Copyright © 2003, American Society for Microbiology. All Rights Reserved.
Method for Quantitative Detection and Presumptive Identification of
Group B Streptococci on Primary Plating
Søren Mose Hansen and Uffe B. Skov Sørensen*
Department of Medical Microbiology and Immunology, University of Aarhus, DK-8000 Aarhus C, Denmark
Received 24 July 2002/Returned for modification 15 October 2002/Accepted 27 November 2002
Maternal prenatal screening for group B streptococci (GBS) followed by offering of intrapartum
chemopro-phylaxis to carriers is one of the strategies used to reduce the incidence of neonatal early-onset GBS infections.
Culturing of vaginal and anorectal swab specimens in selective broth is the screening procedure recommended
by the Centers for Disease Control and Prevention. This technique is sensitive; it does not, however, allow
either evaluation of the degree of colonization or detection of cocolonization with several GBS clones. We have
examined the carriage rate and population dynamics of GBS in a group of Danish women during pregnancy
and 1 year after delivery using a new detection method. In the present paper we describe a mixed blood agar
medium (MB agar) that identifies GBS in the primary cultures by detection of a double hemolysis pattern
consisting of characteristic, large zones of partial hemolysis (“CAMP zones”) and of narrow zones of complete
hemolysis. The MB agar was at least as sensitive as culturing in selective broth for detection of GBS in vaginal
and anorectal swab specimens, and GBS strains could be identified directly on the primary plate due to the
CAMP zones without the need for subculturing. The carriage rate of GBS in a group of Danish women was
found to be more than 30%, a figure considerably higher than the rate that was reported previously.
Streptococcus agalactiae
(group B streptococcus [GBS]) is
the principal cause of neonatal infections in industrialized
countries. A significant decline in the prevalence of neonatal
GBS infections has been observed in the United States as a
result of implementation of preventive measures (7, 28). These
include increased use of chemoprophylaxis during labor in
women at high risk for transmitting the infection to their
new-borns (29). Risk-based or screening-based strategies are
rec-ommended by the Centers for Disease Control and Prevention
to prevent GBS disease (6). Collection of swab specimens at 35
to 37 weeks of gestation is suggested for the screening-based
approach. Rectal and vaginal swab specimens are inoculated
together in a selective broth for 18 to 24 h. The broth is
subcultured on sheep blood agar for another 18 to 24 h, and
suspected colonies are finally examined by conventional
micro-biological techniques. This procedure is not applicable to
semi-quantitative evaluation of colonization intensity, and it is not
feasible for demonstration of colonization with multiple
clones.
For a longitudinal study of the rate of carriage and
popula-tion dynamics of GBS in a cohort of pregnant women, we
needed a medium that compensates for these shortcomings.
We developed a solid medium based on a modification of the
classical CAMP reaction that allows easy detection of
individ-ual colonies of GBS in primary cultures.
GBS generate a unique protein, the so-called CAMP factor,
which interacts with the plasma membrane of red blood cells
(RBCs) and other cell types. CAMP factor causes hemolysis of
sheep RBCs (SRBCs) that have been altered by
Staphylococ-cus aureus

-toxin (sphingomyelinase). This phenomenon is
designated the CAMP reaction, which refers to the authors
Christie, Atkins, and Munch-Petersen (8). CAMP reaction-like
reactions have also been observed after the combined actions
of some other bacterial products on blood, and cohemolysis is
therefore a more adequate expression for a hemolytic reaction
induced by the synergistic actions of two different compounds,
as discussed below. The CAMP factor of GBS has been
char-acterized and has been renamed protein B. It has been
sug-gested that the target of protein B is ceramide (
N
-acyl-sphin-gosine) (4), i.e., a membrane lipid generated from sphingomyelin
by sphingomyelinase cleavage. Binding of protein B causes
disorganization of the lipid bilayer of the cell membrane to an
extent that results in cell lysis (4). The in vivo function of
protein B has not been disclosed yet; however, the hemolytic
activity seems to be an epiphenomenon (19, 27). Binding of
this protein to the Fc region of immunoglobulins has been
demonstrated (12, 19), and the protein seems to be lethal to
rabbits and mice (30). Protein B seems to be essential for
S.
agalactiae
, as the
cfb
gene encoding this protein (24) was found
to be universally present in the genomes of 162 GBS strains
from different geographic areas (20). Furthermore, 96 to 99%
of all human GBS isolates have been found to be positive by
the CAMP reaction (11, 14, 23, 25). Nearly all group A
strep-tococci (
Streptococcus pyogenes
) and
Streptococcus uberis
strains possess equivalent genes named
cfa
and
cfu
,
respec-tively (15, 18). The gene may also be present in some other
streptococcal species (16, 18). The cohemolysis observed with
other species is due to substances different from protein B. The
CAMP reaction-like reaction of
Listeria monocytogenes
is
caused by listeriolysin O (26), and that of
Actinobacillus
pleu-ropneumoniae
is caused by an RTX toxin (17).
Protein B may cause lysis of

-toxin-modified RBCs, as
mentioned above; however, only cells containing more than 45
mol% of sphingomyelin in the plasma membrane are sensitive
to the combined actions of the staphylococcal toxin and
pro-* Corresponding author. Mailing address: Department of Medical
Microbiology and Immunology, Bartholin Building, University of
Aar-hus, DK-8000 Aarhus C, Denmark. Phone: 45 89421740. Fax: 45
86196128. E-mail: [email protected].
1399
on May 15, 2020 by guest
http://jcm.asm.org/
tein B (31). Thus, the CAMP reaction is seen on agar plates
prepared from bovine RBCs and SRBCs, whereas RBCs from
humans, rabbits, and guinea pigs and horse RBCs (HRBCs)
are not lysed (8, 31).
We have taken advantage of the unique feature of protein B
as the indicative principle in a new blood agar medium that can
be used for the easy, sensitive, and specific detection and
enu-meration of GBS in primary cultures of complex specimens
such as anorectal and vaginal swab specimens. The agar plates
described in this article contained a mixture of SRBCs and
HRBCs, which were made sensitive to the action of protein B
by pretreatment with sphingomyelinase. The method was
suc-cessfully applied in a longitudinal study of the GBS carrier
status of a group of Danish women during pregnancy and after
delivery (unpublished data). Compared with five other
tech-niques for GBS detection, the new medium was found to be at
least as sensitive and easier to use, since subculturing was
usually unnecessary. The option for semiquantitative
enumer-ation of GBS and detection of cocolonizenumer-ation with several
clones of GBS is an additional advantage of the mixed blood
agar (MB agar) plates.
MATERIALS AND METHODS
Bacterial strains from collections.Two strains of GBS from our own collection were used for control purposes. Strain RH955 is beta-hemolytic and yields a positive CAMP reaction. Strain RH511A also yields a positive CAMP reaction but is nonhemolytic. Staphylococcal-toxin was prepared from cultures of anS. aureusstrain (SSI AB2775) from the Department of Clinical Microbiology, Statens Serum Institut, Copenhagen, Denmark. The toxin was used for treatment of MB agar plates (see below).
Preparation of crude staphylococcal-toxin.Partly purified-toxin (sphingo-myelinase) was prepared from a Todd-Hewitt broth culture (no. CM189; Oxoid Ltd., Basingstoke, United Kingdom) of theS. aureus strain as follows. The culture was incubated for 24 h at 37°C (optimum, 24 to 28 h), and the bacteria were removed by centrifugation (1,750⫻g, 30 min). The clear supernatant was filter sterilized (pore size, 0.45m), and 104 g of (NH4)2SO4per liter was added.
After the mixture was stirred with a magnetic stirrer for 1 h and allowed to settle for 30 min at room temperature, the solution was centrifuged (10,000⫻g) for 20 min. The pellet of the precipitate was discharged, and another 104 g of (NH4)2SO4was added to the total volume of the supernatant. After stirring of
the mixture for 1 h, the mixture was kept at room temperature. On the next day the solution was centrifuged (10,000⫻g) for 20 min. The second supernatant was discharged, and the pellet of crude toxin was dissolved in 20 ml of sterile phosphate-buffered saline (PBS). The toxin concentrate was dialyzed four times against 2 liters of PBS for 24 h at 5°C. Finally, the concentrated toxin stock solution was filter sterilized (pore size, 0.45m) and stored at 5°C until it was used. The toxin preparation was stable for several months when stored at 5°C. The sphingomyelinase activity of the toxin stock was determined by titration. The whole surfaces of a number of MB agar plates (see below) were treated with 200 l of twofold serial dilutions of the toxin stock in PBS. A CAMP reaction-positive strain of GBS (strain RH955) was streaked onto each of the toxin-treated plates before they were incubated overnight at 37°C. The plates were inspected for detection of two different hemolysis phenomena around colonies of GBS (double hemolysis). The action of-hemolysin caused narrow zones of complete hemolysis, and protein B caused larger zones of partial hemolysis (Fig. 1). The highest toxin dilution that gave optimal double hemolysis around the bacterial colonies was considered the end point. The ready-made-toxin solu-tion used for pretreatment of the MB agar plates (see below) was prepared so that it was two times more concentrated than the dilution determined by end-point titration. The use of sphingomyelinase from commercial sources may be a convenient alternative to the in-house preparation of toxin. However, we have not tested such preparations.
MB agar plates.Modified CAMP reaction plates containing mixed blood were prepared with a double layer of agar. One liter of agar contained 40 g of blood agar base (no. CM854; Oxoid), 2 g of sodium pyruvate, 0.247 g of MgSO4, and
50 mg ofL-cysteine hydrochloride monohydrate. The ingredients were mixed
during heating to the boiling point and then autoclaved for 15 min at 121°C. The
medium was cooled and kept in a water bath at 50°C. For preparation of the top layer, 2.5% (vol/vol) packed SRBCs and 2.5% (vol/vol) packed HRBCs were added to the 50°C agar base and mixed. The blood cells were washed three times before use in PBS (0.1 M NaCl, 0.05 M phosphate buffer [pH 7.4]) and packed by centrifugation (at 500⫻gfor 5 min). Each plate (diameter, 9 cm) was made of a lower layer of agar base without blood cells (25 ml). After solidification, a top layer of agar base (20 ml) containing 5% mixed blood cells was added. Hemolysis is more easily seen on the relatively thin blood-containing top layer. Untreated plates were stable and could be stored for several weeks in a refrig-erator. Before use, the MB agar plates were pretreated with staphylococcus -toxin, as described below. After treatment with the staphylococcal toxin, the plates were somewhat sensitive to lowering of the temperature. To avoid spon-taneous hemolysis, these plates were kept at room temperature and used on the same day.
g-MB agar plates.MB agar plates containing gentamicin (g-MB agar plates) in both the lower and the top layers were prepared by adding 1 ml of a filter-sterilized (pore size, 0.45m) stock solution (4 mg per ml of H2O) of gentamicin
sulfate (658g of gentamicin base per mg; no. G 6896; Sigma-Aldrich, St. Louis, Mo.) per liter of agar base at 50°C (see above).
ng-MB agar plates.MB agar plates containing both nalidixic acid and genta-micin (ng-MB agar plates) in both the lower and the top layers were prepared by adding 1 ml of a filter-sterilized (pore size, 0.45m) stock solution (8 mg per ml of H2O) of gentamicin sulfate (no. G 6896; Sigma-Aldrich) and 1 ml of a
FIG. 1. Close-up photos of large CAMP zones surrounding
colo-nies of
S. agalactiae
on MB agar pretreated with staphylococcal

-toxin
1 h before they were streaked with bacteria. (A) A typical
beta-hemo-lytic strain of GBS, strain RH955; (B) a nonhemobeta-hemo-lytic strain, strain
RH511A. The plates were incubated overnight at 37°C (see text for
details). Arrows: 1, bacterial colonies; 2, zones of partial hemolysis
around colonies of both bacterial strains; the SRBCs are lysed in these
areas due to the interaction with protein B generated by the bacteria,
whereas the HRBCs are left intact, as these cells are insensitive to
protein B; 3, after removal of the bacterial colony, a narrow spot of
complete hemolysis, in which both kinds of RBCs are lysed due to the
action of

-hemolysin, is seen in the middle of the CAMP zone; 4, as
for arrow 3, except that the bacterial colonies were left in place;
beta-hemolysis is seen as thin clear rims around the bacterial colonies;
5 and 6, no beta-hemolysis appears around the colonies of strain
RH511 or after removal of a colony.
on May 15, 2020 by guest
http://jcm.asm.org/
filter-sterilized (pore size, 0.45m) stock solution (15 mg per ml of 0.1 N NaOH) of nalidixic acid (no. N 8878; Sigma-Aldrich) per liter of agar base at 50°C (see above).
Pretreatment of MB agar plates with-toxin.A total of 200l of sterile ready-made-toxin solution (see above) was dispersed evenly with a plastic Drigalski spatula over the surface of the MB, g-MB, and ng-MB agar plates before use. The plates were kept at room temperature until the surfaces had dried (30 to 60 min).
Regular blood agar plates.Horse blood (5%) agar plates were purchased from the Statens Serum Institut.
Selective broth.Todd-Hewitt broth (no. 189; Oxoid) was supplemented with 8.0 mg of gentamicin sulfate and 15 mg of nalidixic acid per liter (6). Cotton-stoppered test tubes containing 5 ml of this medium were autoclaved for 15 min at 121°C. After the swabs were streaked on the different solid media they were placed in this selective broth, mixed carefully on a vortex mixer, and incubated overnight at 37°C. A total of 10l of broth from each of the test tubes was streaked on regular 5% blood agar plates, and the plates were incubated over-night at 37°C.
Bacterial identification.Suspected isolates of GBS were identified by a com-bination of standard tests (21). The strains were Gram stained, tested for catalase activity, and examined by the traditional CAMP test on CAMP test plates (Statens Serum Institut). The strains were examined for the presence of the group B antigen by latex agglutination (no. ZL52; Streptex; Murex Biotech Ltd., Dartford, United Kingdom).
Clinical samples.A cohort of 77 healthy women was monitored in a longitu-dinal study (unpublished data) from the 19th week of gestation until shortly before delivery at the Department of Obstetrics, Aarhus University Hospital, Skejby, Denmark. All pregnancies were normal, and no complications due to GBS were observed. The project was initiated in January 1999 with approval from the Ethics Committee, County of Aarhus, Denmark, and after informed consent had been provided by all volunteers. The participants were instructed in the technique for obtaining swab specimens at home. Carbon-containing cotton swabs on plastic sticks (Statens Serum Institut) were used for the sampling. Vaginal and anorectal swab specimens were obtained at different times during the pregnancy and again 1 year after delivery (spring 2001). Each swab was immediately placed in a tube containing Stuart’s transport medium (Statens Serum Institut) and shipped to the laboratory by regular mail. All swabs were examined within 36 h after the samples were taken. For the present study, more than 300 paired samples were examined on up to five different media (Table 1). The swabs were inoculated on the solid media as described below for the ng-MB agar plates. Some samples were not examined on all media (exact figures are given elsewhere in the text).
Evaluation of intensity of GBS colonization on ng-MB agar plates. Presump-tive identification of GBS colonies is possible directly on the ng-MB agar plates (see above). These plates were therefore used for semiquantitative evaluation of the degree of colonization, as follows. Upon arrival in the laboratory in Stuart’s transport medium, each swab specimen (vaginal or anorectal) was rolled and rubbed over an area 2 by 3 cm near the edge of an ng-MB agar plate. The inoculum was streaked from this area five to six times with one side of a sterile 10-l plastic inoculation loop. To obtain well-separated colonies, spreading was continued by making three sets of streaks perpendicular to the previous streak by using the other side of the same loop. One person performed the inoculation of the agar plates to make it as reproducible as possible. Colonization intensity was evaluated semiquantitatively and blindly for each plate after incubation for 18 h at 37°C in an atmosphere containing 5% CO2. Growth of GBS in streaking area
one or two only was considered light colonization, while growth of GBS in one or more of the following streaking areas was considered moderate to heavy colonization. Samples without detectable growth of GBS were considered negative.
Reagents.All chemicals were of analytical grade and were purchased from Merck (Darmstadt, Germany) or Sigma-Aldrich.
Statistics.SigmaStat statistical software (version 1.0; Jandel Corporation, San Rafael, Calif.) was used for analysis of contingency tables by McNemar’s test.
RESULTS
Initial experiments revealed large zones of hemolysis around
colonies of GBS grown on sheep blood agar plates pretreated
with staphylococcal

-toxin. In contrast, only narrow zones of
hemolysis were seen on plates not pretreated with

-toxin. The
hemolysis around colonies of GBS grown on horse blood agar
plates was unaffected by

-toxin pretreatment. On this medium
the bacteria generated only narrow zones of hemolysis. A
dou-ble pattern of partial and complete hemolysis was seen when
the bacteria were cultured on

-toxin-treated agar plates made
with mixed SRBCs and HRBCs (MB agar; Fig. 1). The
com-position of the MB agar plates was optimized in a number of
experiments to promote the most clear-cut reactions for GBS
strains (the recipe is given in the Materials and Methods
sec-tion). On this MB agar, large zones (5 to 8 mm) of partial
hemolysis (“CAMP zones”) and narrow zones (2 to 3 mm) of
complete hemolysis (beta-hemolysis) were generated by the
GBS isolates (Fig. 1A). Nonhemolytic isolates also generated a
large CAMP zone (Fig. 1B), while beta-hemolytic CAMP
re-action-negative isolates did not (data not shown). Experiments
with
S. pyogenes
strains showed that the beta-hemolytic zone
dominates over the CAMP zone for these strains; thus, they
did not look like GBS strains.
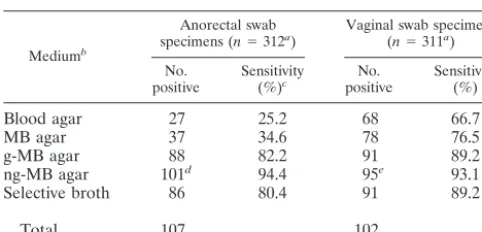
[image:3.603.43.284.88.204.2]More than 300 paired anorectal and vaginal swab samples
were cultured on five different media (Table 1). GBS were
easily detected on the MB agar plates by the presence of the
characteristic double hemolysis around the colonies (Fig. 1A),
and the sensitivity of this medium was found to be considerably
higher than that of regular blood agar plates (Table 1). The
nonselective plates of both media were, however, often
over-grown by the commensal flora in the samples, causing a low
detection rate, especially for the anorectal swab samples.
An-tibiotics were therefore added to the MB agar in order to
improve the isolation of GBS. The best results were obtained
by adding a combination of gentamicin and nalidixic acid to the
agar (Table 1), even though the colonies of GBS appeared
smaller on ng-MB agar medium than on medium containing
gentamicin alone. The sensitivity of the selective MB agar for
detection of GBS in both vaginal and anorectal swab samples
was found to be at least as high as that when a selective broth
was used (Table 1). Full agreement was found between the
tentative diagnosis and the results of the supplementary tests
TABLE 1. Frequency of GBS in swab samples cultured on
different media
Mediumb
Anorectal swab
specimens (n⫽312a) Vaginal swab specimens(n⫽311a) No.
positive Sensitivity(%)c positiveNo. Sensitivity(%)
Blood agar
27
25.2
68
66.7
MB agar
37
34.6
78
76.5
g-MB agar
88
82.2
91
89.2
ng-MB agar
101
d94.4
95
e93.1
Selective broth
86
80.4
91
89.2
Total
107
102
aTotal number of swabs tested on all media.
bBlood agar, 5% blood agar plates; MB agar, agar plates containing SRBCs and HRBCs and pretreated with staphylococcal-toxin; g-MB agar, MB agar with gentamicin sulfate; ng-MB agar, MB agar with nalidixic acid and gentamicin sulfate; selective broth, Todd-Hewitt broth with nalidixic acid and gentamicin sulfate; total, number of swabs yielding growth of GBS on one or more of the media used.
cSensitivity, percentage of swabs that tested positive on the medium of the total number of swabs that tested positive on one or more of the media used.
dThe number of anorectal swab specimens positive on ng-MB agar was sig-nificantly higher (P⬍0.001) than the number of specimens positive in selective broth (see text).
eThe number of vaginal swab specimens positive on ng-MB agar was not significantly different (P ⫽0.30) from the number of specimens positive in selective broth (see text).
on May 15, 2020 by guest
http://jcm.asm.org/
when the typical double hemolysis was observed on the MB
agar plates. Neither
S. pyogenes
nor other protein B-generating
streptococci were isolated from the samples.
More than one-third of all pairs of samples were found to be
positive for GBS (36.5%; 131of 358) when MB agar was used.
There was a high concordance (93%) between the results
ob-tained from the analysis of pairs of vaginal and anorectal swab
specimens (Table 2; Fig. 2). No difference in the number of
positive vaginal swab specimens compared to the number of
positive anorectal swab specimens was seen (
P
⫽
0.31), but 5 to
10% more carriers were found when both vaginal and
anorec-tal swab specimens were examined instead of when only one
swab specimen was examined from each person. An increased
rate of detection of GBS was achieved when samples were
obtained from both the vaginal and the anorectal sites, as
opposed to when samples were obtained from either site alone,
but there was no difference in rates of detection among
sam-pling sites. This finding has also been reported by others (22).
The number of anorectal swab specimens positive for GBS was
significantly higher when ng-MB agar was used for cultivation
than with cultivation in selective broth (355 swab specimens
were tested on these two media; of these, 91 were positive on
both media, 240 were negative on both media, 21 were positive
on ng-MB agar and negative in selective broth, and 3 were
negative on ng-MB agar and positive in selective broth [
P
⬍
0.001]). The number of GBS-positive vaginal swab specimens
detected when ng-MB agar was used for cultivation was,
how-ever, not significantly different from the number detected when
selective broth was used (356 swab specimens were tested on
these two media; of these, 96 were positive on both media, 245
were negative on both media, 10 were positive on ng-MB agar
and negative in selective broth, and 5 were negative on ng-MB
agar and positive in selective broth [
P
⫽
0.30]).
As mentioned above, a strong correlation was found
be-tween the degree of colonization detected in the two swab
samples (vaginal and anorectal) in a sample pair. For more
than 90% of GBS-negative vaginal swab samples, the
corre-sponding anorectal swab sample was negative as well (Fig. 2;
Table 2). In contrast, among the GBS-positive vaginal swab
samples, only 4 to 5% of the corresponding anorectal swab
samples were negative. A clear tendency toward the detection
of heavier colonization of the vaginal swab samples with
in-creasing colonization of the corresponding anorectal swab
samples was also seen (Fig. 2). Vaginal colonization with GBS
therefore reflects the anorectal colonization density to a high
degree.
DISCUSSION
The CAMP test is a standard procedure for the presumptive
identification of
S. agalactiae
(GBS) (9, 11). In the traditional
assay, a

-toxin-producing
S. aureus
strain is streaked across a
blood agar plate made with SRBCs. A number of suspected
strains of GBS are then streaked on the same plate
perpen-dicular to the streak made with
S. aureus
. For most GBS strains
an arrowhead-shaped hemolysis zone appears near the
inter-section with the
S. aureus
strain after incubation of the plate.
This is due to the cohemolysis caused by the concerted action
of

-toxin from the
S. aureus
strain and protein B from the
strains of GBS. While this CAMP test is useful for the
pre-sumptive identification of pure cultures of suspected strains of
GBS (9), the classical assay is not applicable for direct
detec-tion of GBS in primary cultures of clinical samples. Different
modifications of the CAMP test have been developed for this
purpose (3, 10). However, none of these methods are optimal
for routine examination of a large number of samples, since the
procedures are relatively complicated. Others have used sheep
blood agar plates conditioned with sterile

-toxin-containing
supernatant from an
S. aureus
culture for detection of CAMP
test-positive GBS (1), but such agar plates cannot distinguish
the CAMP reaction and ordinary

-hemolysis.
The Centers for Disease Control and Prevention
recom-mended a procedure for screening for carriage of GBS that
requires culture of anorectal and vaginal swab specimens in a
selective broth which, after incubation, is subcultured on blood
agar plates; the isolates are eventually identified by serological
methods (6). This procedure is specific and sensitive but is
somewhat laborious, costly, and time-consuming and does not
allow quantitative evaluation and proportional isolation of the
individual strains in one sample.
[image:4.603.43.286.90.144.2]The selective MB agar has several advantages, although
preparation of the MB agar plates is more complicated than
preparation of the traditional plates. Culturing on MB agar
was found to be at least as sensitive as culturing in a selective
FIG. 2. Correlation between growth densities of
S. agalactiae
(GBS) in 310 paired vaginal and rectal swab samples from 77 pregnant
women. Vaginal samples are grouped on the abscissa according to
density of growth of GBS on ng-MB agar plates. The densities of
growth of GBS from the corresponding rectal swab samples were
recorded as negative (open bars), slight (hatched bars), and moderate
to heavy (solid bars) (see text for details).
TABLE 2. Frequency of GBS in 358 pairs of anorectal and vaginal
swab specimens cultured on ng-MB agar
Result for anorectal swab specimens
No. (%) of vaginal swab specimens
Positive Negative
Positive
107 (30)
15 (3)
Negative
9 (4)
227 (63)
on May 15, 2020 by guest
http://jcm.asm.org/
[image:4.603.49.280.458.656.2]broth, with a sensitivity of over 90%. The primary solid media
can be inspected directly within 18 h, and reculturing is rarely
needed, since typical colonies can be identified as GBS with a
high degree of confidence on the basis of the characteristic
hemolysis phenomenon. The density of GBS colonization can
therefore be estimated semiquantitatively. We found a clear
correlation between the colonization densities of GBS in
paired vaginal and anorectal samples by use of ng-MB agar
plates (Fig. 2). Furthermore, individual bacterial colonies can
be identified on the primary plates and may be selected for
further examination (e.g., for antibiotic resistance testing,
DNA analysis, or serotyping).
Some bacterial species other than GBS may exhibit a
posi-tive CAMP reaction, as mentioned above, but these species do
not constitute a diagnostic problem when MB agar is used for
cultivation of human clinical specimens. On this medium the
hemolytic pattern of group A streptococci differs from that of
GBS (see above).
S. ubris
is not associated with humans, and in
contrast to most GBS strains, strains of this species are not
beta-hemolytic (21).
A. pleuropneumoniae
is found only in pigs.
Occasionally, stool samples may contain
L. monocytogenes
(2),
and colonies of this species resemble colonies of
beta-hemo-lytic streptococci on blood agar. However, L
. monocytogenes
is
sensitive to gentamicin and will, therefore, not appear on the
selective MB agar plates. Furthermore, differentiation from
GBS is easily accomplished by demonstrating the positive
cata-lase reaction of
L. monocytogenes.
Among a group of 77 pregnant women, we found 21% to be
persistent carriers of GBS and an additional 27% to be
tran-sient carriers of GBS. These rates are considerably higher than
the rates previously reported for a large group of Danish
preg-nant women (13), in which only 8 to 15% carriers of GBS were
found. Other recent rates of carriage are not available from
Denmark. Our prevalence findings based on the use of
selec-tive MB agar are similar to those recently reported for younger
women from the United States. In a prevalence study in which
the selective broth technique was used, 34% of female
univer-sity students were found to be colonized with GBS (5), and
among a group of pregnant women in a prospective cohort
study, 28% were found to be colonized with GBS (22).
ACKNOWLEDGMENT
This work was supported by grant 9900316 kg/mp from the Danish
Medical Research Agency.
REFERENCES
1. Anthony, B. C., D. M. Okada, and C. J. Hobel.1978. Epidemiology of group B streptococci: longitudinal observations during pregnancy. J. Infect. Dis.
137:524–530.
2. Armstrong, D.1995.Listeisteria monocytogenes, p. 1880–1885.InG. L. Man-dell, J. E. Bennett, and R. Dolin (ed.), ManMan-dell, Douglas and Bennett’s principles and practice of infectious diseases. Churchill Livingstone, New York, N.Y.
3. Bae, B. H., and E. J. Bottone.1980. Modified Christie-Atkins-Munch-Pe-tersen (CAMP) test for direct identification of hemolytic and non-hemolytic group B streptococci on primary plating. Can. J. Microbiol.26:539–542. 4. Bernheimer, A. W., R. Linder, and L. S. Avigad.1979. Nature and
mecha-nism of action of the CAMP protein of group B streptococci. Infect. Immun.
23:838–844.
5. Bliss, S. J., S. D. Manning, P. Tallman, C. J. Baker, M. D. Pearlman, C. F. Marrs, and B. Foxmann.2002. Group BStreptococcuscolonization in male and non-pregnant female university students: a cross-sectional prevalence study. Clin. Infect. Dis.34:184–190.
6. Centers for Disease Control and Prevention.1996. Prevention of perinatal group B streptococcal disease: a public health perspective. Morb. Mortal. Wkly. Rep.45:1–24.
7. Centers for Disease Control and Prevention.2000. Early-onset group B streptococcal disease—United States, 1998–1999. Morb. Mortal. Wkly. Rep.
49:793–796.
8. Christie, R., E. Atkins, and E. Munch-Petersen.1944. A note on a lytic phenomenon shown by group B streptococci. Aust. J. Exp. Biol. Med. Sci.
22:197–200.
9. Darling, C. L.1975. Standardization and evaluation of the CAMP reaction for the prompt, presumptive identification of Streptococcus agalactiae (Lancefield group B) in clinical material. J. Clin. Microbiol.1:171–174. 10. DiPersio, J. R., J. E. Barrett, and R. L. Kaplan.1985. Evaluation of the
Spot-CAMP test for the rapid presumptive identification of group B strep-tococci. Am. J. Clin. Pathol.84:216–219.
11. Facklam, R. R., J. F. Padula, E. C. Worthham, R. C. Cooksey, and H. A. Rountree.1979. Presumptive identification of group A, B, and D streptococci on agar plate media. J. Clin. Microbiol.9:665–673.
12. Fehrenbach, F. J., D. Jurgens, J. Ruhlmann, B. Sterzik, and M. O¨ zel.1988. Role of CAMP-factor (protein B) for virulence. Zentbl. Bakteriol. Para-sitenkd. Infektkrankh. Hyg. Abt. 1 Orig. Suppl.17:351–357.
13. Feikin, D. R., P. Thorsen, S. Zywicki, M. Arpi, J. G. Westergaard, and A. Schuchat.2001. Association between colonization with group B streptococci during pregnancy and preterm delivery among Danish women. Am. J. Ob-stet. Gynecol.184:427–433.
14. Fuchs, P. C., C. Christy, and R. N. Jones.1978. Multiple-inocula (replicator) CAMP test for presumptive identification of group B streptococci. J. Clin. Microbiol.7:232–233.
15. Gase, K., J. J. Ferretti, C. Primeaux, and W. M. McShan.1999. Identifica-tion, cloning, and expression of the CAMP factor gene (cfa) of group A streptococci. Infect. Immun.67:4725–4731.
16. Hassan, A. A., A. Abdulmawjood, A. O. Yildirim, K. Fink, C. Lammler, and R. Schlenstedt.2000. Identification of streptococci isolated from various sources by determination ofcfbgene and other CAMP-factor genes. Can. J. Microbiol.46:946–951.
17. Jansen, R., J. Briaire, E. M. Kamp, A.L. Gielkens, and M. A. Smits.1995. The CAMP effect ofActinobacillus pleuropneumoniae is caused by Apx toxins. FEMS Microbiol. Lett.126:139–143.
18. Jiang, M., L. A. Babiuk, and A. A. Potter.1996. Cloning, sequencing and expression of the CAMP factor gene ofStreptococcus uberis. Microb. Pathog.
20:297–307.
19. Jurgens, D., B. Sterzik, and F. J. Fehrenbach.1987. Unspecific binding of group B streptococcal cocytolysin (CAMP factor) to immunoglobulins and its possible role in pathogenicity. J. Exp. Med.165:720–732.
20. Ke, D., C. Menard, F. J. Picard, M. Boissinot, M. Ouellette, P. H. Roy, and M. G. Bergeron.2000. Development of conventional and real-time PCR assays for the rapid detection of group B streptococci. Clin. Chem.46:324– 331.
21. Kilian, M.1998. Streptococcus and Lactobacillus, p. 633–667.InA. Balows and B. I. Duerden (ed.), Topley and Wilson’s microbiology and microbial infections, 9th ed. Arnold, London, United Kingdom.
22. Orafu, C., P. Gill, C. Nelson, B. Hecht, and M. Hopkins.2002. Perianal versus anorectal specimens: is there a difference in group B streptococcal detection? Obstet. Gynecol.99:1036–1039.
23. Phillips, E. A., J. W. Tapsall, and D. D. Smith.1980. Rapid tube CAMP test for identification ofStreptococcus agalactiae(Lancefield group B). J. Clin. Microbiol.12:135–137.
24. Podbielski, A., O. Blankenstein, and R. Lutticken.1994. Molecular charac-terization of the cfb gene encoding group B streptococcal CAMP-factor. Med. Microbiol. Immunol.183:239–256.
25. Ratner, H. B., S. W. Lyndell, and C. W. Stratton.1986. Evaluation of spot CAMP test for identification of group B streptococci. J. Clin. Microbiol.
24:296–297.
26. Ripio, M. T., C. Geoffroy, G. Dominguez, J. E. Alouf, and J. A. Vazquez-Boland.1995. The sulphydryl-activated cytolysin and a sphingomyelinase C are the major membrane-damaging factors involved in cooperative (CAMP-like) haemolysis ofListeriaspp. Res. Microbiol.146:303–313.
27. Ruhlmann, J., B. Wittmann-Liebold, D. Jurgens, and F. J. Fehrenbach.
1988. Complete amino acid sequence of protein B. FEBS Lett.235:262–266. 28. Scharg, S. J., S. Zywicki, and M. M. Farley.2000. Group B streptococcal disease in the era of intrapartum antibiotic prophylaxis. N. Engl. J. Med.
342:15–20.
29. Schuchat, A.2000. Neonatal group B streptococcal disease—screening and prevention. N. Engl. J. Med.343:209–210.
30. Skalka, B., and J. Smola.1981. Lethal effect of CAMP-factor and uberis-factor: a new finding about diffusible exosubstances ofStreptococcus agalac-tiaeandStreptococcus uberis. Zentbl. Bakteriol. Parasitenkd. Infektkrankh. Hyg. Abt. 1 Orig. Reihe A249:190–194.
31. Sterzik, B., and F. J. Fehrenbach.1985. Reaction components influencing CAMP factor induced lysis. J. Gen. Microbiol.131:817–820.