Copyright 0 1988 by the Genetics Society of America
Effects of
DNA
Heterologies on Bacteriophage
X
Packaging
Rebecca
Kurth
Pearson’ and Maurice S. FoxDepartment of Biology, Massachusetts Institute of Technology, Cambridge, Massachusetts 02139
Manuscript received April 3, 1987 Revised copy accepted September 18, 1987
ABSTRACT
We have examined the impact of D N A heterologies on the packaging of X DNA in vitro. Heterology- containing D N A molecules were constructed by denaturing and reannealing a mixture of D N A from
cI+ phage and D N A from phage carrying small insertion or deletion mutations in the cI gene. We found that molecules with heterologies of up to 19 base pairs (bp) can be packaged as viable heterozygous phage with approximately the same efficiency as molecules with a base pair mismatch. In contrast, with a heterology of 26-bp heterozygous plaque formers are rare. I n principle, the absence of cI heterozygotes among packaged phage may be due either to a failure to encapsidate the DNA or a failure to inject the packaged DNA on infection. Southern blot analysis of DNA isolated from packaged phage indicates that DNA harboring a 26-bp heterology is almost completely absent in packaged phage. Thus, an upper limit has been established for the size of heterology that can be accommodated by the packaging apparatus. The size of the connector portal could be the basis for this limit.
P
RODUCTS of recombination harbor regions ofheteroduplex
D N A
(for review, see FOX 1978;RADDING 1982). If recombining parental molecules
are distinguished by the presence of an insertion or deletion allele in one of them, the inclusion of such an allele within the heteroduplex region would result in the presence of a heterology. In the case of bacte- riophage, the presence of such structures could com- promise the encapsidation process and thereby influ- ence the likelihood of recovering them as viable re- combination products. Several investigations of phage
T4 morphogenesis have suggested that the encapsi-
dation or “packaging” of
D N A
by bacteriophages is in fact sensitive to structural alterations in theD N A
duplex (for review, see BLACK and SHOWE 1983).
T h e bacteriophages which contain doub1e;stranded
D N A
are thought to packageD N A
by the same fun- damental mechanism, and many aspects of the processhave been characterized both genetically and bio-
chemically for several phages (for review, see BAZINET
and KING 1985; EARNSHAW and CASJENS 1980).
Three basic components interact during packaging: concatemeric
D N A ,
a processed precapsid or “pro- head,” and certain auxiliary packaging proteins. T h eD N A
and proheads are the products of two distinct synthetic pathways of the phage. T h eD N A
replicationpathway produces concatemers of phage chromo-
somes, while coat proteins are assembled into pro-
heads in the head morphogenetic pathway. These two
pathways converge in the packaging reaction, when concatemeric
D N A
is linked to the unique “portal”’
Present address: Department of Cell Biology, Biocenter of the Univer- sity, Klingelbergstrasse 70, Basel, Switzerland.Genetics 1 1 8 5-12 (January, 1988)
vertex of proheads through the action of special pack-
aging proteins which also mediate the subsequent
encapsidation process. Neither the energetic basis nor
the physical mechanism underlying
D N A
transloca-tion and condensation into the prohead is understood for any bacteriophage.
Many mutations have been characterized which re-
sult in prevention of
D N A
packaging, and most of these are characterized by an alteration of the struc- ture of the prohead or auxiliary packaging proteins.Although certain
D N A
structural alterations areknown to prevent packaging in phage T 4 (KEMPER
and JANZ 1976; HSIAO and BLACK 1978; ZACHARY
and BLACK 1981, 1984), the structural requirements
of the
D N A
substrate for packaging remain poorly defined. In particular, it is not known what effectD N A
heterologies have on the process ofD N A
encap-sidation. Since encapsidation seems to require
D N A
passage through the portal vertex, investigation of the effect of the presence of
D N A
heterologies on thisprocess could reveal some features of the tolerance of
the packaging mechanism to perturbations in the du- plex
D N A
structure.T h e effect of
D N A
heterologies on h packaging in vitro has been examined. The heterologies were cre- ated by denaturing and reannealingD N A
from c l +phage with
D N A
from phage carrying small insertion or deletion mutations in the c l gene. We show that heterologies of up to 19 bp can be packaged in vitrowith approximately the same efficiency as a base pair
mismatch; phage heterozygous for c l are found among
the viable products of phage packaging.
A
heterologyTABLE 1
Bacterial strains
Designation Relevant genotype
FA77 dnaBts, su-
M182 lacZ galU galK rpsL
M182mutL mutL218::TnlO lac2 galU
galK rpsL
C600 thr leu lacy sull
C6OOmutL mutL218::TnlO thr leu lacy
S U I 1
BNN45 met gal lacy r-m+ sull suII1
Source
MCMILIN and Russo ( 1 970)
E. SIGNER
S. RAPOSA
APPLEYARD HUISMAN and FOX
(1 986)
D. BOTSTEIN
TABLE 2
Phage strains
Genotype Source
P a m 8 0
Ac160 Oam29
A c I L W ? Pam80 Sam7 plac5ZamNG200 AOam29
Ad286 Oam29 Ad287 Oam29 Ad288 Oam29 Ad289 Oam29 Ad290 Oam29 Ad291 Oam29 kc1292 Oam29 AclAH? Oam29
E. SIGNER
E. SIGNER
LICHTEN and Fox (1 983)
M. LICHTEN
PEARSON and Fox (1 988)
PEARSON and Fox (1 988)
PEARSON and Fox (1988)
PEARSON and Fox (1 988)
PEARSON and Fox ( 1 988) PEARSON and Fox ( 1 988) PEARSON and Fox (1 988)
PEARSON and Fox (1 988)
a 10-fold or more reduction in t h e yield of c l hetero- zygotes. Their frequency is near the limit of t h e detection methods used. In principle, this absence of
c l heterozygotes among the packaged phage may be
due either to a failure to encapsidate the DNA or a failure to inject t h e DNA from the phage packaged with this DNA into a bacterial host. Restriction anal- ysis of DNA isolated from packaged phage indicates that DNA harboring the 26-bp heterology is excluded from packaging. These results suggest that it is some aspect of t h e X packaging process, rather than the injection process, that is sensitive to the size of DNA heterologies.
MATERIALS AND METHODS
Bacterial strains and phage strains: Bacterial and phage
strains are described in Tables 1 and 2, respectively.
Media and buffers: Media for growing cultures, diluting
and plating phage have been described by PEARSON and Fox (1 988).
Preparation of artificial heteroduplex and sham DNA:
High titer phage stocks were grown, concentrated, and purified on density step gradients as described by LICHTEN and Fox (1983). An ODnso measurement of the dialyzed phage preparation was used to estimate its DNA concentra- tion. High titer phage were diluted into SM buffer contain- ing 20 mM EDTA and mixed at a concentration of 50 pg DNA/ml of each parent (1 00 pg/ml total). After "popping" the phage by heating at 65" for 15 min, the phage DNA
mixture was divided into two aliquots. One half was used to prepare heteroduplex (het) DNA: the sample was denatured by adding 1/10 volume 1 M NaOH. After 20 min at room temperature, the mixture was neutralized with 1/10 volume of 2 M Tris-HCI (pH 7.2). The second aliquot of phage DNA was used to make "sham" control DNA: The NaOH and Tris-HCI solutions described above were added simul- taneously to the DNA and allowed to stand at room tem- perature for 20 min. An equal volume of formamide was added to both het and sham samples and the mixtures were incubated for 1 hr at 37". Small samples of the het and sham DNAs were then diluted 1/100 into 20 mM Tris-HCI (pH 8.0) to be used for transfections. The remaining het and sham DNA mixtures were dialyzed for 48 hr at 4" against 1 liter of STE buffer (10 mM Tris-HCI, pH 8.0, 1
mM EDTA, 100 mM NaCI) with three changes, then for 12 hr against 1 liter of 2.5 mM Tris-HCI, 0.1 mM EDTA (pH 8.0). The dialyzed het and sham DNA samples were concen- trated IO-fold by evaporation in a Speed-vac. To promote concatemerization, the DNA samples made up to 10 mM
MgClp were incubated at 42" for 2 hr.
Transfection: One hundred microliters of het or sham
DNA at approximately 0.5 pg/ml was heated 10 min at 65 O
to monomerize the DNA and then quickly chilled on ice. Each sample was mixed with 200 pl of ice-cold calcium- treated competent CGOOmutL cells (MANDEL and HIGA 1970) and successively incubated 15 min on ice and 30 min at 37". Infective centers were plated on trypticase plates, along with 0.2 ml of a fresh plating culture of BNN45 and 2 ml of X top agar. Plates were incubated overnight at 37". Twenty microliters of the transfection mixture generally gave 10 to 50 plaques per plate.
Zn vitro packaging: Five microliters of het or sham D N A
were mixed with a thawed Packagene extract (Promega Biotec) and incubated 2 hr at 22". The Packagene extract is prepared with Xcos2 lysogens and therefore will not pack- age endogenous DNA (ROSENBERG et al. 1985). The pack- aging reaction was stopped by addition of 0.5 ml SM+ buffer (5.85 g/liter NaCI, 20 mM Tris-HCI, pH 7.5, 10 mM MgS04) and 25 pl of CHC13. Packaging efficiencies ranged from 1
X 10' to 3 X lo8 phage per microgram of DNA. Het D N A generally gave about a 5-fold lower yield of phage than its sham counterpart. Packaged phage to be analyzed for cI
allele composition were plated as follows: 0.02 to 0.15 ml of an appropriate dilution of phage was mixed with 0.2 ml of a fresh CGOOmutL plating culture and allowed to adsorb for
15 min at 37". Each sample was then mixed with 2 ml of X
top agar containing 0.2 ml of a fresh plating culture of BNN45 and poured on a trypticase agar plate.
Analysis of plaques for cZ allele composition: Plaques
from transfections and packaged phage were analyzed for their cI allele composition by picking random plaques and streaking them on individual agar plates as described by PEARSON and FOX (1 988).
Isolation of DNA from packaging reactions and from
packaged phage DNA: Five microliters of each het and
Packaging D N A Heterologies in X 7
aged D N A , 0.4 ml of each sham packaging reaction was mixed with 5 PI of the analogous het DNA and treated with
DNAse as above. The packaged phage were purified o n a step gradient after adding 10"' carrier phage (XclAH3 Sum7
plac5ZamNG200) to each DNAse-treated phage sample.
( T h e carrier phage carry a deletion of the fragment used as
probe in Southern analysis.) Each sample received 2.1 ml of SM+ buffer saturated with CsCl and was layered on the bottom of two 7.0 ml steps of 1.6 g/ml and 1.4 g/ml. The layered tube was centrifuged for 2 hr at 24,000 rpm in a
Beckman SW28.1 rotor and the phage band between the 1.6 g/ml and 1.4 g/ml density steps was withdrawn with a
25 gauge needle. CsCl was removed by dialysis overnight at
4" against SM+ buffer. D N A was extracted from the di- alyzed phage by the method of MANIATIS, FRITSCH and SAMBROOK (1 982).
Southern blot procedure: D N A was digested with restric-
tion enzymes (New England Biolabs) according to the sup- plier's instructions, using 10- to 20-fold higher enzyme concentrations. The digests fractionated by electrophoresis i n a 1 % agarose gel, using Tris-borate buffer, and trans- ferred to a Biodyne A nylon membrane (Pall), as described i n MANIATIS, FRITSCH and SAMBROOK (1982). The D N A was immobilized on the Biodyne A membrane by UV- irradiation according to CHURCH and GILBERT (1 984). Hy- bridization conditions and radioactive labeling of the 560
bp cZ/rexA fragment (LICHTEN and FOX 1983) were carried
o u t as outlined by MEINKOTH and WAHL (1 984).
RESULTS
In vitro packaging of heterology containing DNA
molecules: Observations reported previously (PEAR-
SON a n d FOX 1988) have shown that DNA harboring heterologies of up to 19 bp can be matured into viable phage progeny and have suggested that chromosomes harboring heterologies larger than 26 bp cannot. In
order to directly investigate the effect that such struc- tural perturbations have on the maturation of phage
DNA, lambda DNAs containing small heterologies in
c l were prepared and packaged in vitro, and the fre- quency of heterozygotes among the viable products was determined.
Het and sham DNAs were prepared using D N A
from X c l + Pam80 in combination with DNA from each of several different c l - Oam29 phage harboring inser- tion or deletion mutations of different sizes. Since sham DNAs have not been denatured, they simply contain a mixture of the two parental c l + Pam80 a n d
c l - Oam29 phage DNAs. Het D N A mixtures, on the other hand, have been denatured and reannealed, so
they contain cl' Pam80/cl- Oam29 heteroduplex
DNA molecules in addition to the reannealed parental phage DNAs. T h e various c l insertion and deletion nlutations would be expected to create heterologies of different sizes in the heteroduplex D N A molecules.
T h e relative amounts of heteroduplex and parental phage DNAs present in the various het and sham
DNA preparations were determined experimentally
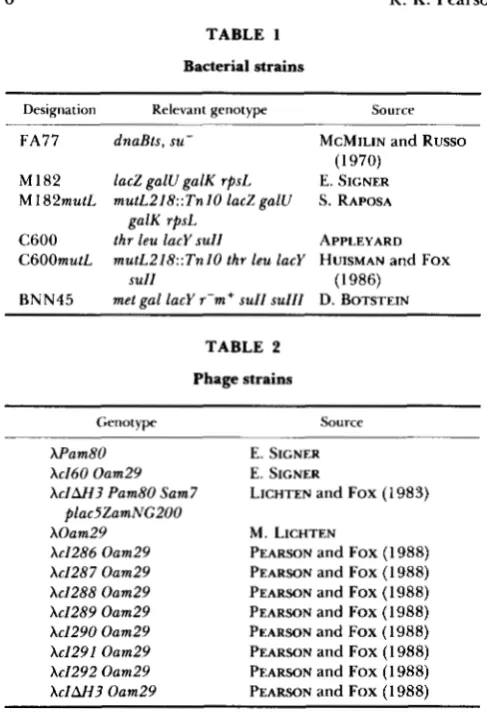
by transfection of an amber suppressing (sull) bacte- rial strain deficient in mismatch repair (C6OOmutL). T h e results are displayed in Table 3. As expected,
TABLE 3
Results of transfection with het and sham DNA
Frequency among transfectants
(X 100)
Observed Predicted
allele change (bp) Turbid Clear het" hetb size
c160 het Base substi- 25 33 42 94
sham tution 37 61 1.9 47 51
c1287 het +4 21 39 40 198
sham 34 62 4.1 45 98
cI289 het +14 20 28 52 99
sham 42 56 2.0 49 50
cI290 het +19 16 30 54 108
sham 48 48 4.0 50 50
~ 1 2 9 1 het +26 27 21 52 100
sham 39 55 6.1 48 49
cI286 het -26 15 49 36 202
sham 39 59 2.1 48 97
cIAH3 het -690 29 45 26 99
sham 45 55 0.0 50 47
GI Sequence Sample
Competent C6OOmutL (sull) cells were transfected with het or sham DNA as described in MATERIALS A N D METHODS. Transfections of c1287 and cI286 het and sham DNAs were carried out on one day and the other transfections carried out on another.
a het: heterozygous. Transfections with sham DNA yield a few
heterozygous plaques presumably as a result of double transfectants. Predicted heterozygous products of transfection were calcu- lated using the quadratic formula, assuming random assortment of reannealed strands. Predicted hets = 2AB, for A = a / a
+
b and B = b/a+
b where a is the frequency of turbid transfectants and b is the frequency of clear transfectants for sham DNA as listed in the table.transfections with sham DNA produce almost exclu- sively turbid and clear plaques. T h e few mixed plaques that are observed are presumably the consequence of
double transfection. T h e ratio of turbid and clear plaques observed in transfection with sham D N A can
be used to calculate the frequency of heterozygous plaques to be expected for the corresponding het
DNA (assuming random assortment of strands). T h e predicted and observed frequencies of heterozygous transfectants are in reasonable agreement for almost all of the het DNA preparations. One exception ap- pears to be the het DNA made with the 690-bp deletion phage, which gives a yield of heterozygous transfectants that is modestly lower than expected; similar reductions were also observed in other prepa- rations of 690-bp het DNA (data not shown).
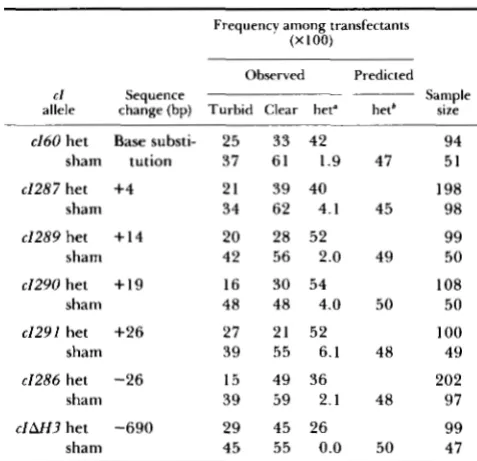
Aliquots from each het and sham DNA preparation were added to X packaging extracts, and the packaged phage were plated on the same suppressing, repair-
deficient strain (CGOOmutL) used in the transfection experiments. T h e results are shown in Table 4. T h e frequency of cl heterozygotes among packaged phage is in all cases lower than expected from the transfec- tion results. A 3-fold reduction is observed with het
TABLE 4
Results of packaging het DNA
Frequency among pack- aged phage (X 100) allele
r l Sequence change Sample
(bp) Turbid Clear het size
e160 Base substitution 45 42 13 388
e1287 +4 23 68 9.5 158
e1289 + I 4 50 44 5.8 393
e1290 + I 9 48 44 8.2 357
e1291 +26 59 40 0.8 357
e1286 -26 21 79 0.0 197
c I A W -690 51 49 0.0 362
substitution mutation in c l , suggesting that the pres-
ence of base pair mismatches in heteroduplex DNA
may result in reduction of packaging efficiency. I t
should be noted that in every case the heteroduplex
DNA harbors mismatches at 0 and P , in addition to a mismatch or heterology in c l . Het DNAs prepared with c l - Oam29 phage harboring 4-bp, 14-bp or 19- bp insertion mutations yield similar frequencies of packaged c l heterozygotes. The presence of a heter- ology of 26 bp results in a 10-fold reduction in the yield of c l heterozygotes, while the yield of c l hetero- zygotes in phage packaged with a 690-bp deletion heterology falls to the limit of this detection method.
Frequencies of heterozygous phage much below 1 0-2
are not readily detected by screening for c l heterozy- gotes.
Another approach to the detection of heterozygous packaged phage, which allows us to examine a greater range in heterozygote frequencies, exploits the pres- ence of mismatches at 0 and P in the heteroduplex
DNA. Phage harboring amber mutations in 0 or P
cannot replicate in a nonsuppressing bacterial host and therefore do not produce plaques on su- indicator strains. Phage that are heterozygous for each of these mutations cannot replicate either. However, correc- tion of the heterozygous phage to O+P+ by mismatch repair on the transcribed strand can restore growth in the su- indicator. Thus, the capacity to give rise to
0 + P + plaques on an su- indicator is another way to
detect the presence of heterozygous phage among
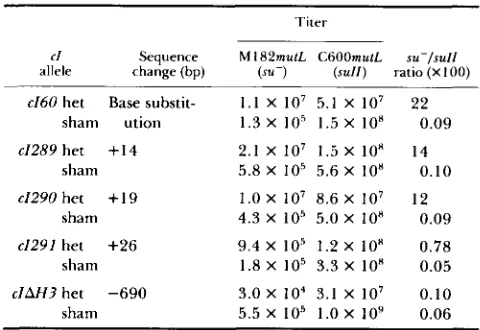
packaged products. Table 5 presents the results of titration of the packaged phage on an s u - indicator
(M182mutL). In order to correct for differences in
overall efficiency of packaging, the yield on the su- indicator was normalized to the phage titer detected
on a suppressing (sull) host. Phage packaged with het
DNA prepared from 26-bp and 690-bp deletion phage
show detectable differences in the yield o f ' O+P+
phage. We regard the O+P+ plaques seen with phage
packaged with sham DNA and the 690-bp deletion
het DNA as background, since their frequency is
sensitive to the number of phage plated and can
therefore be accounted for as the products of coinci-
TABLE 5
Results of titering packaged phage on su- and s d Z indicators
Titer
allele change (bp) (xu-) ( s u l l ) ratio (X 100)
C l Sequence M 182mut.L CGOOmutL su-/su/l
e160 het Base substit- 1 . 1 X 10' 5.1 X 1 O7 22
sham ution 1.3 X I O 5 1.5 X 10" 0.09
e1289 het + I 4 2.1 X IO7 1.5 X 10" 14
sham 5.8 X 10' 5.6 X 10" 0.10
e1290 het
+
19 1.0 X lo7 8.6 X I O 7 12sham 4.3 X IO5 5.0 X 10" 0.09
e1291 het +26 9.4 X 10' 1.2 X l o H 0.78
sham 1.8 X 10' 3.3 X 10" 0.05
c I U 3 het -690 3.0 x 104 3.1 X 107 0.10
sham 5.5 X I O 5 1.0 X IO9 0.06
dent infection. DNA harboring the 26-bp insertion
heterology gives rise to O+P+ plaques at a frequency approximately 8-fold higher than background.
Restriction analysis of packaged phage DNA: In
the experiments described above, phage were identi- fied as heterozygous for GI if they produced plaques containing a mixture of clear and turbid phage gen- otypes. T h e failure to recover c l heterozygotes among
phage packaged with DNA containing 26-bp and 690-
bp heterologies could be due either to a failure to
encapsidate DNA harboring these heterologies or a
failure of the packaged phage harboring these heter- ologies to produce plaques. For example, the presence
of these heterologies in the phage DNA may interfere
either with threading the DNA into the prohead during packaging or with the process of injecting the
DNA into a bacterial host. These two possibilities were distinguished by purifying phage packaged with the
26-bp and 690-bp het DNAs and then carrying out a
restriction analysis of the DNA isolated from the
packaged phage to determine the presence or absence
of the DNA heterologies. DNA from phage packaged
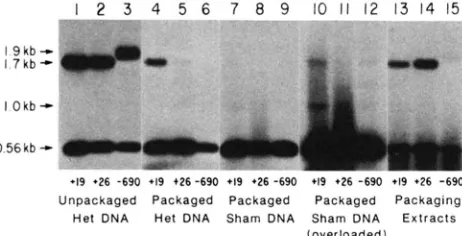
with the 19-bp heterology was included as a positive control. T h e results are shown in Figure 1.
Digests of unpackaged het DNA produce two bands
that hybridize to the cl/rexA probe. Since het DNA is
made by denaturing and reannealing DNA from a
mixture of c l + Pam80 and c l - Oam29 phage, it con- tains a mixture of reannealed parental and heterodu-
plex DNAs. With the HindIII-XhoI-MspI digestion
scheme, all reannealed parental DNAs produce a sin-
gle 0.56-kb band that hybridizes to the probe (Figure
2). In heteroduplex DNA containing a 19- or 26-bp
Packaging DNA Heterologies in X 9
I 2 3 4 5
-
6 7 8 9 1011 12 1 3 1 4 1 5 T" " . _
I 9 k b -
1.7 kb -C
l Okb-,
+l9 '26 -690 +l9 4 2 6 -690 +l9 '26 -690 *l9 126 -690 +l9 '26 -690
Unpackaged Packaged Packaged Packaged Packaging Het DNA Het DNA Sham DNA Sham DNA Extracts
(overloaded)
FIGURE 1.-Southern blot o f DNA from packaged phage and packaging extracts. Unpackaged het DNA and DNA isolated from packaged phage and packaging extracts was digested with HindlII, Xhol and Mspl and probed with the rexA/cl Hindlll-Hind111 frag- ment. In all of the samples o f packaging extract DNA, the parental band is smeared; this is also evident with DNA isolated from the extract alone (data not shown) and is therefore probably the con- sequence of endogenous DNA present in the extracts.
with a 690-bp single-stranded loop, which migrates to
the position expected of a 1.9-kb fragment. Digestion of the unpackaged het DNA in all three cases thus gives separate heteroduplex and parental bands, as seen in the autoradiogram. These parental and het- eroduplex bands also appear at equal intensities, cor- roborating the transfection results in which the het DNAs gave rise to approximately equal numbers of parental and heterozygous transfection products.
Digestion of DNA isolated from phage which were packaged with the 19-bp het DNA yields both a 0.56- kb (parental) band and a 1.7-kb (heceroduplex) band. There is roughly a 10-fold difference in the relative intensities of the parental and heteroduplex bands. This agrees well with the 8% frequency of c l hetero- zygotes observed among phage packaged with the 19- bp het DNA. DNA isolated from phage packaged with the 26-bp heterology produces a strong 0.56-kb (pa- rental) band, but only a very faint 1.7-kb (heterodu- plex) band. No heteroduplex band is visible in the case of the packaged 690-bp het DNA, but this obser-
vation is compromised by the poor recovery of pack- aged phage DNA as reflected in the low intensity of the parental band.
DNA isolated from the packaged sham controls
produces strong parental bands, but virtually no het- eroduplex bands, indicating that unpackaged hetero- duplex DNA was successfully removed by the phage purification steps. A faint 1.7-kb band, as well as an equally faint 1.0-kb band, can be seen in lanes over- loaded with digests of packaged sham control DNA. These t w o bands would be expected with a low level
of incomplete Hind111 digestion of the sham DNA
itself. I t is unlikely that the very faint heteroduplex band observed in the case of the packaged 26 bp het DNA is a partial restriction fragment, however, since the digest was clearly not overloaded, and the 1.0-kb partial restriction fragment is absent.
Digestion of the total DNA isolated from the 19-bp and 26-bp het DNA packaging extracts at the end of the 2-hr incubation gives both 0.56-kb and 1.7-kb bands at approximately the same relative intensities. Thus, there does not appear to be a selective loss of heteroduplex material during these in vitro packaging incubations. In contrast, the heteroduplex band is less intense relative to the parental band in the case of the 690-bp het DNA packaging extracts. This effect was also seen with DNA isolated from another packaging extract in which a different preparation of 690-bp het DNA was used (data not shown). T h e 690-bp het DNA also displayed a frequency of heterozygous products of transfection lower than that expected on the basis of random assortment of strands. These observations suggest the possibility that heteroduplex chromosomes harboring a 690-bp heterology are sub- ject to degradation.
DISCUSSION
Bacteriophage lambda chromosomes harboring cl
heterologies of up to 19 bp were packaged in vitro and found to produce heterozygous c l - / c I + phage
-Probe
cl289 or cl290
- M x A + + W ~ +
(+l9 bp or + 26 bp) + o ~ + o . 5 s ~ ~ - 1 . s * D - ! FIGURE 2.-Restriction Map of
Xcl Homoduplex and Heteroduplex
0. k - I . ? &
-
DNA. Both +19-bp and +26-bp in-Msp I Hind 111 Mspl sertions have XhoI linkers in the cl
Mspl HndIII Xhol
+
*
+*
W T Hind111 site. Parental DNA from
XclAH3 is deleted for the probe se-
quences. A restriction fragment har- boring the large heteroduplex loop created by the 690-bp deletion mi- grates aberrantly in agarose gels as a Msp I Hnd 111 HIM 111 Msp I
c'AM
-
'1.9 kb'
-
(- 690 bp) 0 4 5 k -1,Okb- MW I Msp I
Msp I Htrd Ill Msp I 1.9-kb fragment.
Parental Chromosomes
Heteroduplex
among the packaged products. Thus, DNA that in- cludes small heterologies can be packaged successfully. Phage packaged with DNA harboring a base pair mismatch or with DNA harboring heterologies of up to 19 bp yield similar frequencies of heterozygotes,
showing that phage DNA containing a heterology as
large as 19 bp is packaged nearly as efficiently as is phage DNA harboring only base pair mismatches. In contrast, the yield of heterozygotes is greatly reduced when a heterology of 26 bp is present. With a 690 bp heterology, no heterozygotes are observed above the background level of detection. T h e relative yield of packaged heterozygotes was assessed by measuring the frequency of heterozygosity at c l and heterozy- gosity at 0 and P in the packaged phage, as well as by
Southern analysis of packaged phage DNA. There
was a 10- to 20-fold reduction in the frequency of c l
heterozygotes and of 0-P heterozygotes packaged
with DNA harboring a 26-bp heterology as compared
to heterozygotes packaged with a 19-bp heterology. Southern analysis of packaged phage similarly shows evidence of a large reduction in the amount of het- erology-containing DNA when the size of the heter-
ology increases from 19 to 26 bp. Thus, DNA har-
boring a 26-bp heterology appears to be encapsidated much less efficiently than DNA with a modestly smaller heterology.
What is the basis of the difference in packaging efficiency of DNA containing a 19-bp heterology as compared to DNA containing a 26-bp heterology? One possibility is that the size of the larger heterology physically interferes with the packaging process. It would appear that during encapsidation, the phage DNA passes through the head-tail connector into the
prohead (KOCHAN and MURIALDO 1983; TSUI and
HENDRIX 1980), raising the possibility that structural
perturbations in the DNA duplex could block passage through the portal of the connector. Translocation of DNA into the phage T 4 prohead, for example, ap- pears to be blocked by the presence of branches in the phage DNA substrate, which must first be re-
moved by the action of T 4 endonuclease VI1 (MIZU-
UCHI et al. 1982; HSIAO and BLACK 1978; KEMPER
and JANZ 1976; KEMPER and BROWN 1976). T h e
dimensions of the portal would naturally limit the size of heterology that could be packaged, and this could account for the cut-off between packaging DNA with 19 bp and 26 bp heterologies. T h e diameter of the portal has been estimated to be similar to the width
of the DNA double helix (KOCHAN, CARRASCOSA and
MURIALDO 1984). It is therefore surprising that even
a 19-bp heterology can be accommodated in packag- ing. It may be that the portal is dilated during pack- aging, thus allowing the smaller heterologies to get through. In other variations of this model, large het- erologies could interfere with the packaging apparatus
or a packaging monitoring mechanism (SYVANEN and
YIN 1978; FEISS and WIDNER 1982); for example, if
terminase and/or some monitoring protein traverses the DNA during packaging, the 19-bp heterology
might be tolerated while a 26-bp heterology might
not. A larger portal size in the case of phage T4 could
perhaps accommodate the evidence of Mosrc and
POWELL (1985), of packaging of DNA with a 150-bp
heterology when the gene 49 (endonuclease VII)
product is disabled.
Another possibility to account for the boundary between 19-bp and 26-bp heterologies could be the presence of a DNA-binding protein in the crude pack- aging extract that recognizes and binds only heterol-
ogies of 26 bp or more. This bound protein could
physically prevent entry of the DNA into the prohead. It is also possible that the 26-bp heterology could be selectively repaired or degraded, but this seems un- likely since Southern analysis of DNA isolated from the packaging extract at the end of the two hour incubation does not indicate a selective loss of the 26-
bp heterology-containing restriction fragment. How-
ever, one cannot rule out the possibility that the larger heterologies are enzymatically nicked on one strand, since a nicked DNA fragment would migrate to the same position as the intact fragment in a gel. Evidence obtained with phage T 4 suggests that nicked DNA
substrates are not packageable (ZACHARY and BLACK
1981). This class of models, in which a DNA-binding protein selectively binds and/or nicks heterologies of
26 bp or more, seems less attractive than models
postulating that the larger heterologies physically ob- struct DNA translocation.
T h e in vitro packaging of each het DNA preparation
yields a lower frequency of heterozygotes among pack-
aged products than expected on the basis of transfec-
tion. For example, cZ60 het DNA was prepared by
denaturing and reannealing DNA from XcZ+ Pam80
with DNA from XcI60 Oam29; it therefore contains
heteroduplex molecules which would harbor three
base mismatches (at cZ60, Pam80 and Oam29), as well
as parental homoduplex chromosomes. As indicated by the frequency of
CZ
heterozygous products of trans- fection, the cI60 het DNA preparation had approxi- mately 42% heteroduplex molecules, as expected on the basis of random assortment of annealed strands. However, only 13% of the phage packaged with this DNA are heterozygous for cZ, suggesting that the mismatches may have interfered with the encapsida- tion of the heteroduplex molecules. Perhaps similarly, the presence of pyrimidine dimers in bacteriophage T 4 DNA appears to impair the in vivo encapsidationof the phage DNA, although some dimers are ulti-
mately packaged (ZACHARY and BLACK 1984). It
Packaging DNA Heterologies in h 1 1
sult of the disruption in the DNA duplex caused by
the mismatch, since a heterology of 19 bp, which
represents a much greater disruption, is packaged to nearly the same extent. It is also possible that heter- ologies, including mismatches, are targets of endonu- clease activity during incubation in the extract. Such an activity might cause the selective loss of heterodu- plex chromosomes. Another perhaps related possibil- ity could be the presence of a protein in the extract that binds to DNA mismatches, thereby impairing
encapsidation of the heteroduplex DNA. In these
models, all heteroduplexes would be impaired in pack-
aging to about the same extent, but homoduplex DNA
molecules, i.e., parental chromosomes, would be un-
impaired.
Although in vitro packaging has been the focus of the experiments described here, sotne interesting ge- netic implications are evident. First, as seen in Table
5 , phage packaged with DNA harboring heterologies
of 19 bp or less all give high titers of O+P+ plaques on the su- mutL indicator strain. T h e packaged hetero-
zygotes have mismatched 0 and P markers and cannot
give rise to O+P+ plaques unless corrected by mis-
match repair. One might have expected that the mutL
indicator, which is deficient in mismatch correction
(Lu, CLARK and MODRICH 1983; WAGNER et al. 1984),
would not give a high frequency of O+P+ plaques
from
0-P
heterozygotes. HUISMAN and Fox (1986),for example, observed only about 10% mismatch-
correction of heterozygotes in a mutL host that was not selective. However, in the case described here, the
0-P
heterozygotes cannot replicate, and this couldallow time for more extensive correction by the resid- ual mismatch repair activity in the mutL indicator. It was also observed that the O+P+ titers of packaged heterozygous phage were 1 0-fold lower on an su-mut+
indicator than on the su- mutL indicator. Although the level of mismatch correction would be expected
to be higher in the wild-type indicator, it is also likely that the repair tracts could be considerably longer in this host (FOX and RAPOSA 1983). Co-correction of
both 0 and P mismatches would reduce the yield of
O+P+ plaques.
I t is also surprising that packaged heterozygous phage display such high yields of recombinants on the
su- mutL indicator. For example, it would appear that
14% of the phage packaged with the 19-bp het DNA
yield O+P+ plaques. Transfection and the results of
Southern analysis indicate that about 10% of these
same packaged phage are heterozygous for GI. Thus,
it appears as though all of the heterozygous phage
would have to produce O+P’ plaques on the su- mutL
indicator. In other words, we are led to the unlikely possibility that all mismatch repair events restore the wildtype alleles on the transcribed strand. It is possible that the high frequency of O+P+ plaques is only ap-
parent for the following reason. T h e “frequency” of
O+P+ plaques among packaged phage reflects the plating efficiencies of the packaged phage on two different hosts, su- and sull. If the efficiency of plating
of the packaged phage were for some reason low on
the sull strain, then the ratio of the titers on selective and permissive indicators would not reflect the rela-
tive proportion of heterozygous phage among the
packaged products. It is possible, for example, that as the result of the action of nucleases in the in vitro
packaging extracts or the process of denaturation and
reannealing, DNA damages are present in the pack- aged phage. Suppression of the amber mutations in the permissive host could result in replication of phage DNA before the damages had been repaired, and result in loss of viability of the products. In the selec- tive host, however, no replication could occur until the 0 or P mismatch was corrected to wild type. T h e resulting delay in replication might permit repair of DNA damages, so that more phage DNA survives as an intact product. Thus, the sull titer may be a dis- torted reflection of the total yield of infectious pack- aged phage.
We have shown that small heterologies can be pack- aged in vitro to give viable heterozygous progeny and that the size of the heterology that can be packaged is subject to a boundary condition. T h e results of the
recombination experiments presented previously
(PEARSON and FOX 1988) corroborate these in vitro
packaging results, and suggest a discontinuity in pack- aging competence both in vivo and in vitro when the size of a heterology increases from 19 to 26 bp.
We thank BOB BOURRET and DAVE PEARSON for their help in preparing this and the accompanying manuscript. R.K.P. was sup- ported by training grant GM07287 from the National Institutes of Health and by a duPont Graduate Fellowship. This work was supported by National Institutes of Health grant A105388 to M.F.
LITERATURE CITED
BAZINET, C., and J. KING, 1985 The DNA translocating vertex of dsDNA bacteriophage. Annu. Rev. Microbiol. 3 9 109-129. BLACK, L., and M. SHOWE, 1983 Morphogenesis. pp. 219-245.
In: Bacteriophage T4, Edited by C. MATTHEWS, E. KUTTER, G. MOSIG and P. BERGET. American Society for Microbiology, Washington, D.C.
CHURCH, G . M., and W. GILBERT, 1984 Genomic sequencing. Proc. Natl. Acad. Sci. USA 81: 1991-1995.
EARNSHAW, W. C., and S. R. CASJENS, 1980 DNA packaging by the double-stranded DNA bacteriophages. Cell 21: 3 19-33 1 . FEISS, M., and W. WIDNER, 1982 Bacteriophage lambda DNA
packaging: Scanning for the terminal cohesive and site during packaging. Proc. Natl. Acad. Sci. USA 79: 3498-3502. FOX, M. S., 1978 Some features of genetic recombination in
procaryotes. Annu. Rev. Genet. 12: 47-68.
Fox, M. S., and S. RAPOSA, 1983 Mismatch repair. pp. 333-335. In: Cellular Responses to DNA Damages, Edited by E. C. FRIED
BERG and B. A. BRIDGES. Alan R. L i s , New York.
HSIAO, C. L., and L. W. BLACK, 1978 Head morphogenesis of bacteriophage T4.111. The role of gene 20 in DNA packaging. Virology 91: 26-38.
products of bacteriophage lambda recombination. Genetics
KEMPER, B., and D. T. BROWN, 1976 Function of gene 49 of
bacteriophage T4. 11. Analysis of intracellular development and the structure of very fast-sedimenting DNA. J. Virol. 18:
KEMPER, B., and E. JANZ, 1976 Function of gene 49 of bacterio- phage T4. I. Isolation and biochemical characterization of very fast-sedimenting DNA. J. Virol. 18: 992-999.
KOCHAN, J., and H. MURIALDO, 1983 Early intermediates in bac- teriophage lambda prohead assembly. 11. Identification of bio- logically active intermediates. Virology 131: 100-1 15. KOCHAN, J., J. L. CARRASCOSA and H. MURIALW, 1984
Bacteriophage lambda preconnectors. Purification and struc- ture. J. Mol. Biol. 174 433-447.
LICHTEN, M. J., and M. S. FOX, 1983 Detection of non-homology- containing heteroduplex molecules. Nucleic Acids Res. 11:
Lu, A. L., S. CLARKand P. MODRICH, 1983 Methyldirectedrepair of DNA base-pair mismatches in vitro. Proc. Natl. Acad. Sci.
MANDEL, M., and A. HIGA, 1970 Calciumdependent bacterio- phage DNA infection. J. Mol. Biol. 53: 159-162.
MANIATIS, T., E. F. FRITSCH and J. SAMBROOK, 1982 Molecular
Cloning: A Laboratory Manual. Cold Spring Harbor Laboratory,
Cold Spring Harbor, N.Y.
MCMILIN, K. D., and V. RUSSO, 1962 Maturation and recombi- nation of bacteriophage lambda DNA molecules in the absence of DNA duplication. J. Mol. Biol. 6 8 49-55.
MEINKOTH, J., and G. WAHL, 1984 Hybridization of nucleic acids 112 409-420.
1000-1015.
3959-397 1.
USA 8 0 4639-4643.
immobilized on solid supports. Anal. Biochem. 138 267-284. MIZUUCHI, K., B. KEMPER, J. HAYS and R. A. WEISBERG, 1982 T 4
endonuclease VI1 cleaves holliday structures. Cell 2 9 357- 365.
MOSIG, G., and D. POWELL, 1985 Heteroduplex loops are pack- aged in gene 49 (endonuclease VII) mutants of bacteriophage T5. Abst. Ann. ASM Meet. p. 209.
PEARSON, R. K., and M. S. Fox, 1988 Effects of DNA heterologies on bacteriophage X recombination. Genetics 118 000-000.
RADDING, C. M., 1982 Homologous pairing and strand exchange in genetic recombination. Annu. Rev. Genet. 1 6 405-437. ROSENBERG, S. M., M. M. STAHL, I. KOBAYASHI and F. W. STAHL,
1985 Improved in vitro packaging of coliphage lambda DNA: a one-strain system free from endogenous phage. Gene 3 8
165-175.
SYVANEN, M., and J. YIN, 1978 Studies of DNA packaging into the heads of bacteriophage lambda. J. Mol. Biol. 126 333- 346.
Tsur, L., and R. W. HENDRIX, 1980 Head-tail connector of bac- teriophage lambda. J. Mol. Biol. 142: 419-438.
WAGNER, R., C. DOHET, M. JONES, M. P. DOUTRIAUX, F. HUTCH-
INSON and M. RADMAN, 1984 Involvement of Escherichia coli mismatch repair in DNA replication and recombination. Cold Spring Harbor Symp. Quant. Biol. 4 9 61 1-615.
ZACHARY, A., and L. W. BLACK, 1981 DNA ligase is required for encapsidation of bacteriophage T 4 DNA. J. Mol. Biol. 149
ZACHARY, A., and L. BLACK, 1984 UV irradiation impairs in vivo encapsidation of bacteriophage T 4 DNA. J. Virol. 5 0 293- 300.
641-658.