Molecular Analysis
of the Maize wx-B3 Allele Indicates That Precise
Excision of the Transposable
Ac
Element Is Rare
George Baran,' Craig Echt,2 Thomas Bureau and Susan Wessler'
Botany Department, University of Georgia, Athens, Georgia 30602
Manuscript received June 24, 1991 Accepted for publication October 10, 199 1
ABSTRACT
The somatic and germinal behavior of the maize wx-B3 mutation indicates that this Ac allele rarely reverts. Endosperms containing wx-B3 display tiny and infrequent Wx revertant sectors while no significant reversion is detected when wx-B3 pollen is stained with I/KI. Previous studies of other transposable element alleles that revert infrequently have implicated low levels of element excision. Unlike these other alleles, the wx-B3 Ac element is indistinguishable from fully active Ac elements with
respect to its structure, and its ability to transpose from the Wx gene or to trans-activate a Ds element. Characterization of somatic and germinal excision events lead us to conclude that excision of the wx-
8 3 Ac element almost always produces null alleles. Furthermore, the excellent correlation between the position of the wx-B3 mutation on the physical and genetic maps indicates that the Ac insertion is the only lesion of wx-B3. As a result, precise excision of this Ac should restore Wx function. The fact that revertant sectors and pollen grains are rare indicates that precise excision of Ac is also rare. The finding that the wx-B3 reversion frequency is comparable whether wx-B3 is hemizygous or over a wx
allele with a wild-type insertion site illustrates a fundamental difference between the excision mecha-
nisms of Ac and Drosophila P elements.
T
RANSPOSABLE element alleles in maize have characteristic frequencies of somatic and ger- minal reversion. BRINK and WILLIAMS (1973) noted that 26 independent alleles of the R-nj gene, each containing the transposable element Activator (Ac), differed in their frequency and timing of revertant somatic sectors resulting from Ac excision. Based on additional genetic tests, they concluded that the posi- tion of Ac within the R-nj gene was primarily respon- sible for the resultant phenotypes. The discovery of transposon footprints has provided a simple explana- tion for these observations. Insertion of Ac andDs
elements creates an 8-bp duplication of host sequence that flank the element as a direct repeat. Upon exci- sion, the element is removed but part of the duplica- tion usually persists and results in additional nucleo- tides at the original insertion site (SUTTON et al. 1983). T h e effect of these footprints on gene expression
depends on whether the insertion site is exon, intron or untranscribed sequences.
Transposon footprints cannot explain all of the observed differences in the timing and frequency of somatic and germinal reversion. Some unstable ger- minal derivatives of transposable element alleles dis- play fewer and smaller sectors on average than their unstable progenitors. Some of these so-called change in state derivatives have sustained intraelement dele-
' Deceased.
' Present address: Department of Plant and Soil Sciences, Montana State
' To whom correspondence should be addressed. University, Bozeman, Montana 59717.
(;enetics 130: 377-384 (February, 1992)
tions that remove cis-requirements for transposition thus inhibiting element excision (SCHWARZ-SOMMER et al. 1985; SCHIEFELBEIN et al. 1988). In contrast,
somatic sectors may have nothing to do with element excision but rather correlate with the elements meth- ylation state. For example, green leaf sectors on pale green plants containing the hcf106 allele d o not reflect excision events of the Mutator element inserted at the
5' end of the hcf gene. T h e sectors d o however cor- relate with the methylation state of Mutator sequences that accompanies the restoration of hcffunction (MAR-
TIENSSEN et al. 1990).
T h e numerous Ac and
Ds
alleles of the maize w x378 G. Baran et al.
was able to map several wx alleles containing the non- autonomous
Ds
element, including wx-ml, wx-B# and wx-m6, becauseDs
alleles are stably mutant if the autonomous Ac element is not also in the genome. T h e cloning and characterization of the Wx gene (SHURE, WESSLER and FED~ROFF 1983) and many of its mutant alleles including wx-ml (WESSLER et al.1986), wx-B4 (WESSLER, BARAN and VARACONA 1987) and wx-m6 (FEDOROFF, WESSLER and SHURE 1983), revealed the excellent correlation between the genetic fine structure and physical maps of waxy (WESSLER and VARACONA 1985).
Surprisingly, one Ac allele, wx-B3, has been mapped genetically. This allele was isolated by BRINK and ASHMAN (see NELSON 1968) and classified as an Ac- induced mutation based on two criteria. First, wx-B3 endosperms display tiny (1 -1 0 cell) and infrequent revertant sectors on a null wx background. Second, wx-B3 can trans-activate
Ds
alleles. NELSON (1 968) was able to assay intragenic recombination between wx-B3 and other wx mutations because the pollen of wx-B3 containing plants, like the pollen produced by plants containing stable wx mutations, stains uniformly. red- dish brown with I/KI. Both the germinal and somatic behavior of wx-B3 suggests that this Ac allele rarely reverts.Other transposable element alleles with similar un- stable phenotypes have been characterized at the mo- lecular level in maize. For almost all of these alleles the low frequency of reversion is associated with low levels of element excision due to intraelement dele- tions (SCHWARZ-SOMMER et al. 1985; SCHIEFELBEIN et al. 1988). Unlike these alleles, the data presented in this study indicate that the wx-B3 Ac element is indis- tinguishable from fully active Ac elements with respect to its structure and its ability to transpose from the Wx gene or activate a
Ds
element. Rather, our data suggest that excision of the wx-B3 Ac element almost always inactivates the Wx gene and produces null alleles. We discuss the possibility that the W x sectors seen in wx-B3 endosperms result from precise loss of Ac from Wx. T h e low frequency of W x sectors and W x pollen grains conditioned by this allele suggests that precise loss of Ac is an extremely rare event.MATERIALS AND METHODS
Maize strains: Strains carrying wx-B3 and B3-SI alleles were obtained from OLIVER NELSON; wx-m9 was obtained from BARBARA MCCLINTOCK. T h e Maize Genetics Cooper- ation Stock Center provided the strains containing the wx
alleles C, 8 7 , C34 and the Ds allele bz2-m.
Isolation of wx-B3 and B 3 S l : T h e wx-B3 allele was recovered during a selection for M p ( A c ) induced mutations (by Brink and Ashman, see NELSON 1968). It was originally in a W23 background and NEL~ON subsequently crossed it into the M14 background for his recombination studies. A pedigree, kindly provided to one of us (C.E.) by NELSON, showed that the mutant identified as B 3 S l was established from a series of outcrosses of wx-B3 in the M 14 background
to another inbred, W22. Because only the wx character was selected for in these crosses, the loss of Ac activity (in 8 3 S l ) was not detected and when it occurred is not known.
Molecular analysis of wx-B3 and B 3 S l : Isolation of genomic DNA, Southern blot analysis and screening of genomic libraries were described previously (WESSLER et al. 1986). Genomic DNA from plantlets homozygous for either wx-B3 or BPS1 were digested with BglII, ligated into BamHI- digested EMBL4 vector, packaged in vitro and used to infect LE392 cells. Plaques that hybridized to labeled Wx probes
were subcloned as SalI fragments into p U C l l 9 (VIEIRA and
MESSING 1987) (see Figure 2 for the position of relevant restriction sites) and further subcloned as PstI, SphI o r BamHI-Sal1 fragments in preparation for DNA sequencing using the dideoxy method (SANGER, NICKLEN and COULSEN 1977). T h e site of A E insertion (in wx-B3) and the nature of the transposon footprint (in B3-SI) were determined by comparison to the Wx sequence (KLOSGEN et al. 1986). T h e sequence of the B3-Ac termini was compared to the Ac sequence (POHLMAN, FEDOROFF and MESSING 1984).
Isolation of B3-SI cDNA: Poly(Af) RNA was isolated from the endosperms of kernels harvested 18-22 days after pollination (DAP) as previously described (SHURE, WESSLER and FEDOROFF 1983). Double-stranded cDNA was prepared from 2 pg poly(A+) RNA (GUBLER and HOFFMAN 1983), ligated with EcoRI linkers, cloned into XgtlO, screened, subcloned and sequenced as described above for wx-B3.
Polymerase chain reaction (PCR) analysis of somatic excision: Three hundred nanograms of w~B3-containing genomic DNA were added to 100 PI of PCR cocktail con- sisting of 1 X PCR buffer (10 mM Tris-HCI, pH 8.3, 50 mM KC], 1.5 mM MgCI2, and 0.01% gelatin), 200 PM of each dNTP, 70 ng each of the primers 5' (5'-
AGGCGTTGCAGGCGGAGGCG-3') and 3' (5'- CGCTGAGACGGCCCATGTGG-3') [corresponding to
Wx position 2325-2344 and 2974-2955, respectively (KLOS- GEN et al. 1986)], and 2.5 units of Taq DNA polymerase (Perkin-Elmer-Cetus). Forty cycles of amplification were
carried out (95O, 1 min; 65", 2 min; 72", 3 min) in an Ericomp Thermal Cycler followed by a 1 0-min final exten- sion at 72". Fifty microliters of the reaction mixture were digested with 10 units each of the restriction endonucleases PstI and SalI (Figure 2) and the resulting fragments cloned into pUC 1 19 and sequenced by the dideoxy chain termina- tion method using a Sequenase kit (U. S. Biochemical).
Wx protein analysis: Wx protein was isolated from fro- zen 18-22 DAP endosperms and analyzed on sodium do- decyl sulfate (SDS)-polyacrylamide gels as described previ- ously (ECHT and SCHWARTZ 1981). Isolation of starch gran- ules from mature dried endosperm was modified by soaking the kernels in water for 24 hr then proceeding as for frozen immature endosperms. T h e gels in this study were 10% polyacrylamide and were stained with silver (OAKLEY,
KIRSCH and MORRIS 1980).
RESULTS
Ac Excision from wx-B3 379
TABLE 1
Number of wx-E3 kernels showing Wx seetors
No. Wx sectors/filed section of
endosperm" Endosperm
genotype 0 1 2 (or more)
B?/wx/wx 35 12 12
B?/B?/wx 41 16 2
83/83/83 42 12 5
a The wx allele used was B7, a deletion of most of the Wx gene
(WESSLER and VARACONA 1985). The values indicate the number o f sectors seen on an area equivalent to about 20% of the kernel surface, not on the entire endosperm. Most sectors were only one or a few cells large. Most kernels in the 2 (or more) class had just two sectors except for B?/wx/wx kernels which frequently had three sectors.
filed portions of the endosperm revealed few, small amylose containing sectors (Wx sectors). Such sectors were usually only composed of one or a few cells. The frequency of Wx sectors in endosperms containing wx- 8 3 is given in Table 1. Generally, as Ac dosage in- creases the timing of Ac and
Ds
transposition during development is delayed a n d the number of revertant sectors decreases. Similarly, increased dosage of wx- B 3 was correlated with fewer sectors (Table 1) and a slight decrease in the already small Wx sector size.The wx-B3 allele displays fewer revertant sectors than all other active Ac alleles examined. There are four possible reasons for this phenotype: (i) the Ac element may be active only late in the development
of the endosperm, (ii) the Ac element may be defective due to the production of a mutant transposase, (iii) the Ac element may be defective in its response to transposase, e.g., it may lack certain cis-requirements for tranposition or (iv) the Ac element is normal, however, excision from waxy usually results in null derivatives.
Truns-activation of
Ds:
T o assess the temporal expression and quality of transposase synthesized by the wx-B3 Ac element, we analyzed the effect of in- creasing wx-B3 dosage on aDs
element at the bz2 locus. The results, presented in Figure 1, indicate that the ability of wx-B3 to trans-activate aDs
element is indistinguisable from an active Ac element with re- spect to frequency, timing and dosage. More impor- tantly, the large Bz2 sectors seen in the presence of one and two doses of wx-B3 indicate that the wx-B3 Ac transposase activity is not restricted to the latter stagesof kernel development. These data indicate that rea- sons i and ii, above, are not responsible for the phe- notypic expression of wx-B3.
Cloning of the m-B3 gene, correlation between the genetic and physical maps: T o assess the integrity of the Ac element and its position of insertion, the element and flanking sequences were cloned from the wx-B3 gene. The insertion was first localized by South- ern blots to a 0.8-kb SalI fragment that includes intron 9 through exon 12 of the Wx gene (data not shown)
"
wxB3
"dosage
~-Ac
dosage
"_
-
0 1 0 1
. '
J:, ' '. 'I'
.
, .'i ' .
-., b
3
23
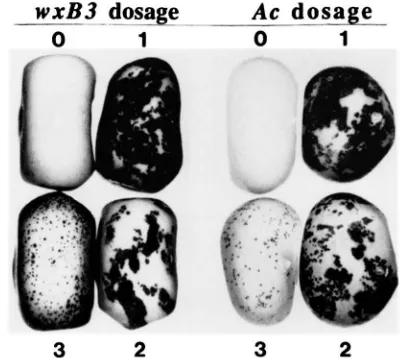
2FIGURE 1 .-Demonstration of trans-activating ability of wx-B?
(Ac) upon the Ds element in 6z2-m. The four kernels in each set are
representative of the four phenotypic classes of aleurone pigmen- tation patterns expected (and found) among sibling progeny from self pollinations of 622-m, wx-B?(Ac)/wx parents (left set) and 6zr?-m, wx/wx, Ac/- parents (right set). Excision of Ds from 622-m during kernel development results in fullcolored Br sectors in the lighter bronze-colored 622-m tissue. Increasing copies of Ac progressively delay the time of Ds excision during development.
(see Figure 2 for the position of this fragment). This Sal1 fragment was cloned and the sequences of the insertion site and the Ac termini determined (see
a 4.5-kb insertion at the exon 10-intron 10 junction. The 5' splice site of intron 10 is part of the 8-bp direct repeat that flanks the Ac element and was presumably generated upon Ac insertion (Figure
2).
As mentioned in the Introduction, NELSON (1968) was able to determine the relative position of the wx- B3 mutation on the fine structure map of the locus. With the cloning of wx-B3, correlation between the physical and genetic maps was possible. The lesions responsible for many of the wx mutations located near wx-B3 on the genetic map have been determined including wx-m8 (dSpm) (SCHWARZ-SOMMER et al. 1984), wx-ml
(Ds)
(WESSLER et al. 1986), wx-BI(Ds)
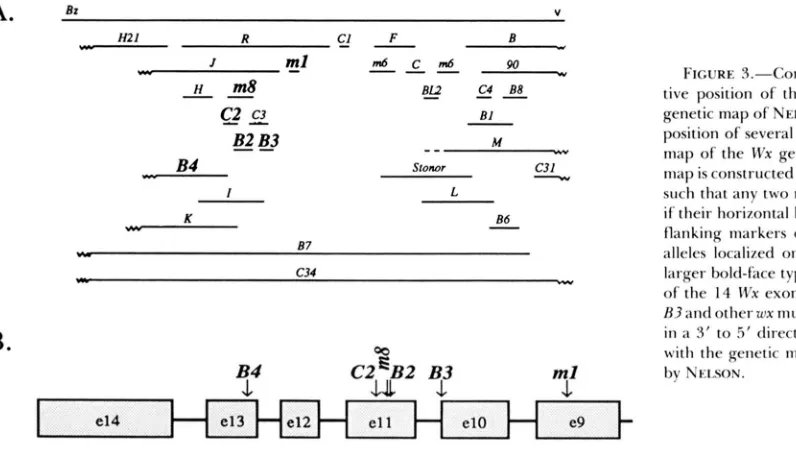
(WESSLER, BARAN and VARAGONA 1987), wx-B2 (a 133-bp insertion) (T.BUREAU and S . WESSLER, unpub- lished results) and wx-C2 (a 7-bp deletion) ( S . WHITE and S . Wessler, unpublished results). Comparison be- tween the genetic and physical maps (Figure 3, A and
B) reveals excellent correlation despite the short dis- tances involved (about 1 kb separates the wx-ml and wx-BI mutations whereas only about 120 bp are be- tween wx-B3 and wx-C2). Nelson's ability to accurately map the wx-B3 mutation in this crowded region of the fine structure map underscores the extremely low frequency of germinal reversion resulting from exci- sion of the wx-B3 Ac element.
380 G. Baran et al.
W X - B ~ - A C (4.5 kb)
e10 FIGURE 2.-The position m t l ori-
entation of the Ar insertion in the wx- R3 allele w i t h respect t o the exons (open boxes) and introns (connecting lines) of the IVx gene. The X-hp target site i n the Il'x gene is boxed and the
wx:
W X - B ~ :
J, 5' splice site
TCGT [TCTGCTG dtacgt
exon 10 " Intron 10
TCGT
lTCTCCTCd
t AC element +ITCTGCTGgltacgto r excision usually results in inactive Wx protein. To address this question directly we compared the rela- tive frequencies of somatic excision of wx-B3 Ac and another Ac element from the wx-m9 allele. T h e wx-m9
Ac element was cloned previously (FEDOROFF, WE.+ SLER and SHURE 1983) and is inserted only 30 bp upstream of the Ac insertion site in wx-B3. Both ele- ments are inserted into the same 0.8-kb SalI fragment (Figure 2) but in opposite orientation relative to each other. Genomic DNA from wx-m9 and wx-B3 contain- ing strains was isolated from the leaves of ten pooled plantlets for each genotype and analyzed on Southern blots (Figure 4). For each sample two fragments hy- bridize with the 0.8-kb SalI probe: the 5.4-kb frag- ment contains the entire Ac element whereas the 0.8 kb-Sal1 fragment results from Ac excision. These data establish that Ac excises from both the wx-B3 and wx-
m9 alleles at a high frequency. However, this fre-
quency is probably not as high as that suggested by the relative intensities of the 5.4-kb and 0.8-kb bands. T h e 0.8-kb wild-type probe may hybridize better with the 0.8-kb fragment than it does to the same fragment interrupted by a 4.6-kb imertion.
In addition, consistent with this demonstration of transposition competence is our finding that the se- quence of approximately 200 bp of each wx-B3 Ac
terminus is identical to the published Ac sequence (POHLMAN, FEDOROFF and MESSING 1984; MULLER- NEUMANN, YODER and STARLINCER 1984).
The W x protein of wx-B3: One measure of Ac
excision from the Wx gene during kernel development is the appearance of amylose sectors visualized by I/ KI staining. However, amylose sectors signify only those excisions that restore Wx enzymatic activity. I f the wx-B3 Ac element excises frequently in endosperm, as it does in leaf tissue (Figure 4), then the rarity of
B.
position of the normal 5 ' splice site is shown. This X-bp IVx sequence flanks the Ar insertion in wx-R3.
I
K w
00
44
C2,:,B2 B3FIGURE 3.-Correlation between the rela- tive position of the wx-R3 mutation on the genetic map of NEUON ( 1 968) and the actual position of several mutations on the physical map of the Wx gene. A, The fine structure map is constructed as a complementation map such that any two mutants do not recombine if their horizontal lines overlap. Rz and V are flanking markers on the chromosome. The alleles localized on the physical map are in larger bold-face type. B. A physical map of 9 of the 14 Wx exons with the position of wx- R 3 and other wx murations. The gene is drawn i n a 3' to 5 ' direction to fitcilitate alignment
w i t h the genetic map a s presented originally by NEISON.
1
2
3
..
9 - 5 . 4 k b-
II.-
-0.8FIGURE 4."Southern blot analysis of genomic DNA from the
rux-S3 and wx-m9 alleles. Genomic DNA from plantlets grown from
10 kernels each of the following kernel genotype: ( 1 ) wx-B3/wx-B3/
w x (2) wx-B3/wx/wx and (3) wx-m9/wx/wx was digested with Sal 1 and
probed with the wild-type 0.8-kb Sal1 probe (Figure 2). The 5.4- kb fragment represents the 4.6-kb Ac element in the 0.8-kb Sal1 fragment. The 0.8-kb fragment results from Ac excision during plant development. The wx allele was wx-C34, a deletion of the entire gene (WFSSLER and VARAGONA 1985).
Wx sectors in wx-B3 endosperms suggests that most excisions produce inactive Wx protein. T o address this question we used SDS-polyacrylamide gels to com- pare the relative amounts of starch granule-bound Wx protein in individual wx-B3 kernels with kernels con- taining many Wx sectors conditioned by the wx-m6
(Ds)
allele. Previous studies demonstrated that wx-B3 endosperms contain a protein that comigrates with wild-type Wx protein on SDS-polyacrylamide gels but displays altered mobility on isoelectric focusing gels (ECHT and SCHWARTZ 1981). In contrast, wx-m6(Ds)
endosperms encode a Wx protein with slower mobility on SDS-polyacrylamide gels (SHURE, WESSLER and FEDOROFF 1983). This slight difference in Wx protein mobility is evident from the profile of starch granule- bound proteins from a heterozygous wx-m6/wx-m6/ wx-B3 endosperm (Figure 5 , center, an asterisk indi- cates the position of Wx protein). Comparison of representative kernels containing either wx-B3 or wx-m6 (+1 Ac) reveals approximately equal amounts of Wx protein despite dramatically different patterns of Wx sectors (Figure 5 ) . If all the Wx protein detected in these endosperms were enzymatically active, ap- proximately the same degree of Wx sectoring would be expected on wx-m6 (+I Ac) and wx-B3 kernels. However, the fact that Wx sectors in wx-B3 endo- sperms are rare indicates that most of the Wx protein made by this allele is inactive. Although the Wx pro- tein associated with the rare revertant sectors in wx- B3 endosperms must be active, it probably represents an insignificant fraction of the total Wx protein.Isolation and cloning of the null wx-BPS1 allele:
- - .
m6
x
W A ) r l CIFIGURE 5.--Correlation of the degree of W x revertant sectoring and the amount of Wx protein among triploid endosperms contain- ing: one dose of wx-B3, one dose of wx-B3 with two doses of wx-
mh(Ds), and one dose of an unlinked Ac with two doses of wx-mh.
The mature dried kernels were cut in half, stained with I/KI
solution and a camera lucida tracing made before their endosperm starch granule-bound proteins were exanlined by SDS-polyacryl- amide gel electrophoresis. The asterisk indicates the position of the Wx protein.
T h e above data suggest that excision of the wx-B3 Ac
element almost always results in null derivatives in both somatic and germinal lineages. T h e preponder- ance of null derivatives could reflect the consequence of insertion into a splice donor site, where any tran- sposon footprint might interfere with normal splicing. Alternatively, the integrity of the insertion site might be critical for Wx enzymatic activity and any footprint might be incompatible with gene function.
To address this question we have characterized a null derivative of wx-B3 called B3-SI. The origin of this allele is described in MATERIALS AND METHODS. T h e B3-SI allele is null for Wx expression despite the fact that it encodes wild-type levels of a protein that comigrates on isoelectric focusing gels with the slightly altered Wx protein found in wx-B3 endosperms (data not shown). To understand how this protein is altered, we cloned the B3-SI gene and determined that there is a 6-bp footprint at the site of Ac excision (Figure 6). If translated and correctly spliced, the additional 6 bp would add two leucine residues to the Wx protein. However, it is also possible that the additional 6 bp near the 5' splice site affected splice site selection. T o
wx: TCGlTCTGCTG gtacgtgtg ... ... atgctgcag GGCACGGGCAA
wx-B3-S1: TCGTTCTGCTGCTGCTG gtacgtgtg ... ... atgctgcag GGCACGGGCAA
FIGURE 6.-The DNA sequence of the B?-SI derivative of wx-
382 G . Baran et al.
wx -TCTGCTGG-
w - B 3 -TCTGCTGG AC TCTGCTGG-
WX-B~SZ
Ex W1 -TCTGCTG.. CTGCTGG-
-TCTGCTG
. .
CTGCTGG-Ex #2
Ex W3
Ex #5 -TCTGCTGECTGCrGG-
- W G C K . ACTGCTGG-
-TCTGCTGC. CKCTGO-
EXM -TCKKTGGACTGCTGG-
FIGURE 7,"Transposon footprints resulting from excision of the wx-B3Ac element. B3-SI is the germinal derivative whereas Ex #1-
#5 are somatic excisions. The underlined bases indicate transver- sions and transitions of central nucleotides.
determine whether sp1icir.g was altered in BJ-SI, we analyzed Wx mRNA on Northern blots and found that the allele encoded wild-type levels of a wild-type sized W X mRNA (2.4 kb, data not shown). In addition, DNA sequence analysis of B3-SI cDNA (see MATE- RIALS AND METHODS) indicated that the mRNA was correctly spliced and differed from the non-mutant mRNA simply by the addition of two Leu codons (Figure 6 ) .
Somatic excision of Ac from the wx-B3 gene: T h e B3-SI allele defines one Ac excision event; attempts to isolate similar germinal events were not successful. We were unable to identify any revertants among
2
1,000 wx-B3/wx/wx kernels screened. In addition, null derivatives like B3-SI are difficult to isolate be- cause they are virtually indistinguishable from the wx- B 3 progenitor with respect to the mutant wx pheno- type. As an alternative approach to defining wx-B3 Ac excision events, we analyzed the footprints that re- sulted from somatic excision in leaf tissue. To this end, the site of wx-B3 Ac excision was amplified by PCR from the genomic DNA used previously for Southern blot analysis (Figure 4) (SAIKI et al. 1988). Since the genomic DNA was likely to contain many independent excision events, the products of two am- plification reactions were cloned and sequenced indi- vidually. T h e sequences of 30 independent clones contained one of the five transposon footprints shown in Figure 7. Among this collection, the only footprint that restored the correct reading frame was the same as the 6-bp B3-SI footprint.DISCUSSION
The wx-B3 mutation displays an extremely low fre- quency of reversion during both endosperm and spo- rophytic development. Previous studies of other trans- posable element alleles that revert infrequently have implicated low levels of element excision (SCHWARZ- SOMMER et al. 1985; SCHIEFELBEIN et al. 1988). Unlike these other alleles, the Ac element of wx-B3 is fully active based on three criteria: (i) the ability to trans-
activate a Ds element (Figure l), (ii) the frequency of excision from the Wx gene (Figure 4) and (iii) the integrity of the terminal sequences determined to be important cis-elements necessary for transposition (COUPLAND et al. 1989).
Despite the restriction of Wx sectors to late kernel
development, the presence of large Bz2 sectors on bz2-
m (+wx-B3) containing kernels (Figure 1) indicates that the wx-B3 Ac transposase is present early in kernel development. The relative abundance of Wx protein in wx-B3 endosperms (Figure 5 ) is also consistent with this conclusion. Similarly, the abundance of the 0.8- kb Sal1 fragment in wx-B3 leaf tissue (Figure 4) indi- cates that the wx-B3 Ac element can excise throughout sporophytic development despite the absence of black- staining pollen grains among wx-B3 pollen.
Recent reports that Ac/Ds elements can function as introns (reviewed in WEIL and WESSLER 1990) sug- gests an alternative explanation for the origin of the altered Wx protein detected in wx-B3 endosperms. Several reasons have led us to conclude that this Wx protein arises from a wx-B3 gene that contains a tran- sposon footprint rather than from Ac containing pre- mRNA that is subsequently spliced. First, the wx-B3 element is not in the same orientation, with respect to the Wx gene, as other Ac/Ds elements that behave as introns (WESSLER, BARAN and VARAGONA 1987; DEN-
NIS et al. 1988). Second, the amount of wx-B3 encoded
protein decreases with increasing doses of the wx-B3 gene in the endosperm (ECHT 1982). T h e splicing model predicts a linear relationship between the num- ber of wx-B3 doses and the amount of Wx protein. In contrast, the result is consistent with the excision- footprint model since the frequency of Ac transposi- tion decreases with increasing Ac dosage. Finally, the altered Wx proteins detected in wx-B3 and B3-Sl endosperms comigrate on isoelectric focusing gels (ECHT 1982) suggesting that they have sustained the same lesion (two additional codons encoded by the transposon footprint, Figure
7).
Taken together our data indicate that low levels of wx-B3 Ac activity and/or transposition cannot account for the low frequency of reversion. Rather, we believe that two factors are responsible: (1) most and possibly all transposon footprints are incompatible with Wx enzymatic activity and (2) precise excision of Ac is extremely rare.
T h e B3-Sl derivative allele contains a 6-bp transpo- son footprint and encodes wild-type levels of a slightly altered but inactive Wx protein (ECHT and SCHWARTZ
1981). As mentioned above, this protein comigrates with the Wx protein isolated from wx-83 endosperms (ECHT 1982) suggesting that this is the predominant Wx protein resulting from wx-B3-Ac excision. In sup- port of this idea, analysis of somatic excision events from leaf tissue (Figure 4) identify this same 6 bp footprint as the only excision product that maintains the correct reading frame (Figure 7).
Ac Excision from 383
pendent lesions and that the B 3 S l Wx protein is null, not because it contains two additional Leu codons, b u t due to a second mutation elsewhere in the B3-S1 gene. Although a two lesion scenario might also ex- plain the rarity of Wx sectors and revertants condi- tioned by wx-B3 (both mutations would have to revert to restore Wx activity), this explanation is not consist- ent with the excellent correlation between the position of wx-B3 on the genetic and physical maps. T h e wx- B3 mutation was localized to a very restricted region of the Wx gene. If two independent mutations are responsible for the wx-B3 phenotype, they must be very tightly linked. Our data rule out the possibility of a closely linked second lesion; DNA sequence analy- sis of 800 bp flanking the Ac insertion site failed to detect an additional mutation that could potentially alter the Wx protein. If the Ac insertion is the only lesion of wx-B3 then precise loss of Ac should restore Wx function. T h e fact that Wx sectors and black staining pollen grains are rare indicates that precise excision of Ac rarely occurs. Based on this interpreta- tion, the apparent restriction of Wx sectors to late kernel development suggests a probabalistic con- straint rather than a temporal one. T h e tremendous increase in the number of endosperm cells during kernel development provides increasing opportunities to reveal low probability events. Thus, given a large enough number of wx-B3 kernels, there should be a few kernels that display large sectors representing perfect excision occurring earlier in kernel develop- ment. Examination of over 2 1,000 kernels resulting from a cross of homozygous wx-B3 plants onto wx testers identified 10 kernels with larger Wx sectors, however none contained a Wx sector that accounted for more than 10% of the kernel surface.
T h e wx-B3 allele thus provides a measure of the extremely low frequency of perfect excision of the Ac
element. In fact, since reversion may result from 3-bp footprints, rare wx-B3 sectors and black staining pollen grains may overestimate precise excision. It is impor- tant to know whether this behavior is peculiar to this allele or whether perfect excision is also unusual when other Ac and Ds alleles are examined. Virtually all previously reported Ac and Ds footprints are between 5 and 9 bp with most between 6 and 8 bp. These include footprints of the Ac allele wx-m9 (POHLMAN, FEDOROFF and MESSING 1984) and Ds footprints of the Bz-wm (SULLIVAN, SCHIEFELBEIN and NELSON 1989), Adhl-2FZl (CHEN, FREELING and MERCKEL-
RACH 1986), Adhl-Fm335 (SUTTON et al. 1983; PEA-
COCK et al. 1984), sh6233 (WECK et al. 1984), wx-ml
(WESSLER et al. 1986), wx-B4 (M. VARACONA and S. WESSLER, unpublished results) and Wx-m5 (WEIL et al. 1992) alleles. Although most of these footprints were isolated from revertants, there is not a single example of a 3-bp footprint. Similarly, no 1-, 2- or 4-bp foot- prints have been reported among somatic excisions or
revertants of Ds insertions in untranslated regions. Interestingly, there are two reported instances of perfect excision; in both cases the possibility of wild- type contamination was ruled out. CHEN, FREELING and MERCKELBACH (1986) identified a perfect excision of the Ds2 element from the Adhl-2Fll allele. This excision event was only one of 18 Adh+ alleles found among 94,000 kernels analyzed. T h e low frequency supports the notion that precise excision of Ac and Ds elements is a rare event. In contrast, DENNIS, GER- LACH and PEACOCK ( 1 986) identified a precise excision event of Dsl among 8 Adh+ derivatives of Adhl- Fm335. In this study the frequency of reversion was almost 3%. The isolation of a single precise excision event does not contradict the notion that these are rare events. However, if there is a difference in the frequency of precise excision between Dsl and Ac-like
Ds
elements, this difference may be a consequence of their dramatic structural differences and their distinct cis-requirements for transposition (COUPLAND et al.1989).
It is surprising that there are two reports of precise excision yet not a single report of 1-,
2-,
3- or 4-bp footprints. T w o models have been proposed to explain the origin of footprints. According to PEACOCK et al. (1984) and SAEDLER and NEVERS (1 985), Ac and Dselements are excised by staggered cuts in the 8-bp target site duplication. T h e single-stranded ends are repaired prior to ligation. Differences in the length and sequence of footprints are hypothesized to result from exonuclease activity associated with DNA repair. Although precise excision might reflect more exten- sive exonuclease activity resulting in the loss of one copy of the duplication, less severe exonuclease activ- ity should produce small footprints of 1 to 4 bp. Interestingly, footprints of this size have not been reported to date. To explain why precise loss is seen but not small footprints, DENNIS, GERLACH and PEA-
COCK (1 986) suggested that a second mechanism, pos-
sibly transposase mediated reciprocal recombination between the duplicated flanking sequences, may be involved in generating precise excision events. We agree with their idea concerning two mechanisms and further suggest that the wx-B3 allele provides a mea- sure of the frequency of the alternative mechanism leading to precise excision.
Precise excision is frequently associated with exci- sion of the Drosophila P element. Recently ENCELS et al. (1 990) demonstrated that precise loss o f P elements only occurred when the homolog had a wild-type copy of the donor sequence. They proposed that precise
384 G . Baran et al.
mechanism cannot explain precise loss of the Ac ele- ment. For P elements, the frequency of precise exci- sion increases 100-fold when the homolog contains a wild-type sequence at the insertion site. In contrast, the frequency of wx-B3 reversion is indistinguishable in somatic and germinal tissues whether the allele is hemizygous or over a wx allele with a wild-type inser- tion site (NELSON 1968; S. R. WESSLER, unpublished results). As a consequence of these different excision mechanisms, it is unlikely that the Ac element can be used to facilitate gene replacement.
We thank STEVE DELLAPORTA and RITA VARAGONA for technical assistance and CLIFF WEIL and MICHAEL PURUGGANAN for critical reading of the manuscript. This work was supported by National Institutes of Health (NIH) grant GM32528 to S.R.W. and an NIH postdoctoral fellowship to G.B.
L I T E R A T U R E C I T E D
BRINK, R. A., and E. WILLIAMS, 1973 Mutable R-nauajo alleles of cyclic origin in maize. Genetics 73: 273-296.
CHEN, C.-H., M. FREELING and A. MERCKELBACH,
1986 Enzymatic and morphological consequences of Ds ex- cision from maize A d h l . Maydica 31: 93-108.
COUPLAND, G., C. PLUM, S. CHATTERJEE, A. POST and P. STARLIN-
GER, 1989 Sequences near the termini are required for trans-
position of the maize transposon Ac in transgenic tobacco plants. Proc. Natl. Acad. Sci. USA 8 6 9385-9388.
DENNIS, E. S., W. L. GERLACH and W. J. PEACOCK, 1986 Excision
of the Ds controlling element from the Adhl gene of maize. Maydica 31: 47-57.
DENNIS, E. S., M. M. SACHS, W. GERLACH, I. BEACH and W. J. PEACOCK, 1988 T h e D s l transposable element acts as an intron in the mutant allele Adhl-Fm335 and is spliced from the message. Nucleic Acids Res. 16: 3315-3328.
ECHT, C., 1982 An analysis of the wx gene of maize and its alleles by examination of the protein product of the waxy locus. Ph.D. thesis, Indiana University.
ECHT, C., and D. SCHWARTZ, 1981 Evidence for the inclusion of controlling elements within the structural gene at the waxy locus in maize. Genetics 9 9 275-284.
ENGELS, W. R., D. M. JOHNSON-SCHLITZ, W. E. EGGLESTON and J. SVED, 1990 High-frequency P element loss in Drosophila is homolog dependent. Cell 6 2 515-525.
FEDOROFF, N., S. WESSLER and M. SHURE, 1983 Isolation of the transposable maize controlling elements Ac and Ds. Cell 35:
GLOOR, G. B., N. A. NASSIF, D. M. JOHNSON-SCHLITZ, C. R. PRES-
TON and W. R. ENGELS, 1991 Targeted gene replacement in Drosophila via P element-induced gap repair. Science 253:
1110-1117.
GUBLER, U., and B. J. HOFFMAN, 1983 A simple and very efficient method for generating cDNA libraries. Gene 25: 263-269.
KLOSGEN, R. B., A. GIERL, ZS. SCHWARZ-SOMMER and H. SAEDLER,
1986 Molecular analysis of the waxy locus of Zea mays. Mol. Gen. Genet. 203: 237-244.
MARTIENSSEN, R., A. BARKAN, W. C. TAYLOR and M. FREELING,
1990 Somatically heritable switches in the DNA modification of Mu-transposable elements monitored with a suppressible mutant in maize. Genes Dev. 4 331-343.
MULLER-NEUMANN, M., J. I. YODER and P. STARLINGER,
1984 T h e DNA sequence of the transposable element Ac of Zea mays L. Mol. Gen. Genet. 198: 19-24.
235-242.
NELSON, 0. E., 1968 T h e waxy locus in maize. 11. Location of the NELSON, 0. E., and H. W. RINES, 1962 T h e enzymatic deficiencv
controlling element alleles. Genetics 99: 275-284.
in the waxy mutant of maize. Biochem. Biophys. Res. Commun. 9: 297-300.
OAKLEY, B. R., D. R. KIRSCH and N. R. MORRIS, 1980 A simple ultrasensitive silver stain for detecting proteins in polyacryl- amide gels. Anal. Biochemistry 105: 361-363.
PEACOCK, W. J., E. S. DENNIS, W. L. GERLACH, M. M. SACHS and D. SCHWARTZ, 1984 Insertion and excision of Ds controlling elements in maize. Cold Spring Harbor Symp. Quant. Biol. 49:
347-354.
POHLMAN, R., N. V. FEDOROFF and J. MESSING, 1984 T h e nucleo- tide sequence of the maize controlling element Activator. Cell
SAEDLER, H., and P. NEVERS, I985 Transposition in plants: a molecular model. EMBO J. 4: 585-590.
SAIKI, R. K., D. H. GELFAND, S. STOFFEL, S. J. SCHARF, R. HIGUCHI, G. T. HORN and K. B. MULLIS, 1988 Primerdirected enzy- matic amplification of DNA with a thermostable DNA polym- erase. Science 239: 487-49 1.
SANGER, F., S. NICKLEN and A. R. COULSEN, 1977 DNA sequenc- ing with chain-terminating inhibitors. Proc. Natl. Acad. Sci.
SCHIEFELBEIN, J. W., RABOY, V., KIM, H.-Y. and NELSON, 0. E. ,
1988 Molecular characterization of Suppressor-mutator (Spm)- induced mutations at the bronze-1 locus in maize: the bz-m13 alleles, pp. 261-278 in Plant Transposable Elements, edited by 0. NELSON. Plenum Press, New York.
SCHWARZ-SOMMER, ZS., A. GIERL, R. B. KLOSGEN, U. WIENAND, P. PETERSON and S. SAEDLER, 1984 T h e Spm ( E n ) transposable element controls the excision of a 2 kb DNA insert at the wx-
m8 locus of Zea mays. EMBO J. 3: 1021-1028.
SCHWARZ-SOMMER, ZS., A. GIERL, R. BERNDTGEN and H. SAEDLER,
model of Spm action. EMBO J. 4: 2439-2443.
1985 Sequence comparison of states of a l - m l suggests a
SHURE, M., S. WESSLER and N. FEDOROFF, 1983 Molecular iden- tification and isolation of the waxy locus in maize. Cell 35: 225- 233.
SULLIVAN, T. D., J. W. SCHIEFELBEIN and 0. E. NELSON,
1989 Tissue-specific effects of maize Bronze gene promoter mutations induced by D s l insertion and excision. Dev. Genet. 10: 412-424.
SUTTON, W. D., W. L. GERLACH, D. SCHWARTZ and W. J. PEACOCK,
1983 Molecular analysis of Ds controlling element mutations at the Adhl locus of maize. Science 223: 1265-1268.
VIEIRA, J., and J. MESSING, 1987 Production of single-stranded plasmid DNA. Methods Enzymol. 153: 3-1 1 .
WECK, E., U. COURAGE, H.-P. DORING, N. FEDOROFF and P. STAR- LINGER, 1984 Analysis of sh-m6233, a mutation induced by the transposable element Ds in the sucrose synthase gene of
Zea mays. EMBO J. 3: 1713-1716.
WEIL, C . F., and S. R. WESSLER, 1990 T h e effect of plant trans- posable elements on transcription inititation and RNA process- ing. Annu. Rev. Plant Physiol. Plant Mol. Biol. 41: 527-552.
WEIL, C., S. MARILLONNET, B. BURR and S. R. WESSLER,
1992 Changes in state of the maize Wx-m5 allele are due to
intragenic transposition of Ds. Genetics (in press).
WESSLER, S. R., G. BARAN and M. VARAGONA, 1987 T h e maize transposable element Ds is spliced from RNA. Science 237:
WESSLER, S. R., and M. VARAGONA, 1985 Molecular basis of mutations at the waxy locus of maize: Correlation with the fine structure genetic map. Proc. Natl. Acad. Sci. USA 82: 4177- 4181.
WESSLER, S. R., G. BARAN, M. VARAGONA and S. L. DELLAPORTA,
1986 Excision of Ds produces waxy proteins with a range of
enzymatic activities. EMBO J. 5: 2427-2432. 37: 635-643.
USA 74: 5463-5467.
916-918.