. It has been characterized by SDS-polyacrylamide
gel electrophoresis (PAGE), Western blotting
(immunoblot-ting) (9, 20), and PCR-RFLP analysis in most of these bacteria
(19, 35). Because both the gene and its product demonstrate
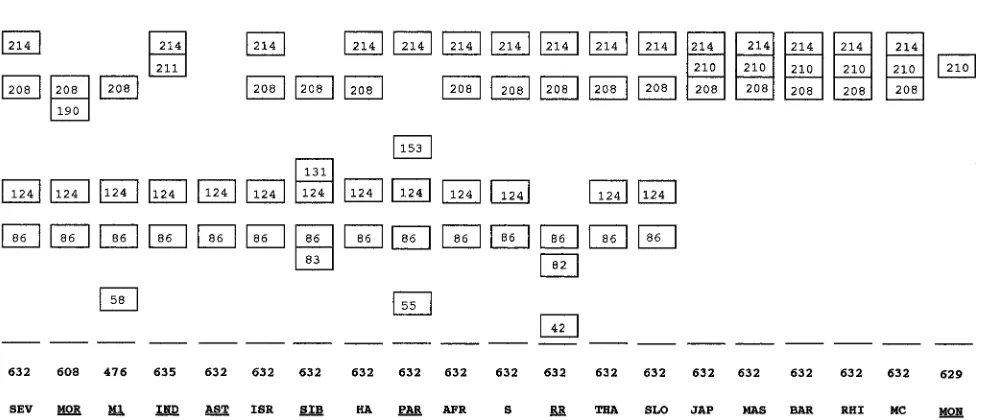
FIG. 2. Schematic representation of the profiles obtained after enzymatic digestion with PstI of PCR-amplified DNA derived from the ompA genes of 20 strains of the SFG rickettsiae. The profiles were deduced from the sequences that were determined. Fragment sizes are in base pair (boxed), and total fragment sizes are indicated. The strains with specific profiles are underlined. See Fig. 1 legend for definitions of strain abbreviations.
on May 15, 2020 by guest
http://jcm.asm.org/
marked diversity within the SFG rickettsiae (contrary to the
gene coding for a protein of 17 kDa [4] and the gene encoding
citrate synthase [35]), it is potentially a good tool for the
differentiation of the SFG rickettsiae. The first methods used
for identification were serotyping by
microimmunofluores-cence (33) and SDS-PAGE (2). However, because both
meth-ods require culture of all of the recognized serotypes and
microimmunofluorescence requires the production of
[image:6.612.62.556.81.359.2]antibod-FIG. 3. Schematic representation of the profiles obtained after enzymatic digestion with RsaI of PCR-amplified DNA derived from the ompA genes of 20 strains of the SFG rickettsiae. See Fig. 1 and 2 legends for definitions of abbreviations and symbols, respectively.
FIG. 4. Schematic representation of the profiles obtained after enzymatic digestion with AluI of PCR-amplified DNA derived from the ompA genes of 20 strains of the SFG rickettsiae. See Fig. 1 and 2 legends for definitions of abbreviations and symbols, respectively.
V
OL. 34, 1996
DIFFERENTIATION OF SPOTTED FEVER GROUP RICKETTSIAE
2063
on May 15, 2020 by guest
http://jcm.asm.org/
[image:6.612.60.552.496.706.2]ies in laboratory animals, both techniques are long and
mate-rial-consuming and are sometimes hazardous. In 1991 Regnery
et al. (35) introduced an identification method for the SFG
rickettsiae based on PCR-RFLP of a fragment of the gene
encoding the rOmpA protein. The nine different strains
in-cluded in the present study could be differentiated from one
another by using a combination of PstI and RsaI. Interestingly,
two slightly different profiles were obtained for the Seven and
Moroccan strains of the species R. conorii. In 1994 Eremeeva
et al. (19) expanded this approach to the study of 36 strains of
the SFG rickettsiae. It was possible to differentiate all of these
strains except R. africae and R. parkeri by using a combination
of three amplifications and four enzyme digestions, and a
fur-ther specific profile was found for anofur-ther isolate of R. conorii,
isolate M1. In both of the studies enzymes were chosen
em-pirically following comparison of only the R. rickettsii and R.
prowazekii rOpmA gene sequences. Subsequently, new isolates
which also cannot be characterized by these methods have
been described: strain HA-91 (45), strain S, (17), and
Astra-khan fever rickettsia (18).
[image:7.612.92.273.72.277.2]In the work described here we sequenced an amplified
frag-ment of the ompA gene from SFG rickettsiae in order to
provide a rationale for their identification by PCR-RFLP
anal-ysis. We sequenced a fragment of the gene slightly longer than
the one amplified by Regnery et al. (35) from all of the
de-scribed strains of the SFG rickettsiae except “R. amblyommii,”
Flinders Island spotted fever isolate, R. felis, and the AB
bac-terium, which were not available in our laboratory. Our
objec-tive was to determine enzymes which would allow for the
identification of all of the strains of SFG rickettsiae studied
and to obtain an exact size of the different fragments generated
after endonuclease restriction. Our results were in
concor-dance with those of Regnery et al. (35) and Eremeeva et al.
(19). PCR-amplified fragments of 632 bp were obtained from
all strains except for R. conorii M1, R. conorii Moroccan,
In-dian tick typhus rickettsia, and R. montana. These strains
pos-sessed either insertions or deletions of one or several codons.
We used primer 190-701 instead of primer Rr 190.602n
be-cause amplification yielded a fragment which after digestion
with PstI allowed for the differentiation of Astrakhan fever
rickettsia and Israeli tick typhus rickettsia and which after
digestion with RsaI allowed for the differentiation of MC16
and R. rhipicephali. Three endonucleases were found to be
particularly useful; RsaI, AluI, and PstI allowed for the
char-acterization of eight, eight, and seven strains, respectively. The
combination of the profiles obtained with two of these three
enzymes allowed for the identification of all of the strains
studied with the exception of strain HA-91, R. africae, and
strain S. HA-91 did, however, possess a specific site for XbaI at
position 411, and strain S possessed a site for AvaII at position
301 which was present in the other strains but which was absent
from R. africae. Thus, by using the combined profiles obtained
with the five endonucleases, it was possible to differentiate
between all the strains which yielded a PCR product. It was
interesting that we obtained amplification of different sizes for
four isolates of the species R. conorii (M1, Moroccan, Indian
tick typhus rickettsia, and Seven) and, therefore, different
pro-files after enzymatic digestion. We tested six other strains of R.
conorii, five strains from humans isolated in our laboratory and
one strain isolated from a tick (Kenya tick typhus rickettsia
[12]), and found the same sequence as that obtained for R.
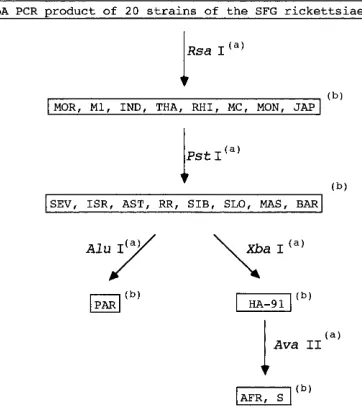
[image:7.612.60.296.510.648.2]FIG. 5. Proposed algorithm for the identification of the SFG rickettsiae by PCR-RFLP analysis of a fragment of the gene encoding the rOmpA protein. (a) Endonucleases used to obtain identification of the strains by combination of the profiles obtained. (b) Strains characterized after combined PCR-RFLP analysis with the endonucleases described. The definitions of the strain abbreviations are given in the Fig. 1 legend.
FIG. 6. Ethidium bromide-stained polyacrylamide gel of an AluI restriction endonuclease digest of DNA amplified by using the Rr 190.70p and 190-701 primer pair. Lane 1, R. conorii Seven; lane 2, R. conorii Moroccan; lane 3: R.
conorii M1; lane 4, R. conorii Indian tick typhus rickettsia; lane 5, Astrakhan
fever rickettsia; lane 6, Israeli tick typhus rickettsia; lane 7, R. sibirica; lane 8, strain HA-91; lane 9, R. parkeri; lane 10, R. africae; lane 11, strain S; lane 12, R.
rickettsii; lane 13, Thai tick typhus rickettsia, lane 14, “R. slovaca”; lane 15, R. japonica; lane 16, R. rhipicephali; lanes S, standard DNA size marker V (sizes are
in base pairs).
FIG. 7. Negative image of an ethidium bromide-stained agarose gel of the PCR-RFLP patterns of Rickettsia species. DNAs were amplified by using Rr 190.70p and 190-701 primers and were digested with XbaI (lane 1, R. africae; lane 2, R. parkeri; lane 3, strain S; lane 4, strain HA-91) and AvaII (lane 5, R. parkeri; lane 6, strain S; lane 7, R. africae; lane 8, strain HA-91). Lanes S, standard DNA size marker VI (sizes are in base pairs).
on May 15, 2020 by guest
http://jcm.asm.org/
conorii Seven, so it seems that this strain is typical of the
species R. conorii.
ACKNOWLEDGMENT
We are grateful to Richard Birtles for reviewing the manuscript.
REFERENCES
1. Anacker, R. L., R. H. List, R. E. Mann, S. F. Hayes, and L. A. Thomas. 1985. Characterization of monoclonal antibodies protecting mice against Rickettsia
rickettsii. J. Infect. Dis. 151:1052–1060.
2. Anacker, R. L., R. E. Mann, and C. Gonzales. 1987. Reactivity of monoclonal antibodies of Rickettsia rickettsii with the spotted fever and typhus group rickettsiae. J. Clin. Microbiol. 25:167–171.
3. Anderson, B. E., G. A. McDonald, D. C. Jones, and R. L. Regnery. 1990. A protective protein antigen of Rickettsia rickettsii has tandemly repeated, near-identical sequences. Infect. Immun. 58:2760–2769.
4. Anderson, B. E., and T. Tzianabos. 1989. Comparative sequence analysis of a genus-common rickettsial antigen gene. J. Bacteriol. 171:5199–5201. 5. Andrew, R., J. M. Bonnin, and S. Williams. 1946. Tick typhus in North
Queensland. Med. J. Aust. 2:253.
6. Babalis, T., Y. Tselentis, V. Roux, A. Psaaroulaki, and D. Raoult. 1994. Isolation and identification of a rickettsial strain related to Rickettsia
mas-siliae in greek ticks. Am. J. Trop. Med. Hyg. 50:365–372.
7. Baird, R. W., M. Llyod, J. Stenos, B. C. Ross, R. S. Stewart, and B. Dwyer. 1992. Characterization and comparison of Australian human spotted fever group rickettsiae. J. Clin. Microbiol. 30:2896–2902.
8. Beati, L. Unpublished data.
9. Beati, L., J.-P. Finidori, B. Gilot, and D. Raoult. 1992. Comparison of serologic typing, sodium dodecyl sulfate-polyacrylamide gel electrophoresis protein analysis, and genetic restriction fragment length polymorphism anal-ysis for identification of rickettsiae: characterization of two new rickettsial strains. J. Clin. Microbiol. 30:1922–1930.
10. Bell, E. J., G. M. Kohls, H. G. Stoenner, and D. B. Lackman. 1963. Non pathogenic rickettsias related to the spotted fever group isolated from ticks,
Dermacentor variabilis and Dermacentor andersoni from eastern Montana.
J. Immunol. 90:770–781.
11. Bell, E. J., and E. G. Pickens. 1953. A toxic substance associated with the rickettsias of the spotted fever group. J. Immunol. 70:461.
12. Bell, E. J., and H. G. Stoenner. 1960. Immunologic relationships among the spotted fever group of rickettsias determined by toxin neutralisation tests in mice with convalescent animal serums. J. Immunol. 84:171–210.
13. Burgdorfer, W. 1988. Ecological and epidemiological considerations of Rocky Mountain spotted fever and scrub typhus, p. 33–50. In D. H. Walker (ed.), Biology of rickettsial diseases, vol. I. CRC Press, Inc., Boca Raton, Fla. 14. Burgdorfer, W., A. Aeschlimann, O. Peter, S. F. Hayes, and R. N. Philip. 1979. Ixodes ricinus: vector of a hitherto undescribed spotted fever group agent in Switzerland. Acta Trop. 39:357–367.
15. Burgdorfer, W., L. P. Brinton, W. L. Krinsky, and R. N. Philip. 1978.
Rickettsia rhipicephali: a new spotted fever group rickettsia from the brown
dog tick Rhipicephalus sanguineus, p. 307–316. In J. Kazar, R. A. Ormsbee, and I. N. Tarasevich (ed.), Rickettsiae and rickettsial diseases. Publishing House of the Slovak Academy of Sciences, Bratislava, Slovakia.
16. Dessen, P., C. Fondrat, C. Valencien, and G. Munier. 1990. BISANCE: a French service for access to biomolecular sequences databases. Comput. Appl. Biosci. CABIOS 6:355–356.
17. Eremeeva, M. E., N. Balayeva, V. Roux, V. Ignatovich, M. Kotsinjan, and D.
Raoult.1995. Genomic and proteinic characterization of strain S, a rickettsia isolated from Rhipicephalus sanguineus ticks in Armenia. J. Clin. Microbiol.
33:2738–2744.
18. Eremeeva, M. E., L. Beati, V. A. Makarova, N. F. Fetisova, I. V. Tarasevich,
N. M. Balayeva, and D. Raoult.1994. Astrakhan fever rickettsiae: antigenic and genotypic analysis of isolates obtained from human and Rhipicephalus
pumilio ticks. Am. J. Trop. Med. Hyg. 51:697–706.
19. Eremeeva, M., X. Yu, and D. Raoult. 1994. Differentiation among spotted fever group rickettsiae species by analysis of restriction fragment length polymorphism of PCR-amplified DNA. J. Clin. Microbiol. 32:803–810. 20. Gilmore, R. D., Jr., and T. Hackstadt. 1991. DNA polymorphism in the
conserved 190 kDa antigen gene repeat region among spotted fever group rickettsiae. Biochim. Biophys. Acta 1097:77–80.
21. Gimenez, D. F. 1964. Staining Rickettsiae in yolk-sac cultures. Stain Technol.
39:135–140.
22. Goldwasser, R. A., Y. Steiman, W. Klingberg, T. A. Swartz, and M. A.
Klingberg.1974. The isolation of strains of rickettsiae of the spotted fever
group in Israel and their differentiation from other members of the group by immunofluorescence methods. Scand. J. Infect. Dis. 6:53–62.
23. Golinevitch, H. 1960. A propos de la diffe´renciation de quelques rickettsies du groupe de la fie`vre pourpre´e a` tiques. Arch. Inst. Pasteur Tunis 37:13–22. 24. Higgins, J. A., S. Radulovic, M. E. Schrieffer, and A. F. Azad. 1996. Rickettsia
felis: a new species of pathogenic rickettsia isolated from cat fleas. J. Clin.
Microbiol. 34:671–674.
25. Huebner, R. J., and C. Armstrong. 1946. Rickettsialpox—a newly recognized rickettsial disease. I. Isolation of the ethiological agent. Public Health Rep.
61:1605.
26. Kelly, P., L. Beati, P. R. Mason, L. A. Matthewman, V. Roux, and D. Raoult. 1996. Rickettsia africae sp. nov., the ethiological agent of African tick bite fever. Int. J. Syst. Bacteriol. 46:611–614.
27. Kelly, P., L. Matthewman, L. Beati, D. Raoult, P. Mason, M. Deary, and R.
Makombe.1992. African tick-bite fever: a new spotted fever group rickett-siosis under an old name. Lancet 340:382–383.
28. Marrero, M., and D. Raoult. 1989. Centrifugation shell vial technique for rapid detection of Mediterranean spotted fever in blood culture. Am. J. Trop. Med. Hyg. 40:197–199.
29. McDonald, G. A., R. L. Anacker, R. E. Mann, and L. J. Milch. 1988. Pro-tection of guinea pigs from experimental Rocky Mountain spotted fever with a cloned antigen of Rickettsia rickettsii. J. Infect. Dis. 158:228–231. 30. Philip, C. B., H. Hoogstraal, R. Reiss-Gutfreund, and C. M. Cliford. 1966.
Evidence of rickettsial disease agents in ticks from Ethiopian cattle. Bull W. H. O. 35:127–131.
31. Philip, C. B., L. E. Hughes, K. N. A. Rao, and S. L. Kaira. 1950. Studies of Indian tick typhus and its relation to other human typhus-like rickettsiosis, p. 115. In 5th International Congress for Microbiology. Rio de Janeiro. 32. Philip, R. N., E. A. Casper, R. L. Anacker, J. Cory, S. F. Hayes, W.
Burg-dorfer, and C. E. Yunker.1983. Rickettsia bellii sp. nov.: tick-borne rickettsia, widely distributed in the United States, that is distinct from the spotted fever and typhus biogroups. Int. J. Syst. Bacteriol. 33:94–106.
33. Philip, R. N., E. A. Casper, W. Burgdorfer, R. K. Gerloff, L. E. Hughes, and
E. J. Bell.1978. Serologic typing of Rickettsiae of spotted fever group by microimmunofluorescence. J. Immunol. 121:1961–1968.
34. Pretzman, C., D. R. Stothard, D. Ralph, and A. Fuerst. 1994. A new
Rick-ettsia, isolated from the lone star tick, Amblyomma americanum (Ixodidae), p.
24. In Abstracts of the 11th Sesquiannual Meeting of the American Society for Rickettsiology and Rickettsial Diseases. American Society for Rickettsi-ology and Rickettsial Diseases. St. Sumons Island, Ga.
35. Regnery, R. L., C. L. Spruill, and B. D. Plikaytis. 1991. Genotypic identifi-cation of rickettsiae and estimation of intraspecies sequence divergence for portions of two rickettsial genes. J. Bacteriol. 173:1576–1589.
36. Rehacek, J. 1984. Rickettsia slovaca, the organism and its ecology. Acta Sci. Nat. Acad. Sci. Bohemoslov. Brno 18:1–50.
37. Robertson, R. G., and C. L. Wisseman, Jr. 1972. Tick-borne rickettsiae of the spotted fever group in West Pakistan. II. Serological classification of isolates from West Pakistan and Thailand: evidence for two new species. Am. J. Epidemiol. 97:55–64.
38. Schriefer, M. E., J. B. Sacci, Jr., S. Dumler, M. G. Bullen, and A. F. Azad. 1994. Identification of a novel rickettsial infection in a patient diagnosed with murine typhus. J. Clin. Microbiol. 32:949–954.
39. Tamura, A., N. Ohashi, H. Urakami, and S. Miyamura. 1995. Classification of Rickettsia tsutsugamushi in a new genus, Orientia gen. nov., as Orientia
tsutsugamushi comb. nov. Int. J. Syst. Bacteriol. 45:589–591.
40. Tarasevich, I. V., V. A. Makarova, N. F. Fetisova, A. V. Steoanov, E. D.
Miskarova, N. Balayeva, and D. Raoult.1991. Astrakhan fever, a spotted-fever rickettsiosis. Lancet 337:172–173.
41. Uchida, T., T. Uchiyama, K. Kumano, and D. H. Walker. 1992. Rickettsia
japonica sp. nov., the etiological agent of spotted fever group rickettsiosis in
Japan. Int. J. Syst. Bacteriol. 42:303–305.
42. Weiss, E., and J. W. Moulder. 1984. Order I. Rickettsiales Gieszczkiewicz 1939, 25AL, p. 687–701. In N. R. Kreig and J. G. Holt (ed.), Bergey’s manual
of systematic bacteriology, vol. 1. The Williams & Wilkins Co., Baltimore. 43. Werren, J. H., G. D. D. Hurst, W. Zhang, J. A. J. Breeuvwer, R. Stouthamer,
and M. E. N. Majerus.1994. Rickettsial relative associated with male killing in the ladybird beetle (Adalia bipunctata). J. Bacteriol. 176:388–394. 44. Yan, Y., T. Uchiyama, and T. Uchida. 1995. Nucleotide sequence of
poly-merase chain reaction product amplified from Rickettsia japonica DNA using
Rickettsia rickettsii 190-kilodalton surface antigen gene primers. Microbiol.
Immunol. 38:865–869.
45. Yu, X., Y. Jin, M. Fan, G. Xu, Q. Liu, and D. Raoult. 1993. Genotypic and antigenic identification of two new strains of spotted fever group rickettsiae isolated from China. J. Clin. Microbiol. 31:83–88.