Dendritic cells and macrophages in the spleen
Beijer, M.R.
2014
document version
Publisher's PDF, also known as Version of record
Link to publication in VU Research Portal
citation for published version (APA)
Beijer, M. R. (2014). Dendritic cells and macrophages in the spleen: Development, function, and collaboration.
General rights
Copyright and moral rights for the publications made accessible in the public portal are retained by the authors and/or other copyright owners and it is a condition of accessing publications that users recognise and abide by the legal requirements associated with these rights. • Users may download and print one copy of any publication from the public portal for the purpose of private study or research. • You may not further distribute the material or use it for any profit-making activity or commercial gain
• You may freely distribute the URL identifying the publication in the public portal ?
Take down policy
If you believe that this document breaches copyright please contact us providing details, and we will remove access to the work immediately and investigate your claim.
E-mail address:
mediated bacterial killing by splenic
dendritic cells
Marieke R. Beijer, Ronald Backer, Ellen G.F. Borg, Ayla Kempers,
Georg Kraal, Joke M.M. den Haan
Department of Molecular Cell Biology and Immunology, VU University Medical Center, P.O. Box 7057, Amsterdam, The Netherlands
7
Abstract
Dendritic cells (DCs) continuously scavenge their environment to capture invading pathogens for presentation to T cells. To increase the antigen presentation potential, DCs are equipped with several mechanisms that reduce antigen degradation. Alkalization of the endosomal and phagosomal compartments by reactive oxygen species (ROS) produced by NADPH oxidases is one of these mechanisms. In neutrophils, the respiratory burst mediated by NADPH oxidase results in effective bacterial killing, but whether DCs also eliminate bacteria via NADPH oxidase activity is not clear. Here we show that murine splenic DC subsets differentially express NADPH oxidase subunits and differentially produce ROS. Furthermore, preliminary data indicate that killing of Salmonella typhimurium by DCs is dependent on NADPH oxidase activity. These results suggest that DCs may utilize NADPH oxidase activity to eliminate phagocytosed bacteria.
7
Introduction
Phagocytes such as neutrophils, macrophages, and dendritic cells (DCs) continuously scavenge their environment for the presence of harmful pathogens. Neutrophils and macrophages completely destroy engulfed pathogens while DCs are required to initiate an adaptive immune response. Fusion of phagocytes with endosomal and lysosomal compartments recruits NADPH oxidases, vacuolar ATPases and hydrolases to the phagosomes. As these phagosomes thereby maintain a proper environment for
proteolytic enzyme activity, pathogens are completely degraded 1. In DCs, phagocytosis
serves a different purpose, as immunogenic peptides for antigen presentation should
be generated. Therefore, DCs express less proteolytic enzymes, with reduced activity 2,3.
Together, this prevents complete antigen degradation in order to generate peptides that can be presented to T cells.
The NADPH oxidase complex consists of five subunits and produces reactive
oxygen species (ROS). In steady state, gp91phox and p22phox are located in the plasma
or phagosomal membrane 4 while the activating complex formed by p40phox, p47phox,
and p67phox is located in the cytoplasm 5. Upon cell priming by different stimuli such as
proinflammatory cytokines like TNFα and IL1β and TLR agonists, p47phox is recruited to
the plasma- or phagosomal membrane, where it interacts with gp91phox and p22phox. At
this stage, no ROS is produced but additional activation by for example PMA is required
to initiate the production of ROS 6. Dependent on the cell type, one of two Rac GTPases
(Rac1 and Rac2) participates in the assembly of the NADPH oxidase complex 7,8. The
NADPH oxidase complex transfers an electron from NADPH at the cytosolic side of the
complex to O2 that is located outside the cell or inside the phagosome. This results in the
formation of a superoxide radical (×O2-) that is converted into hydrogen peroxide (H2O2)
and nitric oxide (NO) derived peroxynitrite (ONOO-). Hydrogen peroxide is bactericidal
only at high concentrations 9 while peroxynitrite decomposes to generate the strong
oxidants OH× and NO2× 10. The generated superoxides mediate bacterial killing directly.
In addition, the electron transport over the cell- or phagomal membrane, results in a charge difference, which is neutralized by the influx of potassium ions. This leads to the
activation of intraphagosomal peptides, which also mediates bacterial killing 11. Patients
with chronic granulomatous disease (CGD) where mutations in one of the NADPH oxidase complex subunits cause recurrent fungal and bacterial infections dramatically illustrate
the importance of a functional NADPH oxidase complex 6,12 .
While elimination of pathogens is the most important function for phagocytes, DCs are specialized in the activation of adaptive immune responses, which depends on antigen presentation to T cells. To increase antigen presentation, DCs are equipped with several mechanisms that limit proteolysis. This allows them to present peptides derived from the engulfed antigens to initiate an adaptive immune response. Compared
to macrophages and neutrophils, DCs express low levels of proteolytic enzymes 3. Also
inefficient lysosomal acidification due to low V-ATPase activity limits proteolysis and
7
alkalization of the endosomal and phagosomal compartments by NADPH oxidase is
used to further reduce antigen degradation and thereby induces antigen presentation 14.
These studies clearly indicate a role for NADPH oxidase in antigen presentation by DCs, but whether DCs also utilize this enzyme to kill bacteria is unclear.
In the spleen different DC subsets have been identified according to the expression
of CD4 and CD8 15. These DC subsets are located in distinct areas of the spleen, are
dependent on different differentiation stimuli, express specific surface markers, and
exert distinct functions. CD4+ and CD4-CD8- (double negative (DN)) DCs (together referred
to as CD8- DCs) are specialized in the presentation of antigens via MHC-II towards CD4+
T cells 16,17. On the other hand, CD8+ DCs are specialized in cross-presentation of exogenous
antigens in MHC-I molecules via which CD8+ T cells are activated 18. Differences in NADPH
oxidase subunit expression between CD8+ and CD8- DCs have been reported by us and
others 14,19. Interestingly, Savina et al. recently showed that Rac2 is highly expressed in
CD8+ DCs and determines the subcellular location of the NADPH oxidase complex to the
phagosomes. In contrast, in CD8- DCs, Rac1 mediates the assembly of the complex at the
plasma membrane 4. Therefore, CD8- DCs exert higher production of extracellular ROS,
while only CD8+ DCs are able to produce ROS in the phagosomes 20. This specific capacity
of CD8+ DCs to produce ROS in the phagosomes resulted in an enhanced capacity to
cross-present antigens 14, but whether this also mediates differential bacterial killing is
not yet established.
Here, we investigated the capacity of DC subsets to produce ROS and performed
preliminary studies on the bacterial killing capacity of DCs. We demonstrate that CD4+ and
DN DCs show higher expression of NADPH oxidase complex subunits and generate more
ROS than CD8+ DCs. Furthermore, using flt3L bone marrow derived DCs, we determined
that the expression of NADPH oxidase subunits is stimulated by vitamin A. Preliminary experiments showed that bacterial killing by bone marrow derived DCs is dependent on their ability to produce ROS. Further research will be necessary to elucidate the bacterial killing capacity of DC subsets in vitro and in vivo and its dependence on NADPH oxidase.
Materials and methods
Mice
C57BL/6 mice were purchased at Charles River Laboratories (Maastricht, The Netherlands) and were used at 8-16 weeks of age. The ethical committee of the VU University Medical Center approved all animal experiments.
DC isolation and cell sorting
Splenocytes were isolated from C57BL/6 mice as previously described 21. Briefly, spleens
were cut into grain size fragments and digested with 1 WU/ml liberase TL (Roche Diagnostics GmbH, Mannheim, Germany) and 0.2 mg/ml DNAse (Roche) for 20-40 minutes at 37°C while stirring. Cells were washed with RPMI medium containing 10% FCS, 2% penicillin-streptomycin, 2% L-glutamine, 10 mM EDTA and 20 mM Hepes. Red blood cells
7
were lysed by incubation with 1 ml ACK lysis buffer (0,15M NH4Cl, 10 mM KHCO3 0.1 mM
Na2EDTA at pH 7.2–7.4) for 1 minute at room temperature. Non-digested fragments were
removed by filtration. Subsequently, CD11c+ cells were isolated by positive selection with
anti-CD11c MACS microbeads (Miltenyi Biotec, Bergisch Gladbach, Germany) according
to manufacturers’ protocol. Purified CD11c+ cells were stained using antibodies against
CD11c, CD4 and CD8 (eBioscience, San Diego, CA). Live non-autofluorescent CD11chigh
cells were sorted in individual DC populations (90-95% pure) using MoFlo XDP cell sorter (Beckman Coulter, Woerden, The Netherlands). Cells were stored in TRIzol reagent (Invitrogen Life Technologies, Breda, The Netherlands) at -80°C immediately after reanalysis until further use.
Bone marrow derived DC cultures and cell sorting
Bone marrow derived DCs were generated as described 22. In brief, bone marrow was
isolated from C57/BL6 mice and cultured at a concentration of 3·106 cells/ml in the
presence of 200 ng/ml Flt3L in RPMI 1640 culture medium supplemented with 10% heat inactivated FCS, 2% penicillin-streptomycin, 2% L-glutamine. After 9-10 days, cells were harvested for analysis. Where indicated, cells were stimulated with 10 nM RA, 18h prior to harvesting.
For cell sorting, day 9 bone marrow derived DCs were stained for CD11c, Sirpα
and CD24 (eBioscience, San Diego, CA). Live non-autofluorescent CD11chigh cells were
sorted in individual DC populations (90–95% pure) using MoFlo XDP cell sorter (Beckman Coulter, Woerden, The Netherlands). Cells were stored in TRIzol reagent (Invitrogen Life Technologies, Breda, The Netherlands) at -80°C immediately after reanalysis until further use.
mRNA isolation, cDNA synthesis and quantitative PCR
Total RNA was isolated from sorted ex vivo DCs or bone marrow derived DCs using TRIzol reagent (Invitrogen Life Technologies, Breda, The Netherlands). RNA was precipitated
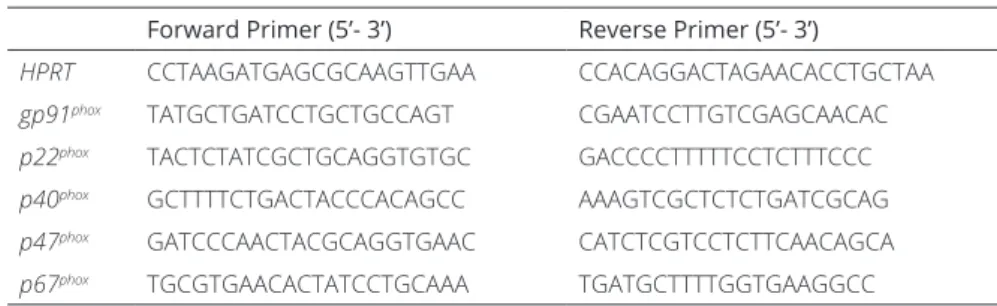
Table 1. Primers sequences used for quantitative polymerase chain reaction.
Forward Primer (5’- 3’) Reverse Primer (5’- 3’)
HPRT CCTAAGATGAGCGCAAGTTGAA CCACAGGACTAGAACACCTGCTAA
gp91phox TATGCTGATCCTGCTGCCAGT CGAATCCTTGTCGAGCAACAC
p22phox TACTCTATCGCTGCAGGTGTGC GACCCCTTTTTCCTCTTTCCC
p40phox GCTTTTCTGACTACCCACAGCC AAAGTCGCTCTCTGATCGCAG
p47phox GATCCCAACTACGCAGGTGAAC CATCTCGTCCTCTTCAACAGCA
p67phox TGCGTGAACACTATCCTGCAAA TGATGCTTTTGGTGAAGGCC
HPRT = hypoxanthine guanine phosphoribosyl transferase, gp91phox = CYBB (cytochrome B-245 beta polypeptide), p22phox = CYBA (cytochrome B-245 alpha polypeptide), p40phox = NCF4 (neutrophil cytosolic factor 4), p47phox = NCF1 (neutrophil cytosolic factor 1), p67phox = NCF2 (neutrophil cytosolic factor 2). Primers were designed using the online NCBI primer design tool (http://www.ncbi.nlm.nih.gov/tools/primer-blast/).
7
with isopropanol and cDNA was synthesized using RevertAid First Strand cDNA Synthesis Kit (Fermentas Life Sciences, Vilnius, Lithuania) according to manufacturers’ protocol. Quantitative PCR was performed using SYBR Green Mastermix (PE Applied Biosystems, Foster City, CA) on an ABI Prism 7900HT Sequence Detection System (PE Applied Biosystems Foster City, CA). A standard curve was generated using pooled lymph node tissue to correct for primer efficiency. mRNA quantities were normalized to HPRT.
Detection of reactive oxygen species by amplex red assay
ROS production by bone marrow derived DCs upon phorbol myristate acetate (PMA) stimulation (1 μg/ml) was determined by measuring the oxidation of the fluorogenic indicator Amplex Red (0.1 units/ml) (Molecular probes) into the fluorescent product resofurine. The NADPH oxidase inhibitor apocynin (1-(4-Hydroxy-3-methoxyphenyl) ethanone, or acetovanillone, Sigma, Missouri, USA) was used where indicated at a concentration of 500 μM. Fluorescence was recorded over time for indicated time period with a fluorstar spectrofluorimeter (BMG Labtec, Offenburg, Germany) equipped with 535 nm excitation and 595 nm emission wavelengths. Relative ROS production is depicted.
Salmonella typhimurium
Virulence of Salmonella typhimurium is dependent on its ability to produce enterochelin 23.
Salmonella typhimurium (SL3261) that was attenuated for the aromatic biosynthetic
pathway (deltaAroA) that produces enterochelin, was kindly provided by Dr. J. Luirink
(department of molecular microbiology, VU medical center, Amsterdam) 24.
Bacterial killing assay
An overnight culture of Salmonella typhimurium was diluted until OD600 0.2 and grown for 3 hours in LB medium until bacteria reached logarithmic growth phase. Salmonella was opsonized by incubation with 50% normal mouse serum for 20 minutes at 37°C to ensure uptake by DCs. Salmonella was washed twice in RPMI containing 10% FCS and
brought to desired concentration assuming OD600 = 1 corresponds to 1.2·109 cells/ml.
Bone marrow derived DCs were incubated with Salmonella at MOI = 10 for 30 minutes
at 37°C and 5% CO2 in RPMI1640 culture medium containing 10% heat inactivated FCS to
allow bacterial uptake. Extracellular bacteria were killed by addition of RPMI 1640 culture medium supplemented with 10% heat inactivated FCS, 2% penicillin-streptomycin and 2% L-glutamine. Apocynin was added at 500 μM as NADPH inhibitor from this step onwards
when indicated. After 30 minutes at 37°C and 5% CO2, free bacteria and antibiotics
were removed by centrifugation and DCs were left for another 3 hours to allow killing of intracellular Salmonella in medium without antibiotics in the presence of apocynin where indicated. DCs were lysed with 0.025% Triton X-100 to release the intracellular
Salmonella, and a dilution range was plated on Salmonella-Shigella specific agar plates
(Becton Dickinson Microbiology Systems, Maryland, USA). Colonies were counted after overnight incubation at 37°C.
7
Statistical analysis
Statistical significance was tested using GraphPad Prism 4 (La Jolla, CA) by performing a two-tailed Student’s T-test or ANOVA with Bonferroni’s correction as indicated.
Results
CD8- dendritic cells express higher levels of NADPH oxidase subunits and produce more ROS
Various studies have shown differential gene expression in CD4+, CD8+ and DN DCs in the
spleen 16,25. We determined the expression of NADPH oxidase subunits in sorted CD4+,
CD8+ and DN DCs by quantitative PCR. CD4+, CD8+ and DN DCs expressed the five subunits
of the NADPH oxidase complex. However, CD4+ and DN DCs expressed significantly higher
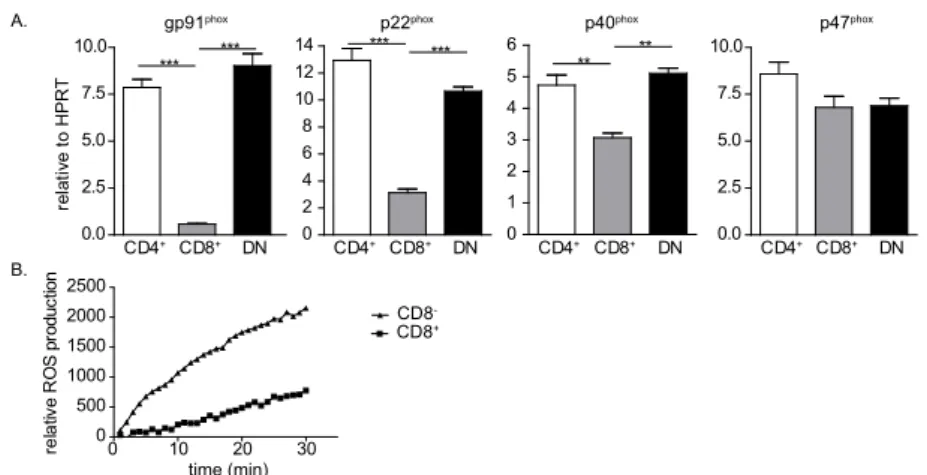
levels of gp91phox, p40phoxand p22phox transcripts than CD8+ DCs (figure 1A). No significant
differences were found between CD4+ and DN DCs in the expression of these genes.
This is in line with previous publications, in which CD4+ and DN DCs showed similar gene
expression profiles 16,25.
To determine whether these differences in expression of the NADPH oxidase subunits was also reflected in the activity of the NADPH oxidase complex, we investigated
the production of reactive oxygen species (ROS) in FACS sorted CD8- and CD8+ DCs. ROS
production was determined in PMA stimulated CD8- and CD8+ DCs by measuring the
oxidation of the fluorogenic substrate amplex red into the fluorescent product resofurine.
CD8- DCs exerted higher ROS production capacity than CD8+ DCs (figure 1B). These data
indicate that higher expression of NADPH oxidase subunits in CD8- DCs results in the
generation of more ROS in sorted splenic DCs.
Figure 1. Ex vivo dendritic cell subsets differentially express NADPH oxidase subunits and differentially produce reactive oxygen species. (A) mRNA expression of NADPH oxidase subunits in sorted CD11chighCD4+, CD11chighCD8+ and CD11chighCD4-CD8- (double negative (DN)) DCs was determined
by quantitative PCR. mRNA levels were normalized to HPRT. (B) Production of reactive oxygen species by FACS sorted CD11chighCD8- (triangles) and CD11chighCD8+ (squares) DCs in the presence of 1 μg/ml PMA
measured by amplex red based luminescence assay. *P<0.05, **P<0.01, ***P<0.001, one-way ANOVA using Bonferonni’s correction. Data are shown as mean + SEM of triplicate wells and are representative of at least 1 (B) or 2 (A) experiments performed.
A. gp91phox CD4+ CD8+ DN 0.0 2.5 5.0 7.5 10.0 *** *** p22phox CD4+ CD8+ DN 0 2 4 6 8 10 12 14 *** *** p40 phox CD4+ CD8+ DN 0 1 2 3 4 5 6 ** ** p47phox CD4+ CD8+ DN 0.0 2.5 5.0 7.5 10.0 re la tiv e to H P R T B. 0 10 20 30 0 500 1000 1500 2000 2500 re la tive R O S pr odu ct io n CD8CD8+ -time (min)
7
Bone marrow derived DCs produce ROS
Bone marrow derived DCs can be generated by culturing bone marrow in the presence of growth factor, FMS-like tyrosine kinase 3 ligand (Flt3L). As the resulting DCs are not fully matured, they do not express CD4 or CD8. Therefore, other subset specific markers
such as Sirpα and CD24are used to discriminate the CD8- and CD8+ DCs, respectively 22.
To determine whether bone marrow derived DCs also exert differences in the expression of NADPH oxidase subunits, we determined mRNA expression of the NADPH oxidase
subunits in Sirpα+ and CD24+ DCs sorted from bone marrow derived DC cultures. In line
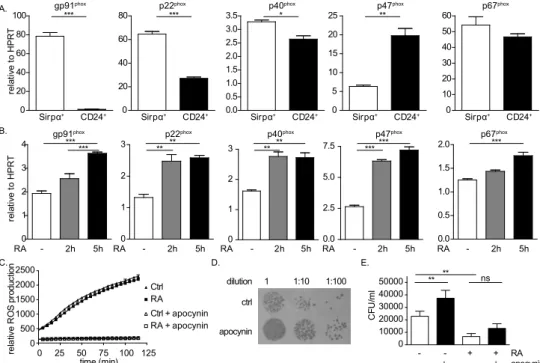
with ex vivo DCs, Sirpα+ DCs expressed significantly more mRNA transcripts of gp91phox,
p40phoxand p22phox than CD24+ DCs. While p47phox expression did not differ in ex vivo DC
subsets, higher p47phox expression was observed in bone marrow derived CD24+ DCs
(figure 2A).
Figure 2. Bacterial killing by bone marrow derived DCs is NADPH oxidase dependent. (A) mRNA expression of NADPH oxidase subunits in sorted CD11c+Sirpα+ and CD11c+CD24+ DCs was determined
by quantitative PCR. mRNA levels were normalized to HPRT. (B) mRNA expression of NADPH oxidase subunits in total bone marrow derived DC cultures stimulated with retinoic acid (RA) for 2 or 5 hours or vehicle control. mRNA levels were normalized to HPRT. (C) Production of reactive oxygen species in total bone marrow derived DC culture in the presence of 1 μg/ml PMA measured by amplex red based luminescence assay. Cells were stimulated with RA (squares) or vehicle control (triangles) 18 hours prior to assay. Apocynin was used at 500 μM as NADPH oxidase inhibitor (open triangles and squares). (D-E) Bone marrow derived DCs and Salmonellatyphimurium were mixed at a multiplicity of infection of 10 and incubated for 30 minutes to allow bacterial uptake by the DCs. Extracellular bacteria were removed after which DCs were incubated for an additional 3 hours in the presence of indicated stimuli to allow bacterial killing. Intracellular bacteria were released and serial dilutions were plated on Salmonella-Shigella specific agar plates. After overnight incubation, bacterial colonies were quantified. ns: not significant, *P<0.05, **P<0.01, ***P<0.001, one-way ANOVA using Bonferonni’s correction. Data are shown as mean + SEM of triplicate wells and are representative of at least 1 (B), 2 (A), or 4 (C-E) experiments performed.
D. 0 10000 20000 30000 40000 50000 ** ** ns C FU /m l - - + + RA - + - + apocynin ctrl 1 1:10 1:100 dilution E. A. Sirpα+ CD24+ 0 20 40 60 80 100 *** Sirpα+ CD24+ 0 20 40 60 80 *** Sirpα+ CD24+ 0.0 0.5 1.0 1.5 2.0 2.5 3.0 3.5 * Sirpα+ CD24+ 0 5 10 15 20 25 ** Sirpα+ CD24+ 0 10 20 30 40 50 60 re la tiv e to H P R T p40phox p67phox
gp91phox p22phox p47phox
- 2h 5h 0 1 2 3 4 *** *** - 2h 5h 0 1 2 3 ** ** - 2h 5h 0 1 2 3 ** ** - 2h 5h 0.0 2.5 5.0 7.5 ****** - 2h 5h 0.0 0.5 1.0 1.5 2.0 *** re la tiv e to H P R T
B. gp91phox p22phox p40phox p47phox p67phox
C. 0 25 50 75 100 125 0 500 1000 1500 2000 2500 time (min) re la tive R O S pr odu ct io n apocynin RA RA RA RA RA Ctrl RA Ctrl + apocynin RA + apocynin
7
Retinoic acid (RA) is described to induce the expression of the NADPH oxidase
subunits gp91phox, p47phoxand p67phox in a human myeloblastic leukemia cellline 26.
To determine whether RA also induces the expression of NADPH oxidase subunits in bone marrow derived DCs, we stimulated bone marrow derived DCs for 2 or 5 hours with RA and determined mRNA expression of the NADPH oxidase subunits by quantitative PCR. Expression of all NADPH oxidase subunits was significantly increased 2 and 5 hours after RA stimulation (figure 2B). Strikingly, this increased expression was not reflected in an increased ROS production (figure 2C). However, since the amplex red assay only
detects extracellular H2O2, predominant intracellular production of ROS or differences in
the production of other types of ROS could possibly explain this discrepancy.
Bacterial killing by bone marrow derived DCs is ROS dependent
In chronic granulomatous disease (CGD) patients, mutations in the NADPH oxidase subunits are associated with recurrent Salmonella infections indicating a role for NADPH
oxidase generated ROS in the killing of Salmonella 27-30. To determine whether ROS is
involved in killing of Salmonella by DCs, we incubated bone marrow derived DCs with opsonized Salmonella typhimurium for 30 minutes to allow bacterial uptake. Subsequently, extracellular Salmonella was washed away with medium containing antibiotics, after which DCs were incubated for 3 hours to allow bacterial killing. Colony forming units were determined after overnight growth on Salmonella-specific agar plates. Inhibition of ROS via the NADPH inhibitor apocynin, resulted in a significant decrease in bacterial killing by the DCs, as indicated by an increase in Salmonella colonies (figure 2D and 2E). Interestingly, even though RA did not increase extracellular ROS production, fewer Salmonella colonies were found when bone marrow derived DCs were stimulated with RA 18 hour prior to and during bacterial killing assay. In addition, blockade of NADPH oxidase in RA treated BM derived DCs resulted in only a small increase of bacterial survival, which suggests that the increase in bacterial killing that was induced by RA incubation was likely to be mediated by other mechanisms than ROS production by NADPH oxidase.
Taken together, these preliminary data indicate that bone marrow derived DCs eliminate bacteria via NADPH oxidase dependent mechanisms. While RA increased the expression of NADPH oxidase components and bacterial killing by DCs, this increased capacity to kill bacteria appeared to be independent on NADPH oxidase activity.
Discussion
Splenic DCs are professional antigen presenting cells that phagocytose pathogens and subsequently present antigens derived from these pathogens. They are equipped with several mechanisms that increase the antigen presentation potential, one of which is the expression of NADPH oxidase which alkalizes the phagosomal and lysosomal compartment thereby reducing antigen degradation. Furthermore, reactive oxygen species (ROS) that are produced by the NADPH oxidase complex mediate bacterial killing. Splenic DCs can
7
exert specific functions. In preliminary studies described in this chapter, we showed
that CD8- DCs express higher levels of the NADPH oxidase subunits than CD8+ DCs and
produced more extracellular ROS in an amplex red assay. These results are in line with
recent studies indicating that differences in Rac1 and Rac2 expression between CD8
-and CD8+ DCs are responsible for differences in the location of NADPH oxidase complex
assembly. Rac2 dependent recruitment of NADPH oxidase subunits to the phagosomes
was observed in CD8+ DCs 8, while increased Rac1 expression mediated the assembly of
NADPH oxidase at the plasmamembrane in CD8- DCs 7. These studies showed a clear role
for the phagosomal NADPH oxidase activity in the excellent cross-presentation capacity
of CD8+ DCs compared to CD8- DCs. However, whether these differential expression
levels and localization of the NADPH oxidase in CD8+ and CD8- DCs are also important
for bacterial killing, is unknown. CD8+ DCs have been described to facilitate the transfer
of bacteria from the marginal zone to the white pulp as they are used as Trojan horse.
Thereby, they are essential for infection with Listeria monocytogenes 31,32. Why only CD8+
DCs carry live bacteria to the T cell area and promote further infection is not clear as
also CD8- DCs take up bacteria 33. One possibility is that CD8+ DCs have lower capacity to
kill intracellular bacteria compared to CD8- DCs, due to the lower expression of NADPH
oxidase subunits.
After oral infection with Salmonella typhimurium, DCs transport the bacteria to the
mesenteric lymph nodes 34,35. Salmonella has been detected in both splenic CD8+ and CD8
-DC subsets upon intravenous infection, which indicates that all subsets can internalize
the bacteria 36. Recently, an increased susceptibility to Salmonella was observed in Sirpα
-/-mice, which have reduced numbers of CD4+ DCs 37,38. However, because these mice
showed defects late after infection and exhibited decreased Salmonella-specific CD4+
T cell responses and antibody responses, the increased susceptibility was likely not due to differential bacterial killing by different DC subsets. Further studies will be necessary to determine the DC subset specific killing capacity of Salmonella and the contribution of NADPH oxidase in the killing of these bacteria by DCs.
Vitamin A deficiency is correlated with increased susceptibility to bacterial
infections 39. Previous studies have shown that the expression of gp91phox, p47phoxand
p67phox, subunits of the NADPH oxidase complex, is vitamin A dependent 26. In addition, we
observed a role for vitamin A in the generation of CD8- DCs that express the highest levels
of the NADPH oxidase subunits 19. Here, we report that the expression of all subunits
of the NADPH oxidase complex is elevated upon retinoic acid (RA) supplementation in bone marrow derived DCs. Furthermore, the Salmonella killing capacity of bone marrow derived DCs is significantly increased upon RA stimulation, although this did not depend on NADPH oxidase activity. Antigen degradation by DCs is tightly controlled by regulation of the expression of proteolytic enzymes and lysosomal acidification via the expression of
V-ATPase 3,13. Further research should determine whether regulation of these mechanisms
explains the observed increase in killing of Salmonella by bone marrow derived DCs upon RA stimulation.
7
Next to bacterial killing, also bacterial uptake or recognition could be affected by vitamin A. Mice lacking toll like receptor (TLR) expression, are more susceptible to
Salmonella infections. Especially TLR2, 4 and 5 are required for the recognition of Salmonella by DCs 40-42 and are differentially expressed by the CD8+ and CD8- DC subsets 43.
More research is required to determine how uptake, recognition, and NADPH oxidase mediated killing of Salmonella by DC subsets is affected by RA stimulation.
Taken together, the results presented in this chapter show differential expression
of NADPH oxidase genes in CD8- and CD8+ DCs. In addition, NADPH oxidase activity is
involved in bacterial killing by DCs. However, more extensive research is required to determine the bacterial killing capacity of DC subsets and its significance for infection.
References
1. Kinchen JM and Ravichandran KS. (2008) Phagosome maturation: going through the acid test.
Nat Rev Mol Cell Biol, 9, 781-795.
2. Delamarre L, Pack M, Chang H, Mellman I and Trombetta ES. (2005) Differential lysosomal proteolysis in antigen-presenting cells determines antigen fate. Science, 307, 1630-1634.
3. Lennon-Dumenil AM, Bakker AH, Maehr R, Fiebiger E, Overkleeft HS, Rosemblatt M, Ploegh HL and Lagaudriere-Gesbert C. (2002) Analysis of protease activity in live antigen-presenting cells shows regulation of the phagosomal proteolytic contents during dendritic cell activation. J.Exp.Med., 196, 529-540.
4. Savina A, Peres A, Cebrian I, Carmo N, Moita C, Hacohen N, Moita LF and Amigorena S.
(2009) The small GTPase Rac2 controls phagosomal alkalinization and antigen crosspresentation selectively in CD8(+) dendritic cells. Immunity, 30, 544-555.
5. Lapouge K, Smith SJ, Groemping Y and Rittinger K. (2002) Architecture of the p40-p47-p67phox
complex in the resting state of the NADPH oxidase. A central role for p67phox. J Biol Chem, 277, 10121-10128.
6. Sareila O, Kelkka T, Pizzolla A, Hultqvist M and Holmdahl R. (2011) NOX2 complex-derived ROS as immune regulators. Antioxid Redox Signal, 15, 2197-2208.
7. Abo A, Pick E, Hall A, Totty N, Teahan CG and Segal AW. (1991) Activation of the NADPH oxidase involves the small GTP-binding protein p21rac1. Nature, 353, 668-670.
8. Knaus UG, Heyworth PG, Evans T, Curnutte JT and Bokoch GM. (1991) Regulation of phagocyte oxygen radical production by the GTP-binding protein Rac 2. Science, 254, 1512-1515.
9. Hyslop PA, Hinshaw DB, Scraufstatter IU, Cochrane CG, Kunz S and Vosbeck K. (1995) Hydrogen peroxide as a potent bacteriostatic antibiotic: implications for host defense. Free Radic Biol Med, 19, 31-37.
10. Radi R, Beckman JS, Bush KM and Freeman BA. (1991) Peroxynitrite oxidation of sulfhydryls. The cytotoxic potential of superoxide and nitric oxide. J Biol Chem, 266, 4244-4250.
11. Reeves EP, Lu H, Jacobs HL, Messina CG, Bolsover S, Gabella G, Potma EO, Warley A, Roes J and Segal AW. (2002) Killing activity of neutrophils is mediated through activation of proteases by K+ flux. Nature, 416, 291-297.
12. Holland SM. (2013) Chronic granulomatous disease. Hematol Oncol Clin North Am, 27, 89-99, viii.
13. Trombetta ES, Ebersold M, Garrett W, Pypaert M and Mellman I. (2003) Activation of lysosomal function during dendritic cell maturation. Science, 299, 1400-1403.
14. Savina A, Jancic C, Hugues S, Guermonprez P, Vargas P, Moura IC, Lennon-Dumenil AM, Seabra MC, Raposo G and Amigorena S. (2006) NOX2 controls phagosomal pH to regulate antigen processing during crosspresentation by dendritic cells. Cell, 126, 205-218.
15. Vremec D, Zorbas M, Scollay R, Saunders DJ, Ardavin CF, Wu L and Shortman K. (1992) The surface phenotype of dendritic cells purified from mouse thymus and spleen: investigation of the CD8 expression by a subpopulation of dendritic cells. J.Exp.Med., 176, 47-58.
7
16. Dudziak D, Kamphorst AO, Heidkamp GF, Buchholz VR, Trumpfheller C, Yamazaki S, Cheong C, Liu K, Lee HW, Park CG et al. (2007) Differential antigen processing by dendritic cell subsets in vivo. Science, 315, 107-111.
17. Liu K and Nussenzweig MC. (2010) Origin and development of dendritic cells. Immunol Rev, 234, 45-54.
18. den Haan JM, Lehar SM and Bevan MJ. (2000) CD8(+) but not CD8(-) dendritic cells cross-prime cytotoxic T cells in vivo. J.Exp.Med., 192, 1685-1696.
19. Beijer MR, Molenaar R, Goverse G, Mebius RE, Kraal G and den Haan JM. (2013) A crucial role for retinoic acid in the development of Notch-dependent murine splenic CD8-CD4- and CD4+
dendritic cells. Eur J Immunol, 43, 1608-1616.
20. Kotsias F, Hoffmann E, Amigorena S and Savina A. (2013) Reactive oxygen species production in the phagosome: impact on antigen presentation in dendritic cells. Antioxid Redox Signal, 18, 714-729.
21. Backer R, van Leeuwen F, Kraal G and den Haan JM. (2008) CD8- dendritic cells preferentially
cross-present Saccharomyces cerevisiae antigens. Eur J Immunol, 38, 370-380.
22. Naik SH, Proietto AI, Wilson NS, Dakic A, Schnorrer P, Fuchsberger M, Lahoud MH, O’Keeffe
M, Shao QX, Chen WF et al. (2005) Cutting edge: generation of splenic CD8+ and CD8- dendritic cell
equivalents in Fms-like tyrosine kinase 3 ligand bone marrow cultures. J Immunol, 174, 6592-6597.
23. Yancey RJ, Breeding SA and Lankford CE. (1979) Enterochelin (enterobactin): virulence factor for Salmonella typhimurium. Infect Immun, 24, 174-180.
24. Hoiseth SK and Stocker BA. (1981) Aromatic-dependent Salmonella typhimurium are non-virulent and effective as live vaccines. Nature, 291, 238-239.
25. Edwards AD, Chaussabel D, Tomlinson S, Schulz O, Sher A and Reis e Sousa C. (2003) Relationships among murine CD11c(high) dendritic cell subsets as revealed by baseline gene expression patterns. J Immunol, 171, 47-60.
26. Shimizu T, Kodama R, Tsunawaki S and Takeda K. (2002) GM-CSF induces expression of gp91phox
and stimulates retinoic acid-induced p47phox expression in human myeloblastic leukemia cells. Eur
J Haematol, 68, 382-388.
27. Mouy R, Fischer A, Vilmer E, Seger R and Griscelli C. (1989) Incidence, severity, and prevention of infections in chronic granulomatous disease. J Pediatr, 114, 555-560.
28. Safe AF, Maxwell RT, Howard AJ and Garcia RC. (1991) Relapsing Salmonella enteritidis infection in a young adult male with chronic granulomatous disease. Postgrad Med J, 67, 198-201.
29. Burniat W, Toppet M and De Mol P. (1980) Acute and recurrent salmonella infections in three children with chronic granulomatous disease. J Infect, 2, 263-268.
30. Moellering RC, Jr. and Weinberg AN. (1970) Persistent Salmonella infection in a female carrier for chronic granulomatous disease. Ann Intern Med, 73, 595-601.
31. Edelson BT, Bradstreet TR, Hildner K, Carrero JA, Frederick KE, Kc W, Belizaire R, Aoshi T, Schreiber RD, Miller MJ et al. (2011) CD8alpha(+) dendritic cells are an obligate cellular entry point for productive infection by Listeria monocytogenes. Immunity, 35, 236-248.
32. Neuenhahn M, Kerksiek KM, Nauerth M, Suhre MH, Schiemann M, Gebhardt FE, Stemberger C, Panthel K, Schroder S, Chakraborty T et al. (2006) CD8alpha+ dendritic cells are required for efficient entry of Listeria monocytogenes into the spleen. Immunity, 25, 619-630.
33. Mitchell LM, Brzoza-Lewis KL, Henry CJ, Grayson JM, Westcott MM and Hiltbold EM. (2011) Distinct responses of splenic dendritic cell subsets to infection with Listeria monocytogenes: maturation phenotype, level of infection, and T cell priming capacity ex vivo. Cell Immunol., 268, 79-86.
34. Swart AL and Hensel M. (2012) Interactions of Salmonella enterica with dendritic cells. Virulence, 3, 660-667.
35. Voedisch S, Koenecke C, David S, Herbrand H, Forster R, Rhen M and Pabst O. (2009) Mesenteric lymph nodes confine dendritic cell-mediated dissemination of Salmonella enterica serovar Typhimurium and limit systemic disease in mice. Infect Immun, 77, 3170-3180.
36. Yrlid U and Wick MJ. (2002) Antigen presentation capacity and cytokine production by murine splenic dendritic cell subsets upon Salmonella encounter. J Immunol, 169, 108-116.
7
37. Saito Y, Iwamura H, Kaneko T, Ohnishi H, Murata Y, Okazawa H, Kanazawa Y, Sato-Hashimoto M, Kobayashi H, Oldenborg PA et al. (2010) Regulation by SIRPalpha of dendritic cell homeostasis in lymphoid tissues. Blood, 116, 3517-3525.
38. Li LX, Atif SM, Schmiel SE, Lee SJ and McSorley SJ. (2012) Increased susceptibility to Salmonella infection in signal regulatory protein alpha-deficient mice. J Immunol, 189, 2537-2544.
39. Stephensen CB. (2001) Vitamin A, infection, and immune function. Annu.Rev.Nutr., 21, 167-192.
40. Arpaia N, Godec J, Lau L, Sivick KE, McLaughlin LM, Jones MB, Dracheva T, Peterson SN, Monack DM and Barton GM. (2011) TLR signaling is required for Salmonella typhimurium virulence. Cell, 144, 675-688.
41. Weiss DS, Raupach B, Takeda K, Akira S and Zychlinsky A. (2004) Toll-like receptors are temporally involved in host defense. J Immunol, 172, 4463-4469.
42. Andersen-Nissen E, Smith KD, Strobe KL, Barrett SL, Cookson BT, Logan SM and Aderem A.
(2005) Evasion of Toll-like receptor 5 by flagellated bacteria. Proc.Natl.Acad.Sci.U.S.A, 102, 9247-9252.
43. Edwards AD, Diebold SS, Slack EM, Tomizawa H, Hemmi H, Kaisho T, Akira S and Reis e
Sousa C. (2003) Toll-like receptor expression in murine DC subsets: lack of TLR7 expression by CD8 alpha+ DC correlates with unresponsiveness to imidazoquinolines. Eur J Immunol, 33, 827-833.