HISTOMORPHOMETRICAL STUDY OF PREGNANCY INDUCED
CHANGES IN THE ISLETS OF LANGERHANS
Alaa K. Al-Asadi*1, May F. Al-Habib(PhD)2, Ula M. Al-Kawaz1(FIBMS)
1
High Institute of Infertility Diagnosis and ART / Al-Nahrain University
2
College of Medicine / Al-Nahrain University.
ABSTRACT
Background: The pancreas is a mixed gland consist of both exocrine
and endocrine tissues the exocrine consist of acini composed of multi
acinar cells; while the endocrine part represented by islet of
Langerhans, these tissues adapted during pregnancy due to nutritional
demands and hormonal changes. Aims of the study: To assess the
histomorphometrical changes induced by pregnancy in islets of
Langerhans including size, number, shape & distribution. Materials
and methods: Forty mice were divided into 2 groups; 20 adult mice in
control group, 20 pregnant mice in the late pregnancy were sacrificed
to extract the pancreas and prepared to blocking sectioning and staining with H&E stain for
microscopic examination and Image J software were used for histomotphometrics. Result:
The pancreatic tissue mass increased in both exocrine and endocrine parts with their
cellularity and vascularity and a significant increasing in the islet size and their numbers with
showing numerous newly formed islets and demarcating coalescence of adjacent islets.
Discussion: The changes in pancreatic tissue during pregnancy is an adaptation changes due
to hormonal and nutritional demands of the fetous. There was an increase in the total
pancreatic mass and the total pancreatic islets due to increase in both β cell hyperplasia and
hypertrophy during pregnancy. Conclusion: The pancreatic tissue changed during pregnancy
like increase in islet number, size, cellularity and their vascularity with increasing in exocrine
part.
KEYWORDS: Pancreas, exocrine, endocrine, islet of Langerhans.
Volume 5, Issue 11, 340-350. Research Article ISSN 2277– 7105
*Corresponding Author
Alaa K. Al-Asadi
High Institute of Infertility
Diagnosis and ART /
Al-Nahrain University. Article Received on 15 Sep. 2016,
Revised on 06 Oct. 2016, Accepted on 27 Oct. 2016
INTRODUCTION
Pancreas is a mixed organ of exocrine and endocrine glands, in mice it is present as
mesenteric type, where the pancreas is relatively diffused, as a scattered masses across the
duodenal mesentery surround by capsule.[1, 2]
The exocrine part consist of serous acini responsible of synthesis, processing and secreting
digestive enzymes or called pancreatic juice that assist digestion and absorption of nutrients
in the small intestine.[3]
The endocrine portion represented by Islets of Langerhans (IL), they are highly vascularized
clusters of different types of cells surrounded by reticular fiber networks; these clusters varies
in number, size and distribution of cells depend on: region, age, disease and physiological
state. The islets are composed of pale staining cords of polygonal cells surrounded by a
network of capillaries. All islets occupy about 2% of the total pancreatic tissue (4, 5). Each
islet has 5 types of cells: 1) Alpha cell (𝛼 cell) which occupy (11-20%) of islet mass cells
which are found peripherally, produce glucagon hormone to control the blood glucose level
especially in hypoglycemia 2) Beta cell (β cell) which occupy (70-83%) of islet
mass-produces amylin and Insulin, which is released by stimulation of increase blood glucose in
hyperglycemia 3) Delta cell (δ cell) which occupy (5-10%) of islet mass produce Vasoactive
Intestinal Peptide (VIP) and somatostatin which in paracrine mode and inhibit HCL secretion
from parietal cells of the stomach 4) Pancreatic poly peptide secreting cells (PP cells)S which
occupy (1%) of islet mass and also called F cell these cells secret polypeptide to inhibit
somatostatin secretion and block secretion of bile 5) Epsilon cell (ε cell) (G cell), which
occupy 1% of islet mass and produce Ghrelin and it is also called Gastrin producing
cells.[6, 7]
These islets may change during the life especially during pregnancy and certain physiological
and pathological conditions. The distribution and composition of the islets cells vary within
the species depends on the physiological state.[8]
Pancreatic β Cell Mass (PβCM) are changed depending on the metabolic demand such as
pregnancy and there are another factors that effect on replication of PβCM like: nutrient
especially the glucose, Insulin like Growth Factor, Growth Hormone, Hepatocyte Growth
Factor, Prolactin, Placental Lactogen and Serotonin, and some diseases like diabetes Miletus
DM causing increase in PβCM.[9]
We aimed in this study to assess the histomorphometrical changes induced by pregnancy in
islets of Langerhans including size, number, shape & distribution.
MATERIALS AND METHODS
40 mature female of white Swiss-webester mice Mus musculus got from the animal house in
the High Institute of Infertility Diagnosis and Assisted Reproductive Technology in
Al-Nahrain University. The virgin mice were chosen in age range between (10-12) weeks old.
The mice in estrus phase which showed cornified cells in the smear were separated in
separate cage each 2 female mice with one male in one cage for mating checked the vaginal
plug next day at early morning to reveal the occurrence of pregnancy, so the first day of
gestation is the day after the vaginal plug was found (10).
Taking specimens: all specimens were collected after mouse was sacrificed by cervical
dislocation after anasthesia to extract the pancreas; then tissue was fixed in 10% formalin for
24 hrs. . The specimen dehydrated by using an ascending concentrations of alcohol 70 %,
90%, 100%, Cleared by Xylene, Infiltrated and Blocked with paraffin wax, Serial sections of
4μ and 5μ thickness were taken, Staining with: Hematoxylin and Eosin.
Morphometric image analysis was done using the software ImageJ software (Java-based
image processing program developed at the National Institutes of Health, USA), version 1.47
p (http://imagej.nih.gov/ij/docs/guide).
RESULT
Histological architecture of the exocrine pancreatic tissue of the pancreas composed of small
lobules, each lobule is formed of closely packed serous acini separated by a thin type of
connective tissue and capillaries, which has distinctive features like: presence of Islets of
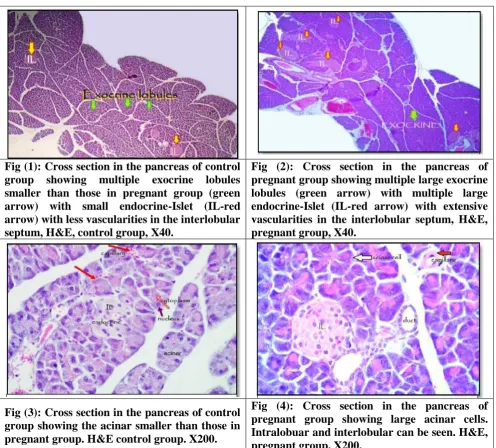
Langerhans. These lobules in pregnant group are bigger than in control group (Fig: 1) and
(Fig: 2)
A closer look to the exocrine acini showed that both acini and lobules are larger in pregnant
group than those in control (non-pregnant) group, it was also notice the extensive blood
vessels infiltration was seen in the interlobular septum (Fig: 2)
The acinar cells in the pregnant group seem to be larger than those in control group; while the
pregnant group as well as control group, the acini are separated by a delicate connective
tissue containing blood vessels, lymphatics, nerves and excretory ducts. (Fig: 3) and (Fig: 4)
The acinar cell has a spherical nucleus, lies on the base, contain a distinct nucleoli or
chromatin in clumps.; the nucleus surrounded by a cytoplasm, which has two parts the basal
part which is basophil, and apical part contain acidophil secretion (zymogenic) granules or
vacuoles (Fig: 5).
The blood vessels seen in different forms in the connective tissue between the acini and
lobules, blood vessels in pregnant group showed increased in numbers and size more than
those in control group within the Islet of Langerhans (Fig: 3) and(Fig: 4).
Fig (1): Cross section in the pancreas of control group showing multiple exocrine lobules smaller than those in pregnant group (green arrow) with small endocrine-Islet (IL-red arrow) with less vascularities in the interlobular septum, H&E, control group, X40.
Fig (2): Cross section in the pancreas of pregnant group showing multiple large exocrine lobules (green arrow) with multiple large endocrine-Islet (IL-red arrow) with extensive vascularities in the interlobular septum, H&E, pregnant group, X40.
Fig (3): Cross section in the pancreas of control group showing the acinar smaller than those in pregnant group. H&E control group. X200.
[image:4.595.51.548.280.728.2]Fig (5): Cross section in the pancreas: where acinar cell has a spherical nucleus, lies on the base (white arrow), contain a distinct nucleoli or chromatin in clumps (black arrow); the nucleus surrounded by a cytoplasm, which has two parts the basal part which is basophil (straighten arrow), and apical part contain acidophil secretion (zymogenic) granules or vacuoles (doted arrow) H&E, pregnant X1000.
Histological examination of the endocrine pancreatic tissue: Coalescence of adjacent
islets due to the enlargement in size of each islet and increasing in their cellularity are seen in
pregnant group exclusively. In additional to see multiple newly formed islets started to appear
in experimental group, they gained an enlargement in their size with the progression of
pregnancy.
These islets composed of a group of cells supported by fine connective tissue fibers pervaded
by capillaries which is a very thin layer of fibers surrounded each islet; the arrangement of
the islets cells were the same as in both groups. (Fig: 3) and (Fig: 4).
The size of the islet cells in pregnant group seems to be larger than the size
of the islet cells in control group with pale cytoplasm and highly basophilic nuclei with
distinct nucleoli, each islet contains secretory cells of different types but not recognized from
one to another by (H&E) stain in both groups; the islet cells arranged in a thin irregular
branched cords of cells or clusters, the nuclei of the cells basically were regular rounded with
coarsely clumped chromatin and inconspicuous nuclei., the cytoplasm of these cells were pale
amphiphilic.
Morphometrical measurements of the endocrine pancreatic tissue: The endocrine
pancreatic tissue (Islets of Langerhans) and the whole pancreatic tissue were measured using
Image-J software and it was found that the whole islets size in control group (2,262.1
μ3±SE56.5) of total pancreatic mass that means it occupy 1.098% of total pancreatic tissue area; While in pregnant group, the islets have the size (5,745.7 μ3 ±SE360.4) of total
pancreatic mass as a percentage of 13.628%.
Whereas the exocrine also increased slightly it was (190,507.8 μ3 ±SE:1980) in control group
[image:5.595.117.540.73.218.2]the whole pancreatic tissue (192,769.8 μ3±SE: 1159,2) in control group while in pregnant group became (200,556 μ3±SE:2438.7).
There was increasing in number of Islets of Langerhans in pregnant group as compared with
control group and the mean for each group was calculated as the mean number of Islets for
pregnant group is (6.6 ±SE 0.327) and for control group is (1.3 ±SE 0.10) .
The islets area of the control group ranged between (51μ2- 210μ2) with the mean of (180.3 μ2± 𝑆𝐸 9.2); While the area of the islets of pancreas observed in pregnant group enlarged more than in those of control group ranged between (32μ2-723μ2) with the mean of
(457.6μ2± 𝑆𝐸 20.4) and the statistical analysis of the Islet area done by using t-test that shows
a significant statistical differences between these groups where the t=5.183 with (P value ≤
0.05).
The area of the islet cells in pregnant group seems to be larger than the area of the islet cells
in control group with the mean (40.1μ2 ± SE 1.8: 25.15 μ2 ±SE 1.5) respectively.
DISCUSSION
During pregnancy the pancreas adapt itself functionally specially the endocrine portion of
islet cells which is one of the aims of the study where the insulin demands increased during
pregnancy due to the enhanced insulin resistance of the maternal tissue and due to increase
food intake.[11, 12]
Some authors showed that during normal pregnancy in the rat, the number of β cell is
increased (hyperplasia) as the volume of the individual β cell (hypertrophy) but there are no
increase in the β cell mass during diabetic pregnancy, the scientist confirm that, there is reduced adaptation of β cell proliferation was accompanied by reduced islet levels of menin
and its targets, this expression prevent islet expansion.[13]
In this study there was an increase in the total pancreatic mass and the total pancreatic islets
during pregnancy with an agreement of different studies which explain the growth of the
islets is due to both β cell hyperplasia and hypertrophy.[14, 15]
Menin controls islets growth in
pregnancy, which stimulated proliferation of maternal pancreatic islet β cells[16]
; Serotonin
also acts to stimulate β cell proliferation.[17]
Kawai & Kishi (1999) confirmed that reduced adaptation of the pancreatic β cells during
pregnancy is the major causal factor for gestational diabetes.[13]
Not only functional changes occurred during pregnancy but also histological changes
happened in both exocrine and endocrine parts. The islet of Langerhans are surrounded
extensively by basophilic stained acini in control (non-pregnant) group, mice changed into
slightly acidophilic acini filled with dark basophilic granules in pregnant group (3).
In this study the exocrine as well as endocrine vascularity increased in pregnant group more
than control group and enlarged in their diameter, this is agreed with the capillaries network
increased with the islet size enlargements.[18]
The same histological architecture of exocrine part in pregnant group but the acini and the
acinar cells seems to be larger than those in control group, also the granules are more and the
vacuoles are larger than those in control group.
Henics & Wheatley (1999) mentioned that cytoplasmic vacuolation is indicative of
nutritional status as evidenced by the presence of the glycogen. This can vary according to
the age, food provided, diet composition, and season sampled.[19]
While other scientist mentioned that diabetes may cause a diffuse vacuolar changes involving
the β cells of the islets. Different agents and conditions can induce the vacuolation while the
degree of vacuolation depends on the cell type, some are easily vacuolated whereas the others
resist.[19]
In this study The endocrine pancreatic tissue (Islets of Langerhans) in control and pregnant
group by light microscope shows a pale rounded to oval area located in between the acini and
a very thin layer of a connective tissue have been seen which is demarcated the edge of the
islets, these islets were scattered through the pancreatic tissue as irregular, rounded to oval
masses of pale staining cells, separated from the acini by the connective tissue Van et al
(1980).[20]
These islets are unevenly distributed through the pancreas and it is possible to view section
In this study, the number and the size of the islets in pregnant group are increased more than
those in control group, and formation of numerous, small islets and hyper atrophy of the other
islets by coalescence of adjacent islets with increasing in their cellularity compared with
small, rounded and oval islets of control group (3).
Genevay reported that islets not only enlarged in their size but also a neogenesis found in the
section which is mechanism that triggers the generation of new cells from precursor cells
which could potentially originate from ductal cells by ductal neogenesis, already
differentiated to pancreatic cell.[21]
Some endocrine buds within the ductal epithelium of adult rat pancreas during pregnancy as a
stage of an endocrine pancreatic “neohistogenesis” occurring in the adult rat pancreas.[21,22,23]
The total pancreatic tissue increased synchronous with the enlargement of both the islets of
Langerhans and the exocrine lobules during the pregnancy period specially the late third
stage, where the whole pancreatic tissue in pregnant group increased more than those in
control group due to enlargement of both exocrine and endocrine parts; the islets of
Langerhans in the pregnant rat were increased in number and enlarged in their size and
possess a more sensitive mechanism for responding to glucose.[24]
The islets number increased in pregnant group with high significant P value < 0.001as same
as their size increased significantly with P < 0.05; these morphological changes reflect the
effect of placental hormones on pancreatic islet of Langerhans[21, 25] where the islets area in
pregnant group ranged from numerous newly formed islets with area (32μ2) to the larges islet
with area (723 μ2) with demarcated coalescence of adjacent islets forming biggest one.[24 , 25]
It was mentioned that the human and the rodent islets might adapt differently to pregnancy.
First, the extent of this adaptation might be less distinct in humans than in rodents. Second,
the mechanism responsible for this adaptation involves increased in replication and decreased
apoptosis in rodents, but possibly islet neogenesis in humans[21]; β cell proliferation and hypertrophy indicated the endocrine pancreas was able to adapt to the metabolic changes of
pregnancy with hyperplasia of the islets tissue and by implication, increased insulin synthesis
and secretion.[24] The cells inside the islets of control group were seen to be arranged in thin
increased significantly with P value<0.05 in pregnant group more than those in control group
as Genova et al (2014).[26]
Peterson et al (1993) reported that β cell mass was determined by the product of the number and size of the β cells.[27]
CONCLUSION
The study conclude that both exocrine and endocrine part of the pancreatic tissue with their
vascularity changed during the pregnancy periods; Increase the total pancreatic tissue in both
exocrine and endocrine parts with their cellularity, vascularity, islets number, islets size and
newly formed islets during pregnancy.
REFERENCE
1. Abraham L.K., Laura L. T. Histology and cell biology 3rd ed. Saundres (2012).
2. Dintizis S. M., Liggitt D. Comparative anatomy and histology. USA Elsever (2012).
3. Abood AH., Al Habib MF, AbdulAmeer HH. Histochemical histomorphometrical &
ultrastructural studies of B cells & exocrine gland of mice pancreas in pregnancy &
postpartum periods. MSC thesis. Al-Nahrain University (2013).
4. Aerts L., Van Assche F.A., Deprins F. “A morphological study of the endocrine pancreas
in human pregnancy” Brit. J. of Obest. & Gyn. 1978; 85: 818-820.
5. Cherly L.S., Mary L. A practical guide to the histology of the mouse 1st ed.. MRC
Harwell U. K. John Wiley & Sons ltd. 2014.
6. Mostafa M. N, Ahmed A.E., Salleh M.A., Mohammad T. “Isolated Pancreatic Islets of
the rat: an immunohistochemical and morphometric study”. The Anat. reco. 1993; 237:
489-497
7. Victor P. Eroschenko. Atlas of histology with functional correlation 12th ed. Wolters
Kluwer. Lippincott. USA. (2013).
8. Aerts L., Van Assche F.A. “Ultrastructural changes of the endocrine pancreas in pregnant
rat”. Diabeto. 1975; 11: 285-289.
9. Tanaka Y., Toyota N., Fujimoto K., Maekawa T., AsakuraT., IchioT., Yoshimura K.,
Yamamoto T., Kozuka Y., Murata K. “Immunohistochemical studies of fetal and
maternal endocrine pancreases during pregnancy and the puerperium
instreptozotocin-induced diabetic rats”.Nihon Sanka Fujika Gakkai Zasshi// Japan participation Fujica
10.Suckow M. A., Danneman P., Brayton C. The Laboratory MOUSE. CRC Press LLC.
2001.
11.Adeyemi DO., Komolafe OA., Adewole OS. et al. Histomorphology and morphometric
studies of the pancreatic islet cells of diabetic rat treated with extracta of Annona
muricata. Folia.Motphol. 2010; 69(2): 92-100.
12.Kawai M., Kishi K. Adaptation of pancreatic islet B cells during the last third of
pregnancy: regulation of B cell function and proliferation by lactogenic hormones in rats,
europ.J.Endocr. 1999; 141: 419-425.
13.Devlieger R., Cas teel K.,VanAssche FA. Reduced adaptation of the pancreatic B cell
during pregnancy is the major causal factor for gestational diabetes. Acta. Obstetr.Gyne.
2008; 87: 1266-1270.
14.Sorenson RL., Berlje TC. “Adaptation of islets of Langerhans to pregnancy: beta- cell
growth, enhanced insulin secretion and the role of lactogenic hormones”. Horm. Metab.
Res. Jun, 1997; 29(6): 301-7.
15.Dean P.M.”Ultrastructural morphometry of the pancreatic β cell”. Diabet. 1973; 9:
115-119.
16.Karnik SK, Chen H et al.”Menin controls growth of pancreatic beta cells in pregnant mice
and promotes gestational diabetes mellitus”. Science. Nov 2007; 2; 318(5851): 806-9.
17.Kim H., Toyofuku Y., et al. ”Serotonin regulates pancreatic beta cell mass during
pregnancy”. Nat. Med.USA. July 2010; 16(7): 804-808.
18.Jansson L.the regulation of pancreatic islet blood flow. Diab. Metab. Rev. 1994; 10(4):
407-416.
19.Henics T.,Wheatley DN. Cytoplasmic vacuolation, adaptation, and cell death: Aviewon
new perspectives and features. Biology of the cell 1999; 91: 485-498.
20.Van Assche FA., Gepts W., Aert L.Immunocytochemical study of the endocrine pancreas
in the rat during normal pregnancy and during experimental diabetic pregnancy.
Diabetalogia, 1980; 18: 487-491.
21.Genevay M.,Pontes H., Meda P. B cell adaptation in pregnancy: a major difference
between human and rodent. Diabetalogia, 2010; 53: 2089-2092.
22.Green IC., TaylorKW. Effect of pregnancy in the rat on the size and insulin secretory
response of the islets of Langerhans. J. Endocr. 1972; 54: 317-325.
23.Bertelli E., Regoli M., Bastianini A. Endocrine tissue associated with the pancreatic
ductal system: A light and electron microscopic study of the adult rat pancreas with
24.Aerts L., Van Assche F.A., Gepts W. “Morphological changes in the endocrine pancreas
in pregnant rats in experimental diabetes”. J. Endoc. 1979; 80: 175-179.
25.Schraenen A., Lemaire K., Faudeur G., Hendrickx N., Gramvik M., Van Lommel L.,
Mallet J., Vodgdani G., Gilon P., Binart N., Veld P., and Schuit F. “ Placental lactogens
induce serotonin biosynthesis in a subset of mouse beta cells during pregnancy”.
Diabetologia, Dec. 2010; 53(12): 2589-2599.
26.Genova M.P., Todorova K., Atanasova B., and Tzatchev. “Assessment of beta cell
function during pregnancy and after delivery”. Acta. Medica. Bulgarica.vol.XLI.2014
27.Peterson JS, Russel S, et al “Differential expression of GAD in rat and human islets”.