Improving Fat Utilization by the Weaned Pig: Effect of Diet Physical Form, Fatty Acid Chain Length, and Emulsification.
73
0
0
Full text
(2)
(3)
(4)
(5)
(8)
(9)
(10)
(11)
(12)
(13)
(14)
(15)
(16)
(17)
(18)
(19)
(20)
(21)
(23)
(24)
(25)
(26)(27)
(28)
(29)
(30)
(31)
(32)
(33)
(34)
(35)
(36)
(37)
(38)
(39)
(40)
(41)
(42)
(43)
(44)
(45)
(46)
(47)
(48)
(49)
(50)
(51)
Figure
+7
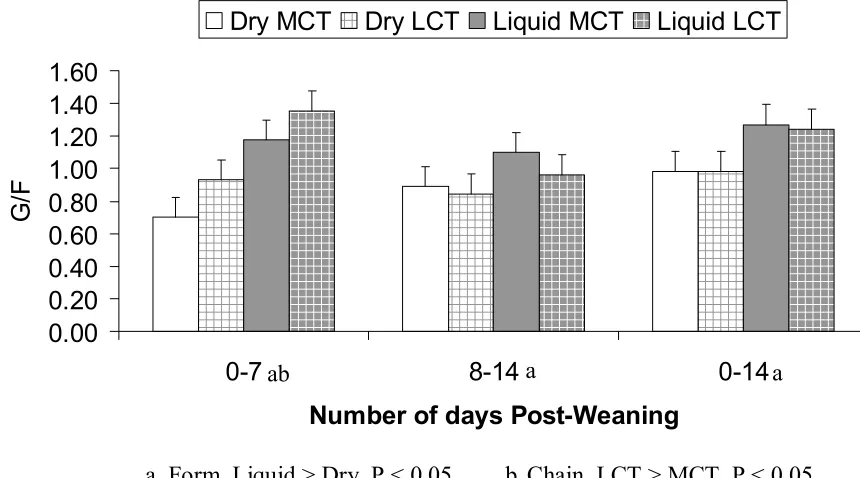

Related documents
Table 1.. Figure 3 and Figure 4 demonstrate the effects of suction parameter on temperature and concentration. It is clear that the fluid velocity decreases significantly
Phylogenetic analysis failed to yield species-specific clusters among the L1 elements isolated, and all Oryzomys sequences had numerous private mutations.. Finally, in
In this paper a new approach for block feature based image fusion is implemented using multi wavelet transform and neural networks and a qualitative analysis has been done for
The result of antibiogram of the isolates is shown in the Table-2 .Of the 242 enterobacteriaceae isolated 71 (29.3%) were ESBL producers, 48 of them belonged to Klebsiella
In summary, the IRO is a simple, engaging, and useful assessment tool that occupational therapists, speech-language pathologists, classroom teachers, and reading specialists can
(7), relating the moistu- re content with the dielectric constant and bulk density for TDR moisture measurements in peat deposits from the Biebrza river valley, is
According to the statement of the American Diabetes Association and European Association for the Study of Diabetes guidelines also endorses metformin is the first-line drug for
From the experimental results it can be concluded that the proposed genetic algorithm based on Hidden Markov Models and Fuzzy Levenshtein Distance works well under
