Copyright1999 by the Genetics Society of America
The P-Ph Protein-Mediated Repression of
yellow
Expression Depends on
Different
cis
- and
trans
-Factors in
Drosophila melanogaster
Inna Biryukova,* Tatyana Belenkaya,* Haik Hovannisian,* Elena Kochieva
†and Pavel Georgiev*
*Department of the Control of Genetic Processes, Institute of Gene Biology, Russian Academy of Sciences, Moscow 117334, Russia and
†Department of Biotechnology, Moscow Agriculture Academy, Moscow, 550400, Russia
Manuscript received September 14, 1998 Accepted for publication April 21, 1999
ABSTRACT
The phP1allele of Drosophila melanogaster encodes a chimeric P-Ph protein that contains the DNA-binding
domain of the P-element transposase and the Ph protein lacking 12 amino-terminal amino acids. It has been shown that the P-Ph protein is responsible for the formation of a repressive complex on P elements inserted at the yellow locus. Here we demonstrate that an enhancer element can suppress the P-Ph-mediated inhibition of yellow transcription. However, an increase of P-element copy number at the yellow locus overcomes the enhancer effect. The mobilization of P-element transposition induced the appearance with a high frequency of Su(y) mutations that partially or completely suppressed the inhibitory effect of phP1
on yellow expression. The Su(y) mutations were localized at different sites on chromosomes. One strong
Su(y) mutation, sneP1, was found to be induced by a 1.2-kb P-element insertion into the transcribed noncoding
region of the singed locus. The Su(y) mutations resulted in a high level of transcription of the 1.2-kb P element that contained the sequences encoding one DNA-binding and two protein-protein interaction domains of the transposase. The effect of Su(y) mutations can be explained by the competition between the truncated transposase encoded by a 1.2-kb P element and the P-Ph protein for binding sites on
P-element insertions.
T
HE polyhomeotic (ph) gene is a member of the group amino acids. The P-Ph chimeric protein can bind to theP-element sequences and recruit other Pc-G proteins,
consisting of at least 20 genes with similar functions
that are required for normal segmental specification leading to the formation of a repressive complex. Previously we described a family of highly unstable in Drosophila and referred to as the Polycomb group
(Pc-G) genes(Jurgens 1985). The Pc-G genes act in y mutations induced by chimeric mobile elements
lo-cated 69 bp upstream to the yellow transcription start normal development as repressors of BX-C and ANT-C
site (Georgievet al. 1997). They appeared against the
genes (Struhl1981;Duraet al. 1985;Duraand
Ing-background of the y2 allele caused by an insertion of
ham 1988). Several Pc-G members were found to be
gypsy at2700 bp position (Geyeret al. 1986;Parkhurst
associated in a multiprotein complex (Franke et al.
andCorces1986). The region of gypsy responsible for 1992;Rastelliet al. 1993;Muller1995;Plateroet al.
its mutagenic effect, su(Hw)-binding region, has the 1996;StruttandParo1997). Cis-regulatory elements
properties of an insulator: only yellow enhancers respon-necessary for maintaining the repressed state of
homeo-sible for the body and wing pigmentation located distally tic genes have been identified (MullerandBienz1991;
from the promoter are affected, whereas all other tissues
Simonet al. 1993;Chanet al. 1994;Chianget al. 1995)
show the wild-type coloration (GeyerandCorces1987, and designated as Pc-G response elements (PREs;
1992). Insertions inducing highly unstable y mutations
Simonet al. 1993).
consist of two identical copies of a defective P element, We have discovered a new P-element-induced
muta-1.2 kb long, and some genomic sequences located be-tion in the polyhomeotic (ph) gene, phP1, which represses
tween them. One such insertion, present in the y1s1
yellow expression in the yellow alleles containing
P-ele-allele, contains a 2.9-kb genomic sequence that includes ment insertions (Belenkayaet al. 1998). The phP1
muta-an enhmuta-ancer-like element activating yellow expression in tion is induced by a 1.2-kb P-element insertion into the
the body cuticle and wing blade (Georgievet al. 1997).
59 transcribed noncoding region of the ph gene. The
A large number of derivative alleles with deletions in
phP1 allele produces a chimeric protein, designated as
the internal part of the chimeric element or with P-Ph, that contains the DNA-binding domain of the
P-element rearrangements can be obtained from the P element and the Ph protein lacking 12 amino-terminal
y1s1allele.
In this article, we have characterized the interactions of the phP1mutation with the y1s1allele and its derivatives. Corresponding author: Pavel Georgiev, Institute of Gene Biology,
Rus-The inhibiting effect of phP1on yellow expression can be
sian Academy of Sciences, 34/5 Vavilov St. Moscow 117334, Russia.
E-mail: [email protected] suppressed either by the enhancer element located in
The removal of phP1was confirmed by Southern blot hybrid-the chimeric y1s1 insertion or by the yellow wing and
ization. body enhancers. However, an increase of P-element
Combinations of autosome Su(y) mutations with y*phP1were copy number either in cis or in trans strongly elevates obtained and analyzed according to the following scheme: F
0: the phP1-mediated repression that overcomes the effect
/y*phP1/FM43?y1s1phP1; Su(y)/CyO or Su(y)/TM3,Sb; analysis
of the y phenotype in y*phP1/Y; Su(y)/1males. of the enhancers. We have also studied the effect of
To study the eye phenotype on the phP1 background we
phP1on P-element-induced mutations in the singed locus,
constructed the yphP1w line. The su(Hw)2/su(Hw)vmutations which is expressed at the same middle pupal stage as
in the su(Hw) gene were combined with the y*phP1mutations
yellow during the formation of adult bristles and hairs, as described previously (GeorgievandKozycina1996). The
and in the female germline during oogenesis (Pater- su(Hw)v mutation is deletion of the su(Hw) gene, whereas
su(Hw)2is a strong mutation induced by jockey insertion in the
son andO’Hare1991). Unexpectedly, the phP1
muta-first intron of the su(Hw) gene (Harrisonet al. 1993).
tion does not influence the phenotype of sn mutations
DNA manipulations:DNA from adult flies was isolated using induced even by double P-element insertions.
the protocol described inAshburner(1989). The genomic During this work, we discovered a number of P-ele- DNA was digested with restriction enzymes according to the ment-induced mutations designated as Su(y) that par- supplier’s instructions and separated in standard agarose gels (Sambrooket al. 1989). The DNA was transferred to Hybond
tially or completely suppress the inhibitory effect of phP1
N1 membranes and probed according to the supplier’s in-on the y1s1allele and its derivatives. The effect of Su(y)
structions. The DNA fragments used as the probes were sepa-mutations results in a high level of P-element
expres-rated in agarose gels and purified using Gene Clean II (BIO sion. The truncated transposase competes with the chi- 101, Vista, CA) according to the supplier’s instructions. meric P-Ph protein for the same binding sites on The genomic DNAs were subjected to PCR to amplify
se-quences from the derivative alleles (Saikiet al. 1985;Mullis
P elements.
andFaloona1987). The primers used in DNA amplification were as follows: ATGCATTCTATGCACGAGCCTCC (y-1, 2409–2432 bp), CAGCGAAAGGTGATGTCTGACTC (y-2,
MATERIALS AND METHODS 2606–2628 bp), TCTGTGGACCGTGGCGCGGTAAC (y-3,
2898–2877 bp), and ACTTCCACTTACCATCACGCCAC (y-4,
Drosophila strains: All flies were maintained at 258 in a 3293–3270 bp) for the yellow gene according to sequences of standard yeast medium. Genetic symbols of the y alleles and the yellow gene presented inGeyeret al. (1986); CGCTGAG the origin of the y1s1allele and some of its derivatives were
GAACTCGAGAAAGGCC (c-1) and ACTGCGTGCTTCAAGC described in other articles (Georgievet al. 1992, 1997). The TTCTACC (c-2) for the internal genomic insertion of the
w;Sb P[ry1D2-3] (99B) e/TM6,e stock, providing a stable source y1s1allele; AAGCTAACAGCGCTGTTGGCTAG (sn-1, 875–898
of transposase (Robertsonet al. 1988), was obtained from bp) and ACACCCTTCACTTCGCCAATTCG (sn-2, 1632–1609
the Bloomington Stock Center (Bloomington, IN). Hereafter bp) for the singed gene according sequences of singed gene
P[ry1D2-3](99B) is referred to asD2-3. The rest of the Drosoph- presented inPatersonandO’Hare(1991); and CTCTCAA
ila melanogaster strains and mutations are described inLinds- CAAGCAAACGTGCACTG (p-1, 89–66 bp), CGTCCGCACA
leyand Zimm(1992). The origin and structure of the phP1
CAACCTTTCCTCTC (p-2, 108–85 bp), CTCAATACGACACT strain are described in another article (Belenkayaet al. 1998). CAGAATACT (p-3, 2802–2824 bp); and ATACGTTAAGTG
Genetic crosses:To induce mutagenesis, y1s1 phP1females
GATGTCTCTTG (p-4, 2853–2875 bp) for the P-element ac-were crossed to w; SbD2-3 e/TM6,e males to produce dysgenic cording to sequences of the P element presented inO’Hare males of the y1s1phP1/Y; SbD2-3 e/TM6,e/1genotype. In each
andRubin(1983). The amplifications were performed with vial,D2-3 males were mated to 10–12 C(1)RM,yf females. The a program including 1 cycle of 3 min at 948, 30 cycles of 1 min F2 progeny was analyzed for mutagenesis. All males with a at 948, 1 min at 648–688 (the temperature depended on the
new y phenotype were individually mated to virgin C(1)RM,yf primers), and 2 min at 728and, the last, 4 min at 728. females and their phenotype was examined in the next genera- The PCR products were directly sequenced using a Seque-tion. Determination of the yellow phenotype was performed nase II DNA sequencing kit for PCR product (Amersham, as previously described (Georgievet al. 1997). Arlington Heights, IL) according to manufacturer’s
instruc-Combinations of phP1with different y alleles were obtained
tions.
according to the following scheme: Northern blot hybridization:Total RNA was isolated by
ho-mogenization in 4mguanidine isothiocyanate, 0.2% N-lauryl F0: /y1scD1phP1wa/y1scD1phP1wa 3 ?y*/Y, where y* is any y
sarcosine, 150 mmmercaptoethanol, 12.5 mmEDTA, and 50 allele used;
mmtris-hydrochloride, pH 7.5, with subsequent phenol extrac-F1:/y1scD1phP1wa/y*3?y1scD1phP1wa/Y;
tion and ethanol precipitation (Parkhurstet al. 1988). Poly
F2: Selection of y*phP1wamales (sc1phenotype and orange
(A)1RNA was selected by chromatography on oligo(dT) cellu-eyes).
lose. Northern blot analysis was carried out as described by Parkhurstet al. (1988).
The introduction of phP1was confirmed by Southern blot
Construction ofP{w1,DP-yellow} andP-element
transforma-hybridization with labeled probes from the ph gene. The wa
tion: The structure and expression of the yellow gene were (1-1.5) mutation was used as the closest marker for the ph
described earlier (Geyeret al. 1986;GeyerandCorces1987). gene (1-0.5).
The 10-kb fragment including the yellow gene and its regula-The removal of phP1from a X chromosome with a y* allele
tory 3-kb region was provided by Dr. P. Geyer. The regulatory was performed according to the following scheme:
SalI/BamHI yellow region (3 kb) was subcloned in pGEM7
restricted by BamHI and XhoI. The XhoI-ScaI fragment of F0:/y1scD1wa/y1scD1wa3?y*phP1/Y;
F1:/y1scD1wa/y*phP13?y1scD1wa/Y; P element was eluted from the gel and blunt ligated in the
pGEM7 vector. A plasmid with two ligated XhoI-ScaI fragments F2: Selection of y*wa males (sc1 phenotype and orange
XhoI-Figure1.—Schematic presentation of the derivatives obtained from the y1s1 allele and their interaction with the phP1and
su(Hw) mutations. The directions of transcription of the yellow gene and of the 1A gene are indicated by arrows. The P-element
sequences and their size are shown by boxes. The P-element orientation is indicated by an arrow. 1A-En is the enhancer located in the chimeric element. Thin lines show the deletion in the 2.9-kb genomic sequence of chimeric mobile element in the y2s10-2
allele. Sequences remaining after P-element excision are indicated below a corresponding P element. The structure of y2s2 is
shown in the lower part of the diagram. Restriction enzymes are as follows: B, BamHI; H, HindIII; P, PstI; R, EcoRI; X, XhoI. The total number of black and white circles indicates the level of pigmentation in ph1flies in 1, body; 2, wings; 3, thoracic bristles; 4, leg bristles; 5, wing bristles; and 6, abdominal bristles. The number of white circles shows the level of pigmentation in flies with the phP1mutation. Thus the number of black circles shows the effect of the phP1mutation. One circle represents one point
as in Table 1. The effect of the phP1mutation was also studied on the su(Hw)2and su(Hw)1backgrounds. The su(Hw)2background
corresponds to the trans-heterozygous su(Hw)v/su(Hw)2.
ScaI fragment was cut out, blunt ligated, and cloned in the yellow1} were made homozygous and their copy number was
determined by Southern blot analysis using the yellow and white regulatory SalI/BamHI yellow region (3 kb) in the KpnI site
(Figure 1). One of the obtained clones was sequenced to sequences as probes. confirm that no unwanted changes in the yellow sequence had
taken place. The 3-kb yellow fragment cut out by BamHI and
XbaI was subsequently cloned in the BamHI and XbaI sites of RESULTS
the CaSpeR3 plasmid with a 5-kb coding region of the yellow
Cis-elements influencing the repression effect of the
gene (Figure 1). The new construct was denoted as P {w1,D
P-yellow}, where w1indicates a miniwhite gene included into the phP1mutation onyellowexpression:In a previous article
vector andDP-yellow indicates a complete yellow gene with an (Belenkayaet al. 1998), we found that the phP1
mutation insertion of a small part of the P element. The control con- completely inhibited yellow expression in y2s alleles
in-struct P{w1, yellow1} has an unchanged structure of yellow
se-duced by two copies of P element inserted 69 bp up-quences. This construct was obtained by direct cloning of
stream of the yellow promoter and mildly inhibited it in the 10-kb fragment including the yellow gene in the CaSpeR3
plasmid. y2salleles induced by a single P-element copy at the same
DNAs of the P {w1,DP-yellow} and P {w1, yellow1} constructs position. In addition, all described y alleles contained and of a P element with a defective inverted repeat used as a a gypsy insertion at2700 bp position. The su(Hw) insula-transposase source, P25.7wc (Kares andRubin 1984), were
tor located in the 59 regulatory region of gypsy blocks injected into y2ac2w67cpreblastoderm embryos as described
the yellow wing blade and body enhancers. As a result, previously (Rubin and Spradling 1982; Spradling and
Rubin1982). Survivors were crossed to y2ac2w67c flies and the y alleles display y2phenotype (yellow color of body
transgenic flies were separated on the basis of eye color. Chro- cuticle and wings).
mosome assignments of the various transgene insertions were In this article, we studied the y1s1allele (Georgievet
made by crossing the transformants with y2ac2w1118balancer
al. 1997) that contains, in addition to a pair of
1.2-stock containing dominant markers: In(2RL),CyO for the
sec-kb P elements, a duplicated 2.9-sec-kb genomic sequence ond chromosome and In(3LR)TM3,Sb for the third
designated as P 1 and P 2 depending on whether they the P 1 element in the head-to-head orientation (Figure 1). Another y allele, y1sA12, had P 2-element duplication were located distally or proximally with respect to the
yellow promoter. The 2.9-kb genomic sequence origi- also in the head-to-head orientation. Both y1s alleles had the same strong mutant phenotype in combination nated from the 1A region of the X chromosome. An
efficiently transcribed putative gene designated as the with the phP1mutation. Thus, only the number of P-ele-ment copies at the yellow locus was important for the
en-1A gene was found in the middle of the 2.9-kb sequence
(Figure 1). y1s1 flies have wild-type level pigmentation hancement of the effect of the phP1mutation, but not their localization in the chimeric element.
in body cuticle and wing blade in contrast to flies with
the original y2 allele, in which the body and wing en- The y1s22 allele was induced by an inversion of the central 2.9-kb region without P-element deletion (
Geor-hancers are blocked by su(Hw) insulators. A number
of derivatives with y2phenotype were obtained from the giev et al. 1997). y1s22 flies had the wild-type level of pigmentation indicating the 1A enhancer to act in an
y1s1. All of them had deletions of various size in the
2.9-kb genomic sequence of chimeric mobile element orientation-independent fashion. However, the phP1 mu-tation had a stronger effect on the y1s22alleles than on (Georgievet al. 1997;I. BirukovaandP. Georgiev,
unpublished results). The mapping of these deletions the original y1s1allele (Figure 1). Thus, the orientation of the 2.9-kb insertion influences the level of phP1 -medi-allowed us to localize the enhancer element between
the 1A gene and P 2 element (Figure 1). ated repression. Possibly, the insertion of the 1A pro-moter between the 1A enhancer and the yellow propro-moter The phP1mutation only partially repressed yellow
tran-scription in the y1s1allele: the pigmentation of the body, facilitates inhibition by the P-Ph protein of the 1A en-hancer action on yellow transcription. Another explana-wings, thoracic, and leg bristles was decreased (Figure
1). Thus, the presence of the genomic insertion pos- tion is that the inversion brings a distance-dependent negative element cooperating with phP1into close vicin-sessing the 1A enhancer reduces the effect of the P-Ph
protein. ity to the yellow promoter. However, the y2s2 derivative,
which had, in addition to the P 2 element, a 410-bp To further analyze the role of different cis-regulatory
elements in formation of repressive complex, we studied sequence just from the distal part of the 2.9-kb insertion (Figure 1), was affected by phP1to the same extent as y2s the y1s1 derivatives obtained after P-element
mobiliza-tion in the y1s1 phP1 and y1s1 strains. The structure of alleles with an insertion of the P 2 element alone (Fig-ures 2 and 3).
insertions in some of these derivatives was described in
a previous article (Georgievet al. 1997), while for new To further study the dependence of phP1repression on the orientation of the chimeric element, we mobilized P
y alleles, it was determined using Southern blot analysis
and the PCR cloning and sequencing technique. The elements in the y1s22phP1strain. As a result, we obtained a y1s32derivative characterized by a variegated pigmenta-presence of nonmodified P element in the phP1 allele
was verified by Southern blot analysis as described by tion of bristles in combination with phP1(Figure 1). The
y1s32allele was generated by an excision of P 2 element,
Belenkayaet al. (1998). All derivatives may be divided
for several classes because of their structure and the leaving only 14 and 18 bp of the terminal repeats. Thus, the phP1 mutation can inhibit yellow expression in the interaction with phP1. To simplify the presentation, we
described here only one representative y allele for each presence of just a single distal P element located at a 3-kb distance from the yellow promoter if an inversion class of derivatives, although several independently
ob-tained y alleles with the identical or similar structure of the 2.9-kb sequence takes place.
The upstream region of the yellow gene contains the have been analyzed in each case.
The y2s10-2allele was obtained as a result of 1A enhancer yellow wing and body enhancers, which are blocked due to the presence of the su(Hw) insulator (Figure 2). To deletion (Georgiev et al. 1997). The combination of
y2s10-2
with the phP1
mutation led to a complete inhibition study the role of yellow enhancers in the phP1
-mediated repression, we inactivated the su(Hw) insulator, cross-of yellow expression (Figure 1). Thus, the 1A enhancer
is responsible for the reduction of the P-Ph protein ing the combination of y and phP1 mutations into
su(Hw)2background. The su(Hw)v/su(Hw)2 transhetero-effect.
The y1s27 allele was generated by excision of P1 ele- zygote completely suppressed the mutant y phenotype of flies carrying the combination of the phP1 mutation ment. PCR cloning and sequencing showed that only
15–17 bp from each terminus were retained (Figure 1). with either y1s1or y1s32mutation (Figure 1). Thus, the
yellow body and wing enhancers are also able to suppress
The phP1mutation failed to influence the phenotype of
y1s27 flies, suggesting that the 1A enhancer completely a weak phP1-mediated repression.
The su(Hw)v/su(Hw)2transheterozygote partially sup-overcomes phP1-mediated repression, if only one P
ele-ment is present at the yellow locus. pressed the phP1-mediated repression in the y1sA11, y1sA11, and y1s22 alleles. This result shows that 1A and yellow The y1salleles associated with duplication of any P
element were characterized by an almost complete inac- enhancers cooperatively protect yellow expression from the phP1repression. However, the yellow enhancers fail tivation of yellow expression by the phP1mutation (Figure
Figure2.—Structure of the
yellow locus in wild-type and
mutant y alleles and in the
P{w1, DP-yellow} construct.
Ex-ons of the yellow gene and the direction of transcription are indicated by arrows. The exons are separated by one intron. Transcriptional enhancers are indicated by circles. En-w, en-hancer for the wing blade; En-b, enhancer for the body cuticle; En-br, enhancer for bristles. Insertions responsible for different alleles are repre-sented by triangles. The gypsy element is inserted in the 59 region of the yellow gene in the
y2allele and its derivatives. The
arrows flanking gypsy are long terminal repeats (LTRs). They indicate the direction of gypsy transcription. Deletion of DNA sequences in the y1s8 allele is
represented by a horizontal line. The su(Hw) binding site is represented by an ovoid lo-cated in the 59 transcribed-untranslated region of gypsy. Re-striction enzymes are as follows: H, HindIII; G, BglII; X, XhoI; K, KpnI; B, BamHI; S, ScaI. The primers used for PCR cloning are indicated by arrowheads. The P{w1,DP-yellow} indicates the site of insertion (KpnI) of the double XhoI-ScaI fragments of P element.
In the lower part of the diagram P{w1, DP-yellow} is shown schematically with the wing (En-w) and body (En-b) enhancers, P-element sequences (P), and miniwhite and yellow fragments indicated. The arrow shows the direction of transcription of the miniwhite and yellow genes. The miniwhite gene is not shown in scale.
su(Hw)2flies (Figure 1). Thus, the yellow enhancers have additional binding site for a truncated transposase (Lee
et al. 1996).
weaker effect on the phP1-mediated inhibition of
tran-scription than the 1A enhancer in the y1s1allele. Another derivative, y2s42allele, which had only 41 bp of the 39end and 852 bp of the 59end, was affected by
The P-element sequences responsible forphP1
-medi-ated repression:In an earlier article (Belenkayaet al. phP1: the notum and leg bristles acquired a variegated
mutant phenotype (Figure 3). The P element in the y2s42 1998), we demonstrated the absence of phP1effect if the
P-element deletion led to the loss of transposase-binding allele had a deletion of 39transposase-binding sites, but it differed from the y2s34allele by the presence of a part sites. Here we have further studied the role of different
P-element sequences. From this, the derivatives of y2s14 of the central region of the defective P element between 767 and 852 bp. The latter included the breakpoint of and y1s1 alleles generated by partial deletions of the
P elements were characterized. The breakpoints of the the deletion and the filler sequence TAGCTACAAA, suggesting the presence of additional binding sites for deletions in the P elements were cloned by PCR and
sequenced. The effect of the phP1
mutation was studied the chimeric P-Ph protein in this region.
To identify the role of the central region of the in y*phP1/Y males and in y*/y1
phP1heterozygous females. As was shown previously, the phP1 mutation mildly
P element in the phP1repression, we prepared the P {w1,
DP-yellow} construct containing two 179-bp XhoI-ScaI
re-repressed yellow expression in the y2s14allele containing
a single P-element copy (Belenkaya et al. 1998). Its striction fragments of the 1.2-kb P element (the frag-ment between 730 and 909 bp of the latter) at the derivative, y2s34allele, was induced by a P element with
an internal deletion. The breakpoints of the deletion position of2340 bp from the yellow transcription start site (Figure 2). Seven independent transformants were were mapped at distances of 767 bp from the 59terminus
and 93 bp from the 39terminus; i.e., both termini con- obtained, possessing a single insertion as confirmed by Southern blot analysis. All of them displayed wild-type tained transposase-binding regions, but the 11-bp
in-verted repeat at the 39terminus was deleted (Figure 3). levels of yellow expression. The combination of the phP1 mutation with the P {w1,DP-yellow} construct in five cases
The phP1 mutation had no visible effect on the yellow
expression in combination with y2s34. The suppression led to mosaic pigmentation of bristles, while the pigmen-tation of the body and wings was not changed (data of the phP1 effect may be explained by the previous
Figure 3.—Schematic pre-sentation of the phP1effect on
P-element deletion derivatives
in y alleles. Above is a structure of the 1.2-kb P element. The double line in the center of this box indicates the breakpoints of the internal deletion. IR, 31-bp inverted repeat; TPS, trans-posase-binding site. Below are the P-element sequences pres-ent in the alleles indicated at the left. The punctuated boxes represent P-element sequences remaining in the deletion de-rivatives; the black boxes repre-sent the complete 1.2-kb copy of the P element. The numbers below the boxes are the num-bers of base pairs remaining in the P-element sequences from the corresponding end to the break point. The arrows show the direction of P element in the corresponding alleles. The total number of black and white circles in each lane indicates the level of pigmentation of y*phP1females in 1, body; 2, wings; 3, thoracic bristles; 4, leg bristles;
5, wing bristles; and 6, abdominal bristles. The number of white circles shows the level of pigmentation in the y*phP1females
(the number of black circles shows the effect of the phP1mutation). The su(Hw)2/su(Hw)vmutations in the su(Hw) gene were
combined with the y*phP1to produce su(Hw)2background.
heterozygous and homozygous states of the P {w1,DP- terminal 82 bp. This deletion includes the 59-terminal binding sites for the P-element transposase or the P-Ph
yellow} construct. As a control, we used P {w1, yellow1}
transposon, which contained the unmodified yellow protein. To find a minimal region of the P 2 element that was sufficient for strong enhancement of the phP1 gene. Nine transgenic lines carrying P {w1, yellow1}
transposon were obtained. The y phenotype of flies from repressive effect, we analyzed the structure of two y2s mutations, derivatives of y1s1, which in combination with these lines was not changed on a phP1background. This
result confirms the role of the central XhoI-ScaI region phP1led to a complete inactivation of yellow expression. Both y alleles contained the unchanged P1 element. of the 1.2-kb P element in the phP1-mediated repression
of the yellow expression. The y2s25allele contained only 39 bp of the 59terminal sequence and 349 bp of the 39terminus of the P 2 ele-We also compared eye pigmentation in either the
presence or absence of the phP1mutation. The miniwhite ment. The y2s26 allele contained just 153 bp of the 39 terminal sequence of the P 2 element (Figure 3). How-gene is very sensitive to repression by Pc-G complex.
For example, a 661-bp PRE core fragment from the Ubx ever the phP1 mutation still completely repressed the
yellow expression in these alleles. Thus, the addition of
gene, inserted in a CaSpeR3 vector, generally causes
variegated miniwhite gene expression in 50–60% of the the P-element transposase binding sites and the 11-bp inverted repeat to the single P element is sufficient for lines (Chanet al. 1994). We found that only one of six
lines carrying P {w1, DP-yellow} transposon in heterozy- strong enhancement of the phP1-mediated repression. To make the difference between strong and weak gous state displayed a weak decrease of eye
pigmenta-tion after cross to a phP1
w background. In the homozy- phP1
-mediated repression more clear, we introduced the combinations of phP1with the y mutations into a su(Hw)2 gous state of the P {w1,DP-yellow} transposon, flies from
four of six lines showed decrease of eye pigmentation background (Figure 3). The su(Hw)2/su(Hw)v transhet-erozygote completely suppressed the mutant yellow phe-and a weak variegation characteristic for Pc-G repression
of white expression (data not shown). Thus, the ends of notype of flies carrying the combination of the phP1 mutation with either y2s14or y2s16 mutation induced by
P elements in the presence of phP1 have only a weak
repression potential compared to natural PREs. single P-element insertion. Thus, the yellow body and wing enhancers are able to suppress a weak phP1 -medi-As shown previously (Belenkayaet al. 1998), the phP1
mutation mildly repressed yellow expression in the y2s14 ated repression. On the contrary, the su(Hw)2/su(Hw)v transheterozygote only weakly influences the phenotype and y2s16 alleles containing a single P-element copy,
whereas it completely inhibited yellow expression in the of flies carrying the phP1mutation with either y2s11, y2s26, or y2s25alleles. Thus, the 39sequences of the P element
y2s11 allele induced by an insertion of two P-element
copies in the head-to-head orientation (Figures 2 and can strongly enhance the phP1-mediated repression.
Pairing-dependent repression of yellow expression
3). The y2s11 allele contains the P1 element with
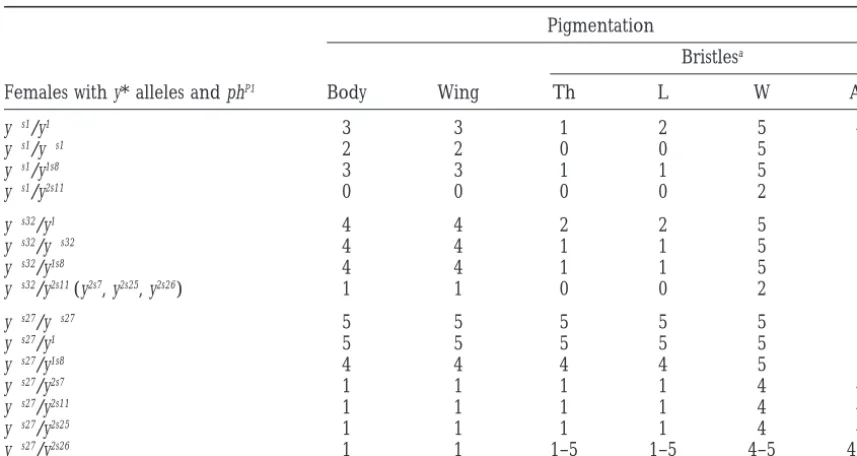
muta-TABLE 1
phP1-mediatedtransrepression of transcription in females heterozygous foryalleles
Pigmentation
Bristlesa
Females with y* alleles and phP1 Body Wing Th L W Ab
y1s1/y1 3 3 1 2 5 4
y1s1/y1s1 2 2 0 0 5 2
y1s1/y1s8 3 3 1 1 5 3
y1s1/y2s11 0 0 0 0 2 1
y1s32/y1 4 4 2 2 5 5
y1s32/y1s32 4 4 1 1 5 5
y1s32/y1s8 4 4 1 1 5 5
y1s32/y2s11(y2s7, y2s25, y2s26) 1 1 0 0 2 1
y1s27/y1s27 5 5 5 5 5 5
y1s27/y1 5 5 5 5 5 5
y1s27/y1s8 4 4 4 4 5 5
y1s27/y2s7 1 1 1 1 4 4
y1s27/y2s11 1 1 1 1 4 4
y1s27/y2s25 1 1 1 1 4 4
y1s27/y2s26 1 1 1–5 1–5 4–5 4–5
The figures show the level of pigmentation estimated visually in 3- to 5-day-old females developing at 258. The level of pigmentation was evaluated on a scale from 0 to 5. Zero is the pigmentation of y1flies and 5 is
the pigmentation of y1flies. Two numbers indicate the variable pigmentation of bristles. Th, thoracic; L, leg; W, wing; Ab, abdominal.
tion was stronger in homozygous y1s1 females than in Thus, a single P-element copy failed to induce a strong pairing-dependent repression by the phP1 mutation. In
y1s1 males or heterozygous y1/y1s1 females (Table 1),
where the y1allele was generated by a point mutation contrast, the y2s7, y2s11, and y2s25alleles induced a strong pairing-dependent repression (Table 1). These y2salleles in the coding region of the yellow gene (Geyer et al.
1990). Thus, the presence of two P-element copies in were induced either by two P elements or by the P 2 element and the 39terminus of the P1 element (Figures each homologue resulted in the enhancement of
silenc-ing. To study the transinteraction in detail we used well- 2 and 3). However, the effect of phP1 was weaker in heterozygous females with the y2s26allele. The y2s26allele studied y alleles in combination with phP1. As has been
shown above, the y1s32allele is associated with a deletion differed from the y2s25 allele only by a deletion of the central region of the P1 element, further confirming of the P 2 element proximal to the yellow gene and the
y1s27 allele—with a deletion of the distal P1 element. the role of this part of the 1.2-kb P element in the phP1 -mediated repression of yellow transcription.
The phP1mutation influenced almost to the same extent
on heterozygous y1/y1s32 and homozygous y1s32/y1s32fe- Su(y)mutations:In the progeny obtained after P-ele-ment mobilization in y1s1phP1flies, we could frequently males and had no effect on the y1s27allele (Table 1).
To further study the role of P-element sequences in observe the appearance of autosomal suppressors of
phP1-mediated repression. These mutations, designated the formation of a phP1-dependent repression complex,
we studied the phenotype of heterozygotes for the above as Su(y), were widely spread on the X, second, and third chromosomes and displayed different levels of the sup-mentioned y1s1, y1s32, or y1s27 alleles with the y alleles
possessing one, one and one-half, or two P-element cop- pression effect. Eleven Su(y) mutations were lethal in the homozygous state, 9 possessed normal or decreased ies in combination with phP1
. For a y allele with a single
P-element copy, we used the y1s8mutation (Belenkaya viability, and 2 were associated with female sterility. One of the Su(y) mutations appeared simultaneously with a
et al. 1998) associated with a deletion of the yellow
pro-moter, but still containing a single P-element copy (Fig- mutation in the ebony (e) locus, and the recombination analysis showed them to be one and the same mutation. ure 2). This y allele displays the same y-null phenotype
as the combination of phP1 and y2s alleles containing Another strong Su(y) mutation was obtained in the singed locus (sneP1).
more than one P-element copy.
The combination of y1s8with either y1s1, y1s32, or y1s27 For further study, we selected three Su(y) alleles,
Su(y)P31(third chromosome), Su(y)P22 (second chromo-did not significantly increase the effect of phP1as
The most plausible explanation was that Su(y) muta-tions were induced by P-element insermuta-tions into regula-tory or transcribed regions of different genes that led to a high level of transcription of the defective P ele-ments and to the accumulation of truncated transposase that might suppress the mutant phenotype of the y al-leles competing with the P-Ph protein for binding sites. To test this hypothesis, we performed a Northern blot analysis of mRNA from the Su(y) and control strains using as a probe the P-element HindIII-XhoI fragment that included the sequences encoding the DNA-binding domain of transposase (AndrewsandGloor1995;Lee
et al. 1996). A high level of mRNA hybridizing with
the P-element sequences was detected only in the Su(y) strains (Figure 4). The highest level of P-element tran-scripts was found in the Su(y)P31strain at the larval, pupal, and adult stages of development. The Su(y)P22strain had a lower but still rather high level of P-element transcripts at all tested stages of development. In the sneP1strain, a high level of P-element transcripts was detected only at the early pupal stage of development, when the singed gene was actively transcribed (Paterson andO’Hare
1991).
Molecular study of thesneP1 allele:To gain a better
insight into the mechanism of Su(y) action, we used for molecular analysis the sneP1 mutation in the well-Figure4.—The effect of Su(y) mutations on the inhibitory
characterized singed locus (Roihaet al. 1988;Paterson effect of the phP1. (A) The effect of Su(y)P31, Su(y)P22, and sneP1
and O’Hare 1991). The sneP1 mutation had a strong
mutations on the y* phP1/y1phP1females. The total number of
mutant phenotype. Southern blot hybridization re-circles in each lane indicates the level of pigmentation of y*
phP1/y1phP1females in the presence of the Su(y) mutations: 1,
vealed anz1.0-kb insertion localized between the EcoRI the body; 2, wings; 3, thoracic bristles; 4, leg bristles; 5, wing and SalI restriction sites of the DNA segment (Figure bristles; and 6, abdominal bristles. The number of black circles
5). This insertion was within the 700-bp singed “hot spot” indicates the level of yellow expression in phP1flies in the
ab-region (Hawley et al. 1988; Roiha et al. 1988). For
sence of Su(y) mutations. The dotted circles indicate variations
cloning of the P-element insertion, the oligonucleotides of yellow expression. Upper lines show the level of
pigmenta-tion in ph1flies; lower lines show the level of yellow expression were designed from both sides of the insertion (Figure in y-null mutation. (B) Northern blot analysis of transcripts 5). The PCR-amplified fragment was directly sequenced. hybridizing with P-element HindIII-XhoI probe in Su(y) strains.
The sneP1insertion was shown to be a 1180-bp internally
The mRNA was isolated from the y2s14(1, 8, 12, 16, 20), y2s14;
deleted P element with a structure identical to that
Su(y)P31/TM3,Sb (3, 6, 10, 14, 18), y2s14; Su(y)P22/CyO (4, 5, 9,
described for the P element responsible for superunsta-13, 17), and y2s14 sneP1 (2, 7, 11, 15, 19) strains at different
stages of development: first instar larvae (1–4), second instar ble y alleles and phP1mutation. Based on the published
larvae (5–8), third instar larvae (9–12), early pupae (13–16), sequences of a full-length P element (O’Hare and and late pupae (17–20). The HindIII-XhoI probe was
hybrid-Rubin1983), this sneP1P element contained an internal
ized to the 1.0-kb major transcript and some other minor
deletion from 830 to 2560 bp and a filler sequence transcripts of higher molecular weight.
TAGCTACAAA at the break point. Interestingly, as in the case of highly unstable y mutations, the P-element insertion at the singed locus was associated with a dele-we studied the phenotype of females with the y phP1/y*
phP1; Su(y)/1genotype, where y* was one of the y alleles tion: 4 bp (CGCG) at the target site were lost, and no 8 bp duplication typical of P-element insertions was described above (Figure 4). All three Su(y) mutations
completely suppressed the effect of phP1on the y2s14and observed. The P element in the sneP1allele is localized in the transcribed but not translated part of the singed
y1s1alleles. The Su(y)P31mutation also almost completely
suppressed the effect of phP1on the y alleles induced by genez80 bp downstream from the signal of transcrip-tion initiatranscrip-tion (PatersonandO’Hare1991;Burkeand a chimeric element containing even three P elements
(y1sA8and y1sA22) or an inversion of the chimeric element Kadonaga1996).
To prove the responsibility of sneP1for the Su(y) effect, (y1s12). Su(y)P22 and sneP1 had only a weak suppressive
effect in these combinations. However, the Su(y) muta- we studied revertants induced by crossing with D2-3. Five sn1Prevertants were obtained and shown by South-tions did not have any visible effect on the combination
addition to this phenotype, many strong sn mutations lead to female sterility (Lindsley andZimm 1992).
The P element in the above described sn mutations was inserted at a distance of ,50 bp from the core region of the downstream promoter element (DPE) of the singed gene (Paterson andO’Hare 1991; Burke
andKadonaga1996). However, the phP1mutation did not increase the visible mutant phenotype of the sn mutations induced by one (sne, sneP1, and sn1) or two (snw, snwP13, and snwP14) P elements (data not shown). This result could not be explained by the fact that sn mutations suppressed the phP1 effect, because such a weak suppressor as the snwmutation also was not modi-fied by phP1
. The phP1 mutation also did not influence the sn-phenotype in females, suggesting the absence of the pairing effect.
P-element-induced sn mutations did not significantly
Figure 5.—Structure of sn alleles. Start and direction of
influence the singed function in gonads: females
homo-singed transcription is indicated by an arrow. Insertions of P
zygous for a sn mutation were fertile. Only the sneP1
elements responsible for different sn alleles are represented
by triangles. The P-element sequences and their size are shown mutation reduced female fertility. The phP1 mutation
by boxes. The orientation of P elements are indicated by did not significantly increase the sterility of females with arrows. The sequence of the region of P-element insertions homozygous sn mutations.
is shown in the lower part of the figure. The duplicated eight nucleotides of the host DNA are underlined. The deleted cgcg
nucleotides are shown by lowercase letters. The primers used DISCUSSION
for PCR cloning are shown by arrowheads.
The properties ofP-element-mediated repression of
the yellow gene by the phP1 mutation: The chimeric
ously lost the ability to suppress the mutant phenotype P-Ph protein encoded by the phP1locus consists of the of the y2s14
phP1
combination (Figure 5). P-element transposase DNA-binding domain and an al-We also studied two snwPmutations, snwP13 and snwP14,
most full-length Ph protein (Belenkayaet al. 1998). It
obtained from sneP1as a result of P-element duplication
binds to the P-element sequences and leads to Pc-in the head-to-head orientation, as Pc-in the case of the G-dependent silencing in y alleles induced by P-element well-known snwmutation (Roiha et al. 1988). The snwP
insertions. Thus, against the phP1background the P-ele-mutations only slightly suppressed the y2s14phP1 mutant
ment sequences function as the known response PREs. phenotype, possibly due to a low level of P-element PREs were found in the regulatory regions of many
transcription (Figure 5). homeotic genes and are responsible for transcription
The P elements in the previously described snw
muta-repression (Pirrotta 1997). PREs were identified by tion (Engels1989) are located in the same region as their silencing effect on a reporter gene and by the in the sneP1mutation. The snwallele contains two P
ele-introduction of variegated or pairing-sensitive expres-ments of 1.15 and 0.95 kb inserted at the same site in sion of the white gene in transgenic flies (Fauvarque
the inverse orientation. Excision of the 0.95-kb P ele- and Dura 1993; Simon et al. 1993; Chanet al. 1994;
ment gives rise to sne, and excision of the 1.15-kb
Kassis1994). DNA fragments with PRE activity are
gen-P element, to sn1(Roihaet al. 1988). The snwand sn1
erally from several hundred to several thousand base mutations did not influence the effect of phP1. Only the pairs in length. They are composite in structure and
sne
allele had a weak suppressive effect (Figure 5). The can be dissected into multiple subfragments that display
P elements in sne and sneP1have the same orientation
a progressively weakening PRE activity. PREs have a and are inserted at a distance of 13 bp from each other strong cooperative effect on repression. For example, (Figure 5). The only difference between the P elements in the Ubx gene, where silencing is required to be strong is the deletion in the P element inserted in the sneallele
and stable, several weaker sites of Pc-G protein binding of the second region important for the protein-protein cooperate in establishing a strong silencing complex interaction that enhances DNA binding (Lee et al. (MullerandBienz1991;Pouxet al. 1996).
1996). This result shows that a strong DNA-binding abil- The y alleles used in this study have been generated ity is important for realization of the Su(y) effect. by an insertion of the 2.9-kb genomic DNA sequence
The phP1mutation does not affect singedexpression
from the 1A region of X chromosome located more
inP-element-inducedsnmutations:The singed gene of distally with respect to the yellow gene. The genomic
D. melanogaster plays a role in both somatic and germ duplication is flanked by two identical copies of a de-cells (LindsleyandZimm1992). The bristles and hairs leted 1.2-kb P element (Georgievet al. 1997). The 1A
for yellow activation in the body cuticle and wing blade tion can be partially suppressed by Su(y) mutations in-duced by a P-element insertion into regulatory or coding (Georgiev et al. 1997) compensating the yellow body
and wing enhancers blocked by the su(Hw) insulator. regions of different genes. The inserted 1.2-kb P ele-ment is transcribed at a high level and gives rise to a The appearance of a strong enhancer disturbs the
re-pressive effect of the P-Ph protein. Addition of the wing truncated transposase. For example, in the sneP1 muta-tion, the 1.2-kb P element is inserted into the 59 tran-and body enhancers by inactivation of the su(Hw)
func-tion further suppresses the silencing effect of the phP1 scribed noncoding region leading to an enhanced
P-element transcription. As shown before, mRNA
en-mutation. It has been shown that the assembly of a
silencing complex depends both on the strength of a coding truncated transposase was also formed as a result of the phP1mutation (Belenkaya et al. 1998) and this PRE site and on the transcriptional activity of the region
involved (Pirrotta1997). For example, the formation may partially decrease its effect.
The 1.2-kb P element has a deletion between 830 of a Pc-G complex and the binding of GAL4, which
activates transcription, are mutually exclusive and the and 2560 bp and resembles the previously described KP element (Blacket al. 1987). The DNA-binding domain
silencing state can be prevented by a strong activation
of transcription (Zink and Paro 1995; Cavalli and is located within the region of 98 amino-terminal amino acids (Rio1990; AndrewsandGloor1995). The KP
Paro1998).
The level of repression induced by phP1depends di- protein also contains two protein-protein interaction regions, and dimerization of the KP protein is essential rectly on the number of P elements but not on their
localization. The phP1-mediated repression may act at a for high-affinity DNA binding (Leeet al. 1996). A puta-tive leucine zipper is located between 101 and 122 distance from the yellow promoter. In the y1s32allele the
P element is located z3 kb from the yellow promoter amino acids of the KP protein (Rio1990;Andrewsand
Gloor 1995). The second protein-protein interaction and is separated from the latter by the 1A enhancer.
Still in this case, the phP1mutation inhibits yellow tran- region is present within a segment of 69 carboxy-termi-nal amino acids of the KP protein, i.e., still in the amino-scription. In this respect, the P elements act like PREs
that are responsible for the repressive state in the terminal part of the intact P element (Leeet al. 1996).
All these sequences are also present in the protein en-adjacent genome region in a cooperative manner
(OrlandoandParo1993;Pirrotta1997). coded by the 1.2-kb P element.
The KP protein binds to multiple sites on the P-ele-The phP1mutation induces only a weak repression in
y alleles containing a single P-element copy upstream ment termini with a higher affinity than the full-length transposase (Leeet al. 1996). The binding sites include
of the yellow gene. Thus, one copy of P element is not
sufficient for the formation of a strong repression com- the high-affinity transposase binding sites, an 11-bp transpositional enhancer, and the terminal 31-bp in-plex. Addition to a single copy of the 1.2-kb P element
of 153-bp 39-terminal P-element sequences strongly en- verted repeats. Both protein-protein interaction regions are important for this binding. The 1.15-kb P element hances phP1-mediated repression, leading to a complete
inactivation of yellow expression. As expected, the dele- in sne, a derivative of the snwmutation (Roihaet al. 1988;
Engels1989), is inserted in almost the same place and tion of transposase-binding sites or 11-bp inverted
re-peats reduces the phP1-mediated repression. Unexpect- has the same orientation as the P element in sneP1. The protein product of the 1.15-kb P element contains the edly, the internal region of the 1.2-kb P element is also
important for the phP1-mediated repression that suggests DNA-binding domain and a leucine zipper, but not the second protein-protein interaction region (Roihaet al.
the presence of a potential site(s) for binding of the
chimeric P-Ph protein. 1988;Leeet al. 1996). This explains why the sneP1 muta-tion strongly suppresses the phP1inhibitory effect, while PRE-containing transposons often show a dramatic
enhancement of silencing when a fly is homozygous for sne
does not. TheD2-3 construct generating a full length transposase has a very weak suppression effect on the a transposon insertion, indicating that the
homolo-gously paired PREs interact to produce a more stable phP1 mutation (
T. Belenkaya, unpublished results). This may also be explained by its lower affinity to the and more repressive Pc-G complex (Fauvarque and
Dura 1993; Chan et al. 1994; Kassis 1994). We did P-element DNA.
The defective transposase with the DNA-binding do-not find any pairing-dependent repression when both
paired y alleles contained a single P-element copy induc- main and two protein-protein interaction domains is most efficient for realization of the suppression effect. ing a weak or no phP1-mediated repression. A
pairing-dependent repression was found only if one of the y Thus, the suppression of phP1-induced inhibition is a result of the competition for binding sites on the alleles in heterozygous females induced a strong
repres-sion complex in the presence of phP1. P elements between the KP-like protein and the P-Ph protein.
TruncatedP-element transposase similar in structure
to the KP protein suppresses the inhibitory effect of the The presence of a strong enhancer in the chimeric
element helps the KP-like protein in blocking the
assem-chimeric P-Ph protein on the expression ofP
Ashburner, M.,1989 Drosophila. A Laboratory Manual. Cold Spring However, the Su(y) mutations, even expressing the
trun-Harbor Laboratory Press, Cold Spring trun-Harbor, NY.
cated P-element transposase at high levels at all stages of Belenkaya, T., A. Soldatov, E. Nabirochkina, I. Birjukova, S. Georgievaet al., 1998 P element insertion at the polyhomeotic Drosophila development, fail to affect the phP1-mediated
gene leads to formation of a novel chimeric protein that
nega-inhibition of y2salleles induced by a double P element
tively regulates yellow expression in P element-induced alleles of
in the absence of the 1A enhancer. The formation of Drosophila melanogaster. Genetics 150: 687–697.
Black, D. M., M. S. Jackson, M. G. KidwellandG. A. Dover,1987
a strong repression complex prevents binding of the KP
KP elements repress P-induced hybrid dysgenesis in Drosophila
protein to transposase-binding sites on the P elements.
melanogaster. EMBO J. 6: 4125–4135.
Thus, the general low level of transcription activity facili- Burke, T. W.,andJ. T. Kadonaga,1996 Drosophila TFIID binds to
a conserved down stream basal promoter element that is present
tates P-Ph protein binding to DNA and the formation
in many TATA-box-deficient promoters. Genes Dev. 10: 711–724.
of a repressive complex.
Cavalli, G.,andR. Paro,1998 The Drosophila Fab-7 chromosomal
singed expression is insensitive to the phP1mutation
element conveys epigenetic inheritance during mitosis and meio-sis. Cell 93: 505–518.
in the P-element-induced mutations: The singed gene
Chan, C.-S., L. RastelliandV. Pirrotta,1994 A Polycomb
re-plays a role in somatic cells during the formation of
sponse element in the Ubx gene that determines an epigenetically
adult bristles and hairs, and in the female germline inherited state of repression. EMBO J. 13: 2553–2564. Chiang, A., M. B. O’Connor, R. Paro, J. SimonandB. Bender,1995
during oogenesis (Paterson andO’hare1991).
Dur-Discrete Polycomb binding sites in each parasegmental domain of
ing metamorphosis, a single 3.6-kb RNA is transcribed,
the bithorax complex. Development 121: 1681–1689.
but in ovaries two additional transcripts, 3.3- and 3.0- Dura, J.-M.,andP. Ingham,1988 Tissue- and stage-specific control
of homeotic and segmentation gene expression in Drosophila
em-kb RNAs, appear. The mRNAs differ only in the length
bryos by the polyhomeotic gene. Development 103: 733–741.
of the 39-untranslated region and a single gene product
Dura, J.-M., H. W. BrockandP. Santamaria,1985 polyhomeotic : a is predicted for all of them. The singed gene contains a gene of Drosophila melanogaster required for correct expression of
segment identity. Mol. Gen. Genet. 198: 213–220.
downstream promoter element (DPE) that is located
Engels, W. R.,1989 P elements in Drosophila melanogaster, pp. 437– only 50 bp upstream from the sites of insertions of P
484 in Mobile DNA, edited by D. E. Berg and M. M. Howe. elements in the sn mutations (PatersonandO’Hare American Society for Microbiology, Washington, DC.
Fauvarque, M.-O., and J.-M. Dura, 1993 polyhomeotic regulatory 1991;BurkeandKadonaga1996). However, the phP1
sequences induce developmental regulator-dependent
variega-mutation does not influence the bristle phenotype and
tion and targeted P-element insertions in Drosophila. Genes Dev.
female sterility of the sn mutants, even those induced 7:1508–1520.
Franke, A., M. Decamillis, D. Zink, N. Cheng, H. W. Brocket al., by a double P element. This result may be explained
1992 Polycomb and polyhomeotic are constituents of a
multi-either by a special organization of the singed promoter
meric protein complex in chromatin of Drosophila melanogaster.
making it insensitive to the P-Ph protein or by a structure EMBO J. 11: 2941–2950.
Georgiev, P.,andM. Kozycina,1996 Interaction between
muta-of chromatin in a vicinity muta-of the singed locus.
tions in the suppressor of Hairy wing and modifier of mdg4 genes of
As we have found here, the miniwhite expression in
Drosophila melanogaster affecting the phenotype of gypsy-induced transposons is weakly sensitive to the phP1mutation that
mutations. Genetics 142: 425–436.
Georgiev, P. G., V. A. Yelagin, E. M. BuffandN. P. Kolyagin, may be explained by a large distance between the white
1992 Properties of super unstable mutations in the Drosophila
promoter and P-element sequences in the transposon.
melanogaster yellow locus. Genetica 28: 98–107 (in Russian). However the effect of the phP1mutation is not restricted
Georgiev, P., T. Tikhomirova, V. Yelagin, T. Belenkaya, E. Gra-chevaet al., 1997 Insertions of hybrid P elements in the yellow
to the yellow gene. In another article (Soldatov et al.
gene of Drosophila cause a large variety of mutant phenotype.
1999), we described the strong phP1-mediated repression
Genetics 146: 583–594.
of TAFII40/e(y)1 expression in the e(y)P1allele that was Geyer, P. K.,andV. G. Corces,1987 Separate regulatory elements
are responsible for the complex pattern of tissue specific and
generated by P-element insertion at the transcribed
non-developmental transcription of the yellow locus in Drosophila
mela-translated region of the gene.
nogaster. Genes Dev. 1: 996–1004. A relatively weak effect of the phP1mutation in several
Geyer, P. K.,andV. G. Corces,1992 DNA position-specific repres-sion of transcription by a Drosophila Zn finger protein. Genes
cases may be explained by the low level of the P-Ph
Dev. 6: 1865–1873.
protein produced by the phP1
allele. To examine this
Geyer, P. K., C. SpanaandV. G. Corces,1986 On the molecular
possibility, we are constructing the transgenic lines pro- mechanism of gypsy-induced mutations at the yellow locus of Drosophila melanogaster. EMBO J. 5: 2657–2662.
ducing a large amount of P-Ph protein.
Geyer, P. K., M. M. GreenandV. G. Corces,1990 Tissue-specific The authors are greatly indebted to M. Gause for her help in some transcriptional enhancers may act in trans on the gene located experiments and to K. O’Hare and P. Geyer for plasmids of the singed in the homologous chromosome: the molecular basis of transvec-and yellow genes, respectively. This work was supported by the Russian tion in Drosophila. EMBO J. 9: 2247–2256.
Harrison, D. A., D. A. Gdula, R. S. CoyneandV. G. Corces,1993 Foundation for Basic Research, N 96-15-97032 and N 97-04-48742, by
A leucine zipper domain of the suppressor of Hairy-wing protein an International Research Scholar’s award from the Howard Hughes
mediates its repressive effect on enhancer function. Genes Dev. Medical Institute, and by a Human Frontier Science Program
Organi-7:1966–1978. zation grant to P.G.
Hawley, R. S., R. A. Steuber, G. H. Marcus, R. Sohn, D. M. Baronas et al., 1988 Molecular analysis of an unstable P element insertion at the singed locus of Drosophila melanogaster : evidence for intracis-tronic transposition of a P element. Genetics 119: 85–94. LITERATURE CITED
Jurgens, G.,1985 A group of genes controlling the spatial expres-sion of the bithorax complex in Drosophila. Nature 316: 153–155.
Andrews, J. D.,andG. B. Gloor,1995 A role for the Kp leucine
zipper in regulating P element transposition in Drosophila melano- Kares, R. E.,and G. M. Rubin,1984 Analysis of P transposable element functions in Drosophila. Cell 38: 135–146.