Engineering and Technology Journal ISSN: 2456-3358 Impact Factor: 4.449
Volume 2 Issue 9 Sept. 2017
247
Codon Usage in Hemoglobin Coding Genes
Li Gun1, Tian Han2, Ren Yumiao3, Du Ning4
1,2,3,4
Department of Biomedical Engineering, School of Electronics and Information Engineering,
Xi'an Technological University, Xi'an, Shaanxi Province, China
ARTICLE INFO ABSTRACT
corresponding Author:
Li Gun1
Dept. of Biomedical Eng., School of Electronics and
Information Eng., Xi'an
Technological Univ., Xi'an,
Shaanxi Province, China
Hemoglobin is a protein responsible for the transport of oxygen. The preference of hemoglobin coding gene is analyzed by CodonW. The relationship between the A3 / (A3 + T3) and G3 / (G3 + C3) and the relationship between GC12 and
GC3 are all investigated. Finally, the preference of hemoglobin coding gene is
shown and discussed. The results show that codon use preference of CUG, CGU, UAA, ACC, GUG and GGC are greater than 2.5. This information facilitates an improved understanding of the structural, functional of hemoglobin
KEYWORDS: hemoglobin, codon usage, codon preference..
Introduction
Proteins are machines that perform a variety of
important function in the body. Some proteins are
composed of multiple peptide chains, each of which
is called a subunit, and the specific spatial
relationship between subunits is called the quaternary structure of the protein. Hemoglobin is a
protein that makes the blood red and consists of
four strands, two alpha chains and two beta chains.
As the main carrier of genetic information, the
number and order of the four bases of A, T, C and G
in the DNA molecule contain very important
information. Analyzing the DNA sequence
characteristics is benefit to understanding the
biological function of the sequence and to finding
the characteristic information related to the specific
function in the sequence. When the codon usage
preference is concerned, many scientists studied vary gens recently, such as virus [1], Y-linked genes
[2], CYB gene [3] and SRY gene [4], et al. In the
virus gene area, most influential virus gene are all
studied by scientist timely, such as Zika [1],
classical swine fever virus [5], Japanese
encephalitis virus [6], enterovirus D68 strains [7], and so on [8-10]. In fact, codon usage bias has
would reveal many mechanism in biology [11-13]
and affect by many factors [14-17]. In this paper,
the codon usage in hemoglobin coding genes is
studied via the CodonW, and the PR2-bias plot
analysis: A3 / (A3 + T3) vs. G3 / (G3 + C3) and the
neutral evolution analysis: GC12 vs. GC3 are plotted
and discussed.
Materials and Methods
This paper considers the most common type of
the accession numbers are: NP_000549.1 and NP_000509.1.
CodonW [18] is used to calculate the CAI,
CBI, ENC and so on, the calculated results are
further processed for getting the relative
synonymous codon usage (RSCU), the PR2-bias
plot analysis: A3 / (A3 + T3) vs. G3 / (G3 + C3),
the neutral evolution analysis: GC12 vs. GC3, et al.
Results and discussion
Codon system is used to predict the codon usage
characteristics of hemoglobin, and the overall
results are shown in Table 1. in the table 1 Codon Adaptation Index (CAI) ranges from 0 to 1, the
greater the value, the stronger the sub-preference of
the codon and the higher the gene expression level, the CAI value of hemoglobin are 0.4 in HBα1 and 0.36 in HBβ respectively. The effective codon number (ENC) reflects the degree of preference for
the use of the codon, with the range of 20-61, the
smaller the value, indicating that the stronger codon
usage preference, the ENC value of hemoglobin are 29.84 in HBα1 and 34.01 in HBβ respectively. Frequency of Optimal Codons (FOP) refers to the
percentage of the optimal codon used, the FOP value of hemoglobin are 0.64 in HBα1 and 0.49 in HBβ respectively. Codon Bias Index (CBI) is the component that reflects the expression of a superior codon in a particular gene, and this index has a
good correlation with the ENC value generally.
Table 1. Nucleotide composition in the coding sequences of hemoglobin.
Title Accession T3s C3s A3s G3s CAI CBI Fop Nc GC3s GC
HBα1 NP_000549.1 0.11 0.61 0.02 0.47 0.4 0.38 0.64 29.84 0.89 0.65 HBβ NP_000509.1 0.35 0.36 0.07 0.45 0.36 0.13 0.49 34.01 0.65 0.56
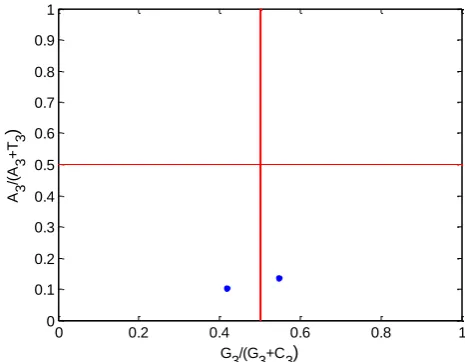
The parity rule states that if there is no mutation in
the two complementary strands of DNA or the bias
on the selection effect, the base content should have
A = T and G = C. This method was used to analyze
the PR2 bias occurring at the third place of the
codon. The center of the graph follows the PR2
principle, A = T and G = C, that is the vertical and
horizontal coordinates are 0.5. The vector from this
center represents the degree and direction of the
PR2 bias. The parity preference is mainly used to
analyze the relationship between pyrimidine and purine at 3rd base of a codon. The content of
pyrimidine (C + T) at the third position of the
hemoglobin codon gene is greater than the purine
content (A + G) (Figure 1).
0 0.2 0.4 0.6 0.8 1
0 0.1 0.2 0.3 0.4 0.5 0.6 0.7 0.8 0.9 1
G 3/(G3+C3)
A3
/(
A3
+T
3
)
Figure 1. PR2-bias plot analysis
In neutrality plotting, GC12 is as the ordinate, and
GC3 is as the abscissa, each point in the figure
Volume 2 Issue 9 Sept. 2017
249
Figure 2, the result shows that the distribution of GC12 is 56.73% and 55.83% in HBα1 and HBβ
respectively, GC3 is 81.41% and 59.51% in HBα1
and HBβ respectively, GC 12 distribution was significantly smaller. These values are distributed
diagonally in the vicinity, and for these regions,
base mutations may be a major factor influencing
codon preference (Figure 2).
0 0.2 0.4 0.6 0.8 1
0 0.1 0.2 0.3 0.4 0.5 0.6 0.7 0.8 0.9 1 GC3 GC 1 2
y = 0.04121x + 0.53376
Figure 2. Neutrality plot of GC12 vs. GC3
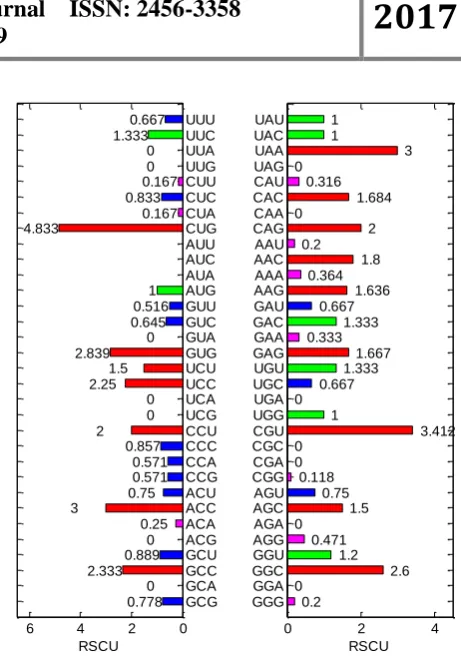
The relative synonymous codon usage is an
estimate of the preference for the usage of synonymous codons, defined as the ratio of the
observed value of the number of synonymous
codons used to the expected number of occurrences
of the codon. If the codon usage is not preferred,
the RSCU value is 1; if a codon is used frequently,
the RSCU value is greater than 1; if a codon usage
frequency is low, the RSCU value is less than 1.
The RSCU value intuitively reflects the preference
for codon usage in hemoglobin (Figure 3). The results show that codon usage preference of CUG,
CGU, UAA, ACC, GUG and GGC are greater than
2.5. Because it is simple to calculate and intuitively
reflects the preference of codon usage, it is usually
used as a measurement of the codon usage
preference analysis. 0 2 4 6 UUU UUC UUA UUG CUU CUC CUA CUG AUU AUC AUA AUG GUU GUC GUA GUG UCU UCC UCA UCG CCU CCC CCA CCG ACU ACC ACA ACG GCU GCC GCA GCG 0.667 1.333 0 0 0.167 0.833 0.167 4.833 1 0.516 0.645 0 2.839 1.5 2.25 0 0 2 0.857 0.571 0.571 0.75 3 0.25 0 0.889 2.333 0 0.778 RSCU
0 2 4
UAU UAC UAA UAG CAU CAC CAA CAG AAU AAC AAA AAG GAU GAC GAA GAG UGU UGC UGA UGG CGU CGC CGA CGG AGU AGC AGA AGG GGU GGC GGA GGG 1 1 3 0 0.316 1.684 0 2 0.2 1.8 0.364 1.636 0.667 1.333 0.333 1.667 1.333 0.667 0 1 3.412 0 0 0.118 0.75 1.5 0 0.471 1.2 2.6 0 0.2 RSCU
Figure 3. RSCU of the hemoglobin coding gene
Codons as a link between nucleic acids and proteins,
it can be used to characterize the evolutionary
relationships within the genome or between
genomes [19, 20]. For example, the same genes of
different kinds of creatures or the different genes of
the same creature have different characteristics of codon usage [21]. The closer the genetic
relationship is, the closer the codon preference is, the closer the genetic relationship is, and the less
difference in codon preference [22]. One kind of a
gene of different species or the same species may
have different characteristics in codon usage [23].
However, the frequencies of the different codons
that encode the amino acids for the same species
may different [24]. Different codon usage could
results differences frequency in amino acids encode
even in the same amino acid, resulting in quite different differences between different species
Conclusion
Codons are the basic units of life information
transmission. In a certain species or genotype the
phenomenon of unequal use of synonymous codons
is common, where the synonymous codons that are
preferentially used are called optimal codons. There
are significant differences in codon usage
preferences in different organisms, meanwhile,
codon usage preference not only plays an important
regulatory role map at the level of gene expression,
but also helps to improve the accuracy and
efficiency of translation. In this paper, the
preference of hemoglobin coding gene is analyzed, such as the relationship between the A3 / (A3 + T3)
and G3 / (G3 + C3), the relationship between GC12
and GC3 are all studied and the preference of
hemoglobin coding gene is also discussed. The
results show that codon usage preference of CUG is
4.833, which is the largest, codon usage preference
CGU is 3.412, codon usage preference UAA is 3,
and so on. All these information are important for
studying the structural, functional of hemoglobin.
References
1. Snawar Hussain, Sahibzada Tasleem Rasool.
Analysis of synonymous codon usage in
Zika virus. Acta Tropica, 2017, 173:
136-146
2. Monisha Nath Choudhury, Arif Uddin,
Supriyo Chakraborty. Codon usage bias and
its influencing factors for Y-linked genes in
human. Computational Biology and
Chemistry, 2017, 69: 77-86
3. Arif Uddin, Supriyo Chakraborty. Codon usage trend in mitochondrial CYB gene.
Gene, 2016, 586(1): 105-114
4. Supriyo Chakraborty, Arif Uddin, Monisha
codon usage bias of SRY gene across mammals Gene, 2017, 630: 13-20
5. Xin Xu, Dongliang Fei, Huansheng Han,
Honggui Liu, Hua Zhang. Comparative
characterization analysis of synonymous
codon usage bias in classical swine fever
virus. Microbial Pathogenesis, 2017, 107:
368-371
6. Niraj K. Singh, Anuj Tyagi, Rajinder Kaur,
Ramneek Verma, Praveen K. Gupta.
Characterization of codon usage pattern and
influencing factors in Japanese encephalitis
virus. Virus Research, 2016, 221: 58-65 7. Uladzimir U. Karniychuk. Analysis of the
synonymous codon usage bias in recently
emerged enterovirus D68 strains. Virus
Research, 2016, 223: 73-79
8. Yu Xu, Renyong Jia, Zhilong Zhang,
Yanyan Lu, Anchun Cheng. Analysis of
synonymous codon usage pattern in duck
circovirus. Gene, 2015, 557(2): 138-145
9. Sudhir Morla, Aditi Makhija, Sachin Kumar.
Synonymous codon usage pattern in glycoprotein gene of rabies virus. Gene,
2016, 584(1): 1-6
10.Gonzalo Moratorio, Andrés Iriarte, Pilar
Moreno, Héctor Musto, Juan Cristina. A
detailed comparative analysis on the overall
codon usage patterns in West Nile virus.
Infection, Genetics and Evolution, 2013, 14:
396-400
11.Arup Kumar Malakar, Binata Halder,
Prosenjit Paul, Supriyo Chakraborty.
Cytochrome P450 genes in coronary artery diseases: Codon usage analysis reveals
genomic GC adaptation. Gene, 2016, 590(1):
Volume 2 Issue 9 Sept. 2017
251
Chen, Xiangyang Xue. Codon usage bias in human cytomegalovirus and its biological
implication. Gene, 2014, 545(1): 5-14
13.Abhishek Subramanian, Ram Rup Sarkar.
Comparison of codon usage bias across
Leishmania and Trypanosomatids to
understand mRNA secondary structure,
relative protein abundance and pathway
functions. Genomics, 2015, 106(4): 232-241
14.Max L. Nibert. Mitovirus UGA(Trp) codon
usage parallels that of host mitochondria.
Virology, 2017, 507: 96-100
15.Devi Lal, Mansi Verma, Susanta K. Behura, Rup Lal. Codon usage bias in phylum
Actinobacteria: relevance to environmental
adaptation and host pathogenicity. Research
in Microbiology, 2016, 167(8): 669-677
16.Mei He, Chun-Bo Teng. Divergence and
codon usage bias of Betanodavirus, a
neurotropic pathogen in fish. Molecular
Phylogenetics and Evolution, 2015, 83:
137-142
17.Sheng-Lin Shi, Yi-Ren Jiang, Rui-Sheng Yang, Yong Wang, Li Qin. Codon usage in
Alphabaculovirus and Betabaculovirus
hosted by the same insect species is weak,
selection dominated and exhibits no more
similar patterns than expected. Infection,
Genetics and Evolution, 2016, 44: 412-417
18.Peden, J.F., 1999. CodonW. (PhD Thesis).
University of Nottingham, Nottinghamshire.
19.Yang Liang, Mei He, Chun-Bo Teng.
Evolution of the vesicular stomatitis viruses:
Divergence and codon usage bias. Virus
Research, 2014, 192: 46-51
20.Arghya Kamal Bishal, Rashmi Mukherjee,
Chandan Chakraborty. Synonymous codon
usage pattern analysis of Hepatitis D virus.
Virus Research, 2013, 173(2): 350-353
21.Laura Kubatko, Premal Shah, Radu Herbei,
Michael A. Gilchrist. A codon model of
nucleotide substitution with selection on
synonymous codon usage. Molecular
Phylogenetics and Evolution, 2016, 94:
290-297
22.Jasmine Saini, Uri Hershberg. B cell
Variable genes have evolved their codon usage to focus the targeted patterns of
somatic mutation on the complementarity
determining regions. Molecular
Immunology, 2015, 65(1): 157-167
23.Tonghai Yu, Jinsong Li, Yang Yang, Liu Qi.
Codon usage patterns and adaptive
evolution of marine unicellular
cyanobacteria Synechococcus and
Prochlorococcus. Molecular Phylogenetics
and Evolution, 2012, 62(1): 206-213
24.Ratna Prabha, Dhananjaya P. Singh, Swati
Sinha, Khurshid Ahmad, Anil Rai.
Genome-wide comparative analysis of
codon usage bias and codon context patterns
among cyanobacterial genomes. Marine