Capsular Polysaccharide Synthesis Gene Clusters of
Streptococcus suis
Masatoshi Okura,aClaude Lachance,bMakoto Osaki,aTsutomu Sekizaki,cFumito Maruyama,d,eTakashi Nozawa,dIchiro Nakagawa,d
Shigeyuki Hamada,f,gCéline Rossignol,hMarcelo Gottschalk,bDaisuke Takamatsua,i
Bacterial and Parasitic Diseases Research Division, National Institute of Animal Health, National Agriculture and Food Research Organization, Tsukuba, Japana ; Faculty of Veterinary Medicine, University of Montreal, St-Hyacinthe, Quebec, Canadab
; Research Center for Food Safety, Graduate School of Agricultural and Life Sciences, The University of Tokyo, Tokyo, Japanc
; Section of Bacterial Pathogenesis, Tokyo Medical and Dental University, Graduate School of Medical and Dental Sciences, Tokyo, Japand
; Section of Microbial Genomics and Ecology, Tokyo Medical and Dental University, Graduate School of Medical and Dental Sciences, Tokyo, Japane ; Research Institute for Microbial Diseases, Osaka University, Osaka, Japanf
; Thailand-Japan Research Collaboration Center on Emerging and Reemerging Infections (RCC-ERI), Nonthaburi, Thailandg
; École Nationale Vétérinaire de Toulouse, Toulouse, Franceh
; The United Graduate School of Veterinary Sciences, Gifu University, Gifu, Japani
We developed a practical and easy two-step multiplex PCR assay to aid in serotyping ofStreptococcus suis. The assay accurately
typed almost all of the serotype reference strains and field isolates of various serotypes and also identified the genotypes of cap-sular polysaccharide synthesis gene clusters of some serologically nontypeable strains.
S
treptococcus suisis an important zoonotic pathogen that causes meningitis, septicemia, endocarditis, and other diseases in pigs and humans.S. suisstrains have been classified into 35 serotypes (serotypes 1 to 34 and 1/2, which reacts with both serotype 1 and 2 typing sera) (1–4) on the basis of antigenic differences in their capsular polysaccharide (CP) (5). Serotyping ofS. suisis one of the most useful methods to understand the epidemiology of a partic-ular outbreak and monitor the prevalence of potentially hazard-ous strains. However, serotyping with all 35 typing antisera is time-consuming, and preparing the antisera is not easy due to the high cost and labor associated with its production. Additionally, cross-reactions in coagglutination tests with typing antisera and the presence of autoagglutinating strains increase the difficulty of serotyping in some cases. Therefore, the development of more practical and easier serotyping methods is desired.CP synthesis (cps) genes are clustered on a single locus of the chromosome inS. suis(6,7). We recently sequenced and analyzed thecpsgene clusters of all 35 serotype reference strains (8) and reported that 31 (serotypes 3 to 13 and 15 to 34) possessed sero-type-specific genes, while thecpsgene clusters of serotypes 1 and 14 and serotypes 2 and 1/2 were almost identical in each pair (8). Liu et al. (9) recently developed multiplex PCR assays to target serotype-specificcpsgenes for the molecular serotyping ofS. suis. In their method, the reference strains of 33 serotypes (serotypes 1 to 31, 33, and 1/2) were sorted into their respective serotypes by three multiplex PCR assays, although serotypes 1 and 1/2 were not distinguished from serotypes 14 and 2, respectively (9). In addi-tion, 84 isolates from a patient and clinically healthy pigs, which covered 20 serotypes, were correctly assigned serotypes predicted by coagglutination tests, except for those of a new serotype (sero-type 21/29) (9). However, these methods have not yet been vali-dated using field isolates from diseased pigs as well as nontypeable ones by the coagglutination test. Moreover, serotypes 32 and 34 were not included as targets for typing, because the reference strains of these serotypes have been reclassified asStreptococcus orisratti(10). In addition to the serotype 32 and 34 reference strains, those of serotypes 20, 22, 26, and 33 were also recently suggested to be removed from the taxon ofS. suis(11). However, whether all isolates of these serotypes actually belong to a species that is different fromS. suisremains unclear. From a diagnostic
point of view, the isolates of these serotypes recovered from dis-eased pigs are still identified asS. suis(12).
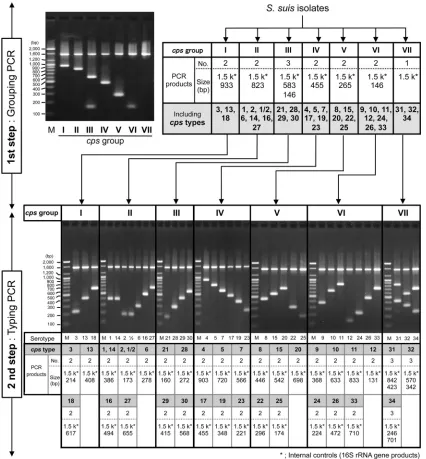
In the present study, we developed a novel method to type the 35cpsgene clusters ofS. suisand validated its usefulness with 483 isolates from diseased and healthy animals and human patients, which covered 30 serotypes. With our methods (cpstyping), we determinedcpstypes, which were numbered corresponding to the serotypes, using only two multiplex PCRs (Fig. 1). The first PCR (grouping PCR) detectedcpsgenes conserved in multiple sero-types and classified the tested strains into sevencpsgroups. The second PCR (typing PCR) detectedcpsgenes specific to the respec-tive serotypes of each group and identified thecpstype of the strain.
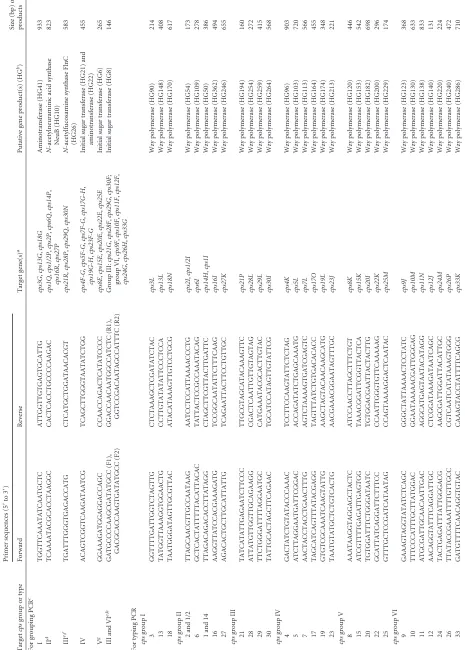
The targetcpsgenes, primer sequences, and predicted product size(s) of each PCR are shown inTable 1. Target genes were se-lected on the basis of the clustering analysis data of allcpsgene products reported previously (8). Universal primers for the am-plification of 16S rRNA genes (13) were used as internal controls for all reactions. The genomic DNA of strains/isolates was ex-tracted as described previously (for determination of the sensitiv-ity of PCR assays) (8) or using InstaGene Matrix (Bio-Rad, Her-cules, CA) following the manufacturer’s instructions. All PCR mixtures contained 1⫻ Qiagen multiplex master mix (Qiagen, Hilden, Germany) and 0.2M each primer. The PCR conditions were as follows: an initial activation at 95°C for 15 min followed by 30 cycles of denaturation at 94°C for 30 s, primer annealing at 60°C (for grouping PCR) or 58°C (for typing PCR) for 90 s, and extension at 72°C for 90 s, and then a final extension at 72°C for 10 min. The 483 putativeS. suisisolates used to validate the devel-oped assays were isolated from diseased pigs (394 isolates),
clini-Received6 December 2013 Returned for modification3 January 2014
Accepted18 February 2014
Published ahead of print26 February 2014
Editor:E. Munson
Address correspondence to Daisuke Takamatsu, [email protected]. Copyright © 2014, American Society for Microbiology. All Rights Reserved. doi:10.1128/JCM.03411-13
on May 16, 2020 by guest
http://jcm.asm.org/
cally healthy pigs (60 isolates), diseased cows (2 isolates), healthy cows (17 isolates), a diseased sheep (1 isolate), and patients (9 isolates). All isolates tested were positive for thegdh-PCR assay (data not shown), which specifically detects the glutamate dehy-drogenase genes ofS. suisstrains, including the reference strains of serotypes 20, 22, 26, and 32 to 34 (14). All isolates were also sero-typed by coagglutination tests in the reference laboratory (Univer-sity of Montreal) as previously described (15).
Ourcpstyping assigned all serotype reference strains, including those of serotypes 32 and 34, to the expectedcpstypes. However, serotypes 1 and 1/2 could not be distinguished from serotypes 14 and 2, respectively, which is consistent with previous observations with the reported molecular serotyping method (9) (Fig. 1). Al-though the developed methods accurately assigned nearly all field isolates (341/357; 95.5%) to thecpstypes predicted from their serotypes, 16 of them were not typed bycpstyping (Table 2). These
FIG 1Scheme ofcpstyping using a two-step multiplex PCR assay and the PCR products amplified from 35S. suisserotype reference strains. The first PCR
(grouping PCR) classified the tested strains into 7 groups (cpsgroups I to VII). Numbers and sizes of grouping PCR products amplified from strains of eachcps
group and the electrophoresis image are shown in the upper panel. Approximately 1.5-kbp bands that appeared in all grouping PCR assays are the internal control products (16S rRNA gene products). Only the internal control products were amplified fromcpsgroup VII strains. The strains classified by grouping PCR were then subjected to the second PCR (typing PCR) using primers specific to the respectivecpsgroups in order to identify thecpstype of each strain. Number and size of typing PCR products amplified from allS. suisserotype reference strains and the electrophoresis images are shown in the bottom panels. Approximately 1.5-kbp bands that appeared in all typing PCR assays are the internal control products (16S rRNA gene products). The serotypes of reference strains are indicated below the lanes. All PCR products were electrophoresed on a 2% agarose gel (100 V, 40 min), stained with ethidium bromide, and photographed under UV light. Lanes M show DNA molecular weight markers (in thousands [k]) (100-bp DNA ladder; Bioneer, Daejeon, South Korea).
on May 16, 2020 by guest
http://jcm.asm.org/
[image:2.585.81.507.67.526.2]TABLE 1 Primer sequences used in this study and the predicted sizes of the PCR products Target cps group or type Primer sequences (5 = to 3 = ) Target gene(s) a Putative gene product(s) (HG b) Size (bp) of products Forward Reverse For grouping PCR c I TGGTTCAAATATCAATGCTC ATTGGTTGTGAGTGCATTG cps3G , cps13G , cps18G Aminotransferase (HG41) 933 II d TCAAAATACGCACCTAAGGC CACTCACCTGCCCCAAGAC cps1Q , cps1/2P , cps2P , cps6Q , cps14P , cps16R , cps27P N -acetylneuraminic acid synthase NeuB (HG10) 823 III e , f TGATTTGGGTGAGACCATG CTCATGCTGGATAACACGT cps21R, cps28P , cps29Q , cps30N N -acetylfucosamine synthase FlnC (HG26) 583 IV ACAGTCGGTCAAGATAATCG TCAGCTTGGGTAATATCTGG cps4F-G , cps5F-G , cps7F-G , cps17G-H , cps19G-H , cps23F-G Initial sugar transferase (HG21) and aminotransferase (HG22) 455 V g GGAAAGATGGAGGACCAGC CCAACCAGACTCATATCCCC cps8E , cps15E , cps20E , cps22E , cps25E Initial sugar transferase (HG6) 265 III and VI e , h GATGCCCCAAGCGATATGCC (F1), GACGCACCAAGTGATATGCC (F2) GGACCAACAATGGCCATCTC (R1), GGTCCGACAATAGCCATTTC (R2) Group III: cps21G , cps28F , cps29G , cps30F ; group VI, cps9F , cps10F , cps11F , cps12F , cps24G , cps26H , cps33G Initial sugar transferase (HG8) 146 For typing PCR cps group I 3 GGTTTTGATTGGTCTAGTTG CTCTAAAGCTCGATATCTAC cps3L Wzy polymerase (HG90) 214 13 TATGGTTAAAGGTGGAACTG CCTTGTATATATTCCCTCCA cps13L Wzy polymerase (HG148) 408 18 TAATGGGATAGTTGCGTTAC ATACATAAAGTTGTCCTGCG cps18N Wzy polymerase (HG170) 617 cps group II 2 and 1/2 TTAGCAACGTTGCCAATAAG AATCCTCCATTAAAACCCTG cps2I , cps1/2I Wzy polymerase (HG54) 173 6 GCTCACTATTTTTACATTACAC TATTACTCCGCCAAATACAG cps6I Wzy polymerase (HG109) 278 1 and 14 TTAGACAGACACCTTATAGG CTAGCTTCGTTACTTGATTC cps14H , cps1I Wzy polymerase (HG50) 386 16 AAGGTTATCCACGAAAGATG TCCGGCAATATTCTTTCAAG cps16I Wzy polymerase (HG362) 494 27 AGACACTGCTTGCATTATTG TCAGAATTACTTCCTGTTGC cps27K Wzy polymerase (HG246) 655 cps group III 21 TATCATATTGAGAATCTTCCC TTGCGTAGCATACAAAGTTC cps21P Wzy polymerase (HG194) 160 28 ATTATGTTGGTTGCAGAAGG CGACTCAATTGTTGTAGTAG cps28L Wzy polymerase (HG254) 272 29 TTCTGGGATTTTAGGAATGC CATGAAATACGCACTTGTAC cps29L Wzy polymerase (HG259) 415 30 TATTGCACTAGCTTCAGAAC TGCATCCATAGTTGTATTCG cps30I Wzy polymerase (HG264) 568 cps group IV 4 GACTATCTGTATACCCAAAC TCCTTCCAAGTATTCTCTAG cps4K Wzy polymerase (HG96) 903 5 ATCTTAGGAATGATTCGGAC ACCAGATATCTGAGCAAATG cps5L Wzy polymerase (HG103) 720 7 AACTACCTACCTGAACTTTG AGTCTAAAAGTGATCGAGTC cps7L Wzy polymerase (HG113) 566 17 TAGCATCAGTTTATACGAGG TAGTTTATCTGTGACACACC cps17O Wzy polymerase (HG164) 455 19 GTGTCGCAAATCAAGTATTG AAGCTAGTACAACAAGCATG cps19L Wzy polymerase (HG174) 348 23 TAATGTATGCTCTGTCACTG AACGAAACGGAATAGTTTGC cps23J Wzy polymerase (HG213) 221 cps group V 8 AAATAAGGTAGGAGCTACTC ATCCAACCTTAGCTTTCTGT cps8K Wzy polymerase (HG120) 446 15 ATCGTTTTGAGATTGAGTGG TAAACGGATTCGGTTACTCA cps15K Wzy polymerase (HG153) 542 20 TGTGGATTTCTGGGATAATC TGTGGACGAATTACTACTTG cps20I Wzy polymerase (HG182) 698 22 GCATTATCAGGATTCTTTCC CCAATTGGGTGTTCAAAAAG cps22K Wzy polymerase (HG200) 296 25 GTTTGCTCCGATCATAATAG CCAGTAAAAGGACTCAATAC cps25M Wzy polymerase (HG229) 174 cps group VI 9 GAAAGTAGGTATATCTCAGC GGGCTATTAAAACTCCTATC cps9J Wzy polymerase (HG123) 368 10 TTTCCCATTTGCTTATGGAC GGAATAAAAACGATTGGGAG cps10M Wzy polymerase (HG130) 633 11 ATGCGATTGCAACAATTGAC AGGCATGAGTAATACATAGG cps11N Wzy polymerase (HG138) 833 12 AACAGGTATTTCAGGATTGC CTCGGATAAAGATAATCAGC cps12J Wzy polymerase (HG140) 131 24 TACTGAGATTTATTGGGACG AAGCGATTGGATTACATTGC cps24M Wzy polymerase (HG220) 224 26 TTATACCGAAATTTTGTTGCC CGTCAATCATATAAAGTGGG cps26P Wzy polymerase (HG240) 472 33 GATGTTTTCAACAGGTGTAC CAAAGTACCTATTTTCAGCG cps33K Wzy polymerase (HG286) 710
on May 16, 2020 by guest
http://jcm.asm.org/
[image:3.585.66.531.71.721.2]nontypeable isolates may have mismatch sequences in their primer sites or may be novelcpstypes that cross-react with known typing sera. All PCR assays were repeated on three different occa-sions using representative strains/isolates from all serotypes and nontypeable isolates to confirm the reproducibility, and the same results were obtained (data not shown). The detection limit of the assays determined using reference strains was 1 to 100 pg of puri-fied DNA/reaction (equivalent to approximately 4⫻102to 4⫻ 104copies of chromosome), which was comparable to or slightly
better than those of the reported method (9).
A total of 52 of the 126 nontypeable isolates by coagglutination tests (41.3%) were typed bycpstyping (Table 2). Three of them (NL146, NL194, and NL290) had been confirmed to be acapsular by transmission electron microscopy analysis (16). Furthermore, we found that they had mutations in theircps2gene clusters but still retained the target genes (cps2Iandcps2P) forcpstyping
[image:4.585.296.544.77.469.2](ref-cps group VII 31 ACAATCGTTTCTGCAATACG GATGAAAACATCGTTGGTAG cps31L Wzy polymerase (HG274) 842 ATCAGTAGTGGGAATAGTTG TTTACTGTTTTTCGACCGTG cps31G Initial sugar transferase (HG269) 423 32 AACCGCTGTTGAATTAAGAG TTCGTTAGTTGAACTGTTCC cps32H Wzy polymerase (HG281) 570 TAGGACTATGGTTCCTAATG TATTCTAGTTCAAGTCGCTC cps32E Initial sugar transferase (HG278) 342 34 AAGTTTCATTCGAGGACTTC GTATATAACACCGCAAGAAG cps34M Wzy polymerase (HG287) 246 ATACAGTGATGTCTTGCAAC ATTGCTTTTTGACAATCGGC cps34E Initial sugar transferase (HG290) 701 For all PCR Internal control GAGTTTGATCCTGGCTCAG AGAAAGGAGGTGATCCAGCC 16S rRNA 1,542–1,553 aRefer to our previous study ( 8 ) for gene names. bHG, homology group of putative cps gene products assigned in our previous study ( 8 ). cOnly the internal control products were amplified from cps group VII strains. dSerotype 13 reference strain has a homologue of target genes for cps group II (putative N -acetylneuraminic acid synthase gene, cps13O ). However, the primers were designed to not detect this gene. eIn addition to the internal control products, cps group III strains produced two cps gene products by the grouping PCR (146-and 583-bp products), and the 146-bp product is common with cps group VI strains. fSerotype 17 reference strain has a homologue of target genes for cps group III (putative N -acetylfucosamine synthase gene, cps17R ). However, the primers were designed to not detect this gene. gAll serotype reference strains in cps -group II also possess the homologues of target genes for cps group V (genes encoding putative initial sugar transferase of HG6). However, the primers were designed not to detect these strains. hBecause the genes encoding putative initial sugar transferase of HG8 have sequence variations among serotype reference strains in groups III and VI, two primers were designed for each primer site to amplify the 146-bp products. Of note, all reference strains in groups III and VI were detected only when the four primers were mixed, while with every single primer pair (forward 1 [F1] -reverse 1 [R1], F1-R2, F2-R1, or F2-R2), some of the reference strains in groups III and VI were not detected.
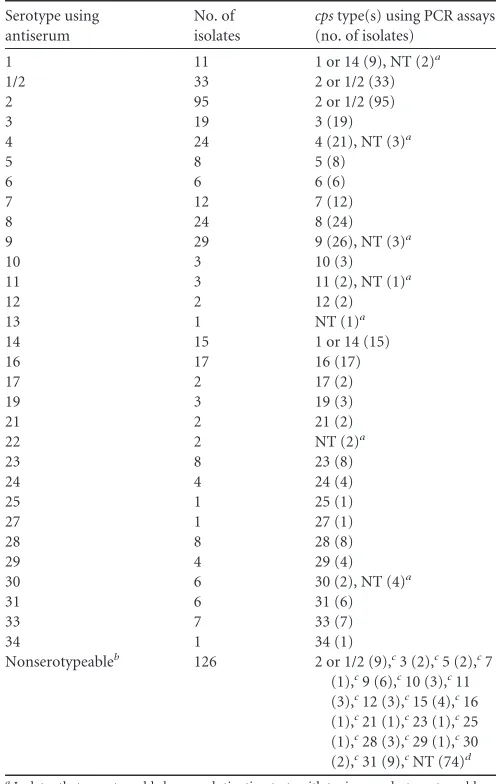
TABLE 2Results of serotyping andcpstyping of the 483 isolates tested
Serotype using antiserum
No. of isolates
cpstype(s) using PCR assays (no. of isolates)
1 11 1 or 14 (9), NT (2)a
1/2 33 2 or 1/2 (33)
2 95 2 or 1/2 (95)
3 19 3 (19)
4 24 4 (21), NT (3)a
5 8 5 (8)
6 6 6 (6)
7 12 7 (12)
8 24 8 (24)
9 29 9 (26), NT (3)a
10 3 10 (3)
11 3 11 (2), NT (1)a
12 2 12 (2)
13 1 NT (1)a
14 15 1 or 14 (15)
16 17 16 (17)
17 2 17 (2)
19 3 19 (3)
21 2 21 (2)
22 2 NT (2)a
23 8 23 (8)
24 4 24 (4)
25 1 25 (1)
27 1 27 (1)
28 8 28 (8)
29 4 29 (4)
30 6 30 (2), NT (4)a
31 6 31 (6)
33 7 33 (7)
34 1 34 (1)
Nonserotypeableb 126 2 or 1/2 (9),c3 (2),c5 (2),c7
(1),c9 (6),c10 (3),c11
(3),c12 (3),c15 (4),c16
(1),c21 (1),c23 (1),c25
(1),c28 (3),c29 (1),c30
(2),c31 (9),cNT (74)d
aIsolates that were typeable by coagglutination tests with typing sera but nontypeable
(NT) bycpstyping PCR assays.
bNonserotypeable isolates, include autoagglutinating, multiagglutinating, and
nonagglutinating strains.
cIsolates that were typeable bycpstyping PCR assays but not by the coagglutination
test.
dIsolates that were nontypeable by either coagglutination tests orcpstyping PCR assays.
on May 16, 2020 by guest
http://jcm.asm.org/
erence16and our unpublished data). Bacterial cells of these three isolates showed autoagglutination, and their antigens extracted by autoclaving did not react with any typing antisera (data not shown). Many of the other 49 nontypeable isolates also showed similar characteristics (data not shown). Thus, these isolates might also be acapsular due to mutations in thecpsgenes, such as NL146, NL194, and NL290, although we cannot rule out the pos-sibility that some of them are encapsulated and have different antigenicity from the strains of the known 35 serotypes. Although CP was believed to be essential for the virulence ofS. suis(6,17, 18), the biological significance of the acapsular phase in the patho-genesis ofS. suisinfection has been proposed recently (16,19). Many acapsularS. suisisolates have been recovered from porcine cases of endocarditis (16). Therefore, the typing methods devel-oped are also considered to be useful for epidemiological studies of acapsular strains by enabling determination of the genotypes of thecpsgene clusters.
Some isolates were regarded as nonserotypeable in our valida-tion tests due to multiagglutinating characteristics with several typing sera. Liu et al. (9) recently identified serotype 21/29 isolates that react with both serotype 21 and 29 typing sera and demon-strated that one of the isolates had a mosaiccpsgene cluster con-sisting of serotype 21 and 29cpsgene homologues. Therefore, this may also have occurred in our multiagglutinating isolates, and novelcpsgene clusters may have been generated. Furthermore, 74 isolates in this study could not be typed by either serotyping or PCR-basedcpstyping. These isolates may be acapsular because of a large deletion in thecpsgene clusters, including the target genes. Most of them had strong hydrophobic and autoagglutinating characteristics, strongly suggesting that they are nonencapsulated (12). However, a few strains were weakly hydrophobic, implying that they have completely newcpsgene clusters. So far, the CP structures have been determined only for serotypes 2 and 14 (20, 21), andcpsgene clusters of most of the nonserotypeable isolates have not been analyzed yet. Further analysis, including structure determination of the CPs andcpsgene clusters ofS. suisstrains with different antigenicity, may provide insights into the evolu-tion ofS. suisCP as well as the relationship betweencpstypes and serotypes.
The presence of many different serotypes in theS. suis popula-tion suggests the continuous and ongoing evolupopula-tion ofcpsgene clusters in this species. Therefore, all reported molecular serotyp-ing methods, includserotyp-ing ours, should lead to improvements in the typing of more field strains. A few serotyped isolates were assigned to the expected serotypes by our method, but could not be typed by the method developed by Liu and colleagues (9), and vice versa (data not shown). However, the presence of a number of nonse-rotypeable isolates that can be assigned to thecpstypes in this study, as well as the consistent results ofcpstyping with serotyping that were observed in most of the tested isolates, including those from many clinical cases, strongly suggest that the developedcps
typing methods are practical and can further contribute to the diagnosis and epidemiological studies ofS. suisinfections.
ACKNOWLEDGMENTS
This work was supported by a Grant-in-Aid for Young Scientists (B) (23780310) and a Grant-in-Aid for Exploratory Research (24659197) from the Japan Society for the Promotion of Science and a Grant-in-Aid for Scientific Research (C) (22592032, 23580420, and 24590525) from the Ministry of Education, Culture, Sports, Science and Technology, Japan,
and was supported partially by the Natural Sciences and Engineering Re-search Council of Canada (NSERC) (grant 154280).
REFERENCES
1.Perch B, Pedersen K, Henrichsen J.1983. Serology of capsulated
strep-tococci pathogenic for pigs: six new serotypes ofStreptococcus suis. J. Clin. Microbiol.17:993–996.
2.Gottschalk M, Higgins R, Jacques M, Beaudoin M, Henrichsen J.1991.
Characterization of six new capsular types (23 through 28) ofStreptococcus suis. J. Clin. Microbiol.29:2590 –2594.
3.Gottschalk M, Higgins R, Jacques M, Mittal K, Henrichsen J. 1989.
Description of 14 new capsular types ofStreptococcus suis. J. Clin. Micro-biol.27:2633–2636.
4.Higgins R, Gottschalk M, Boudreau M, Lebrun A, Henrichsen J.1995.
Description of six new capsular types (29 –34) ofStreptococcus suis. J. Vet. Diagn. Invest.7:405– 406.http://dx.doi.org/10.1177/104063879500700322.
5.Staats JJ, Feder I, Okwumabua O, Chengappa MM.1997.Streptococcus
suis: past and present. Vet. Res. Commun.21:381– 407.http://dx.doi.org /10.1023/A:1005870317757.
6.Smith HE, Damman M, van der Velde J, Wagenaar F, Wisselink HJ,
Stockhofe-Zurwieden N, Smits MA.1999. Identification and
character-ization of thecpslocus ofStreptococcus suisserotype 2: the capsule protects against phagocytosis and is an important virulence factor. Infect. Immun. 67:1750 –1756.
7.Smith HE, de Vries R, van’t Slot R, Smits MA.2000. Thecpslocus of
Streptococcus suisserotype 2: genetic determinant for the synthesis of sialic acid. Microb. Pathog.29:127–134.http://dx.doi.org/10.1006/mpat.2000 .0372.
8.Okura M, Takamatsu D, Maruyama F, Nozawa T, Nakagawa I, Osaki
M, Sekizaki T, Gottschalk M, Kumagai Y, Hamada S. 2013. Genetic
analysis of capsular polysaccharide synthesis gene clusters from all sero-types ofStreptococcus suis: potential mechanisms for generation of capsu-lar variation. Appl. Environ. Microbiol.79:2796 –2806.http://dx.doi.org /10.1128/AEM.03742-12.
9.Liu Z, Zheng H, Gottschalk M, Bai X, Lan R, Ji S, Liu H, Xu J.2013.
Development of multiplex PCR assays for the identification of the 33 serotypes ofStreptococcus suis. PLoS One8:e72070.http://dx.doi.org/10 .1371/journal.pone.0072070.
10. Hill JE, Gottschalk M, Brousseau R, Harel J, Hemmingsen SM, Goh SH.
2005. Biochemical analysis,cpn60and 16S rDNA sequence data indicate thatStreptococcus suisserotypes 32 and 34, isolated from pigs, are Strepto-coccus orisratti. Vet. Microbiol.107:63– 69.http://dx.doi.org/10.1016/j .vetmic.2005.01.003.
11. Tien le HT, Nishibori T, Nishitani Y, Nomoto R, Osawa R. 2013.
Reappraisal of the taxonomy ofStreptococcus suisserotypes 20, 22, 26, and 33 based on DNA-DNA homology andsodAandrecNphylogeny. Vet. Microbiol.162:842– 849.http://dx.doi.org/10.1016/j.vetmic.2012.11.001.
12. Gottschalk M, Lacouture S, Bonifait L, Roy D, Fittipaldi N, Grenier D.
2013. Characterization ofStreptococcus suisisolates recovered between 2008 and 2011 from diseased pigs in Québec, Canada. Vet. Microbiol. 162:819 – 825.http://dx.doi.org/10.1016/j.vetmic.2012.10.028.
13. Dorsch M, Stackebrandt E.1992. Some modifications in the procedure of
direct sequencing of PCR amplified 16S rDNA. J. Microbiol. Methods 16:271–279.http://dx.doi.org/10.1016/0167-7012(92)90017-X.
14. Okwumabua O, O’Connor M, Shull E.2003. A polymerase chain
reac-tion (PCR) assay specific forStreptococcus suisbased on the gene encoding the glutamate dehydrogenase. FEMS Microbiol. Lett.218:79 – 84.http: //dx.doi.org/10.1111/j.1574-6968.2003.tb11501.x.
15. Higgins R, Gottschalk M. 1990. An update on Streptococcus suis
identification. J. Vet. Diagn. Invest.2:249 –252.http://dx.doi.org/10.1177 /104063879000200324.
16. Lakkitjaroen N, Takamatsu D, Okura M, Sato M, Osaki M, Sekizaki T.
2011. Loss of capsule amongStreptococcus suisisolates from porcine en-docarditis and its biological significance. J. Med. Microbiol.60:1669 – 1676.http://dx.doi.org/10.1099/jmm.0.034686-0.
17. Chabot-Roy G, Willson P, Segura M, Lacouture S, Gottschalk M.2006.
Phagocytosis and killing ofStreptococcus suisby porcine neutrophils. Mi-crob. Pathog.41:21–32.http://dx.doi.org/10.1016/j.micpath.2006.04.001.
18. Benga L, Fulde M, Neis C, Goethe R, Valentin-Weigand P. 2008.
Polysaccharide capsule and suilysin contribute to extracellular survival of
Streptococcus suisco-cultivated with primary porcine phagocytes. Vet. Mi-crobiol.132:211–219.http://dx.doi.org/10.1016/j.vetmic.2008.05.005.
on May 16, 2020 by guest
http://jcm.asm.org/
19. Tanabe S, Bonifait L, Fittipaldi N, Grignon L, Gottschalk M, Grenier D. 2010. Pleiotropic effects of polysaccharide capsule loss on selected biolog-ical properties ofStreptococcus suis. Can. J. Vet. Res.74:65–70.http://dx .doi.org/10.1128/JCM.01687-08.
20. Van Calsteren MR, Gagnon F, Lacouture S, Fittipaldi N, Gottschalk M.
2010. Structure determination ofStreptococcus suisserotype 2 capsular
polysaccharide. Biochem. Cell Biol. 88:513–525. http://dx.doi.org/10 .1139/O09-170.
21. Van Calsteren MR, Gagnon F, Calzas C, Goyette-Desjardins G, Okura
M, Takamatsu D, Gottschalk M, Segura M.2013. Structure
determina-tion ofStreptococcus suisserotype 14 capsular polysaccharide. Biochem. Cell Biol.91:49 –58.http://dx.doi.org/10.1139/bcb-2012-0036.