Use of Discriminatory Probes for Strain Typing of Formalin-Fixed,
Rabies Virus-Infected Tissues by In Situ Hybridization
Susan A. Nadin-Davis,* Mary Sheen, and Alexander I. Wandeler
Rabies Centre of Expertise, Ottawa Laboratory—Fallowfield, Canadian Food Inspection Agency,Nepean, Ontario K2H 8P9, Canada
Received 9 December 2002/Returned for modification 20 February 2003/Accepted 18 June 2003
An in situ hybridization (ISH) method has been developed to overcome difficulties encountered in the viral typing of formalin-fixed rabies virus-infected brain tissue. Rabies viruses representative of all strains normally encountered in diagnostic submissions throughout Canada, including 3 strains of terrestrial hosts (arctic fox, western skunk, mid-Atlantic raccoon), 10 strains circulating in several bat reservoirs (BBCAN1 to BBCAN7, LACAN, SHCAN, and MYCAN), and the Evelyn-Rokitniki-Abelseth (ERA) strain, used as an oral vaccine for fox rabies control in Ontario, were targeted. Partial phosphoprotein gene fragments generated from reverse transcription (RT)-PCR products of specimens of each viral type were molecularly cloned and used to produce negative-sense digoxigenin-labeled RNA transcripts. Conditions permitting the use of these transcripts as strain-specific probes were optimized by blotting analyses with RT-PCR amplicons generated with represen-tative rabies viruses and by ISH applied to mouse brains inoculated with these strains. The successful application of this methodology to two rabies virus-positive specimens that were also identified by traditional methods and the retrospective typing of two archival rabies virus-positive equine specimens is described. This technique provides a typing regimen for rabies virus isolates submitted in a form that is normally recalcitrant to alternate typing strategies.
Classical rabies viruses comprise genotype 1 of the Lyssavi-rusgenus and are responsible for encephalitic disease in a wide range of canid and chiropteran species (33). Comparative an-tigenic and genetic studies discriminate several distinct strains which circulate and persist in specific hosts but which can spill over into a broad range of mammalian species (32, 33). Where the geographical ranges of multiple rabies virus strains overlap, there is an ongoing need for accurate, rapid, and cost-effective methods of strain discrimination. The identification of the strain responsible for rabies virus infections provides epidemi-ological data of considerable importance to ongoing rabies control programs. This is particularly important throughout the Americas, where multiple rabies virus strains persist in several wildlife reservoirs (21, 29). Current control activities target specific terrestrial strains of the disease through oral baiting programs, for example, to control fox rabies in Ontario, Canada (18). Ongoing monitoring of cases of rabies, especially in areas considered free of terrestrial forms of the disease, is necessary to preclude the possibility that an incursion of such a strain into the region has occurred; such an event would be likely to precipitate control measures to prevent its spread. However, cases in which a terrestrial species is infected with a bat strain do not generally require a similar response, since these spillover events do not usually result in disease out-breaks.
Since the diagnosis of rabies is normally performed by a fluorescent-antibody test with fresh brain tissue (5), methods of viral typing with such specimens have been developed and
widely applied. Routine identification of rabies virus strains is usually achieved by using panels of monoclonal antibodies with discriminatory reactivities (24, 27). In recent years genetic tests, in which a portion of the viral genome is amplified by reverse transcription (RT)-PCR followed by sequence charac-terization of the resulting product (32), have been applied extensively.
Significant difficulties arise, however, when a specimen is available only as fixed tissue. Such a situation can arise in geographical areas of low rabies incidence (e.g., in the Mari-time provinces of Canada), in which a suspicion of rabies is considered only postmortem, or in species which are very in-frequently infected with this virus (e.g., in humans and horses in North America). Although a rabies diagnosis can be con-firmed with formalin-fixed tissues by immunohistochemistry (IHC), antigenic typing methods cannot usually be performed with such specimens since many of the monoclonal antibodies included in the discriminatory panels identify conformational epitopes which become undetectable upon tissue fixation, ow-ing to either the maskow-ing or the loss of these epitopes after such treatments.
The recovery of RNA from formalin-fixed tissues followed by amplification of viral sequences by RT-PCR has been suc-cessfully applied to the diagnosis of certain other viral infec-tions, e.g., those caused by bovine viral diarrhea virus (9), Sin Nombre virus (26), and hepatitis C virus (6). Moreover, the sequence characterization of amplicons from variable regions of the hepatitis C virus genome has allowed identification of the genotypes of the isolates responsible for the infections as well as molecular evolutionary studies of archival materials (30). However, successful application of this RT-PCR meth-odology depends critically on the recovery from formalin-fixed embedded tissues of RNA of sufficiently high quality to sup-* Corresponding author. Mailing address: Rabies Centre of
Exper-tise, Ottawa Laboratory—Fallowfield, Canadian Food Inspection Agency, 3851 Fallowfield Rd., Nepean, Ontario K2H 8P9, Canada. Phone: (613) 228-6698. Fax: (613) 228-6669. E-mail: nadindaviss @inspection.gc.ca.
4343
on May 15, 2020 by guest
http://jcm.asm.org/
port the RT and amplification reactions. The success of this extraction is variously reported to be adversely affected by prolonged formalin fixation and storage (10, 11). In practical terms these considerations may severely limit this method’s applicability in situations in which the testing laboratory has no control over specimen preparation prior to submission. In-deed, our own efforts to recover RNA suitable for amplifica-tion by RT-PCR from a collecamplifica-tion of archived rabies virus-positive formalin-fixed brain tissue blocks has met with only limited success, despite the targeting of relatively short (⬃ 150-bp) rabies virus sequences.
This report describes an alternative rabies virus typing strat-egy that applies the in situ hybridization (ISH) methodology to fixed brain tissues. Although ISH has been successful in de-tecting rabies virus sequences in research applications (14) and has been shown to be applicable to diagnosis (36), it has not previously been applied to rabies virus typing. Indeed, a major challenge to this approach has been the identification of a sufficiently divergent region within the rabies virus sequence that can provide a suitable discriminatory target. The unseg-mented negative-sense RNA genome of rabies virus contains five genes in the order 3⬘-N-P-M-G-L-5⬘ (32). A variety of rabies virus strains, as well as other members of theLyssavirus genus, have been genetically characterized at several of these loci, especially at the nucleoprotein (N) gene (4, 15, 28) and the glycoprotein (G) gene (1, 2). A sequence comparison along the lengths of the genomes of two lyssaviruses, the laboratory PV strain of rabies virus (genotype 1) and a Mokola virus isolate (genotype 3), which are representative of the extent of divergence within this genus, indicated extensive regions with moderate to high degrees of conservation (17). Two particular regions which exhibited relatively high degrees of divergence were (i) the transmembrane- and cytoplasm-encoding domains at the 3⬘ terminus of the G gene and the neighboring G-L intergenic region adjacent to the polymerase (L) gene and (ii) the central portion of the phosphoprotein (P) gene. The utility of the P gene and its encoded product as a useful discrimina-tory target has been further suggested by the generation of exquisitely strain-specific monoclonal antibodies targeting the central domain of the P protein (20). Furthermore, a recent study on the genetic diversity of the lyssavirus P-gene locus substantiated the divergent nature of its central section (22).
To assess the feasibility of using rabies virus P-gene se-quences for strain discrimination, we sought to develop a panel of probes targeting all of the indigenous rabies virus strains circulating within Canada. This includes strains of terrestrial hosts, as follows: the Western skunk strain (Mephitis mephitis host), with a range from Manitoba to Alberta; the Arctic fox strain (arctic fox [Alopex lagopus] and red fox [Vulpes vulpes] hosts), which persists to a limited degree in Ontario but which exists extensively in northern regions of the country; and the raccoon strain (Procyon lotorhost), a recent incursion from the United States which entered Ontario in 1999 (35) and New Brunswick in 2000. In addition, several rabies virus strains, characterized in detail by Nadin-Davis et al. (21), circulate in particular species of insectivorous bats, including big brown bats (Eptesicus fuscus); various Myotis species, including the little brown bat (Myotis lucifugus); members of the Lasiurus genus, including red and hoary bats; and silver-haired bats
(Lasionycteris noctivagans). While single antigenic viral types
are associated with most of these chiropteran species, those of big brown bats could be divided into seven groups (BBCAN1 to BBCAN7) (21). The ranges of most of these bats are exten-sive and in many areas overlap those of the terrestrial rabies virus reservoir species.
This study describes the development of several rabies virus strain-specific probes, demonstrates their discriminatory capa-bilities by using mouse brains experimentally infected with isolates representative of these Canadian rabies virus strains as well as material from two diagnostic submissions, and finally, confirms their utility by retrospective typing of the viruses responsible for rabies in two archival equine specimens.
MATERIALS AND METHODS
Rabies virus isolates.A description of the rabies virus isolates used for mo-lecular cloning is given in Table 1. Those isolates used for P-gene characteriza-tion, as illustrated in Fig. 1, included three rabies virus isolates of U.S. origin: two raccoon strain isolates from Florida (isolate V125) and New York (isolate NY516) and a red bat isolate (isolate V231) from Texas. All other specimens were received for rabies diagnosis, performed by the direct fluorescent-antibody test, at the laboratory in Nepean, Ontario (N), or Lethbridge, Alberta (L). The year of submission is indicated by the first two digits of the isolate submission number. When possible, original brain material was used directly as the viral source, but in some cases virus was passaged once by intracerebral inoculation into suckling mice of a 10% suspension of the original brain prepared in phos-phate-buffered saline (PBS) supplemented with antibiotics.
RT-PCR.Total RNA was prepared from original brain tissue infected with the selected rabies virus strains by using the TRIzol reagent according to the rec-ommendations of the manufacturer (Gibco BRL). P-gene amplification was undertaken by RT-PCR performed with 2g of RNA, as described elsewhere (22). PCR products were recovered by using a Wizard PCR Purification kit (Promega Corporation, Madison, Wis.) prior to cloning.
Phylogenetic analysis.The nucleotide sequences of the P genes of appropriate strains, as determined previously (22), were aligned by using the CLUSTALX program (31) and were then subjected to neighbor joining analysis with PHYLIP (version 3.5c) software (8), as described by Nadin-Davis et al. (21). A graphical output of the resulting tree was obtained by using the TREEVIEW program (23).
[image:2.603.301.542.79.255.2]Generation of molecular clones of rabies virus gene fragments.Restriction endonuclease maps were generated from the nucleotide sequence of each am-plicon, determined as described previously (22), by using DNASIS software; and appropriate restriction sites flanking the region of interest were identified. PCR products were cleaved by the selected restriction endonucleases by using the
TABLE 1. Rabies virus isolates used for probe generation
Viral strain Isolate Sourcea
AFXb 90N05850 Striped skunk, ONT
AFX 91N08281 Red fox, QUE
ERA 94N06762 Bovine, ONT
RAC V125 Raccoon, Fla.
WCS 00L00481 Striped skunk, W. Canada
BBCAN1 93N02994 Big brown bat, ONT
BBCAN2 93N04001 Big brown bat, ONT
BBCAN3 94N07206 Big brown bat, ONT
BBCAN4 90L01094 Big brown bat, BC
BBCAN5 88L01319 Big brown bat, BC
BBCAN6 93L01950 Big brown bat, BC
BBCAN7 72N04744 Big brown bat, ONT
LACAN (H) 92L02108 Hoary bat, ALTA
LACAN (R) 91N06832 Red bat, ONT
SHCAN 92L01830 Silver-haired bat, MAN
MYCAN (E) 93N09499 Red fox, PEI
MYCAN (W) 93L02414 Little brown bat, BC aONT, Ontario; QUE, Quebec; BC, British Columbia; ALTA, Alberta; MAN,
Manitoba; PEI, Prince Edward Island.
bUsed for generation of N-gene probe.
4344 NADIN-DAVIS ET AL. J. CLIN. MICROBIOL.
on May 15, 2020 by guest
http://jcm.asm.org/
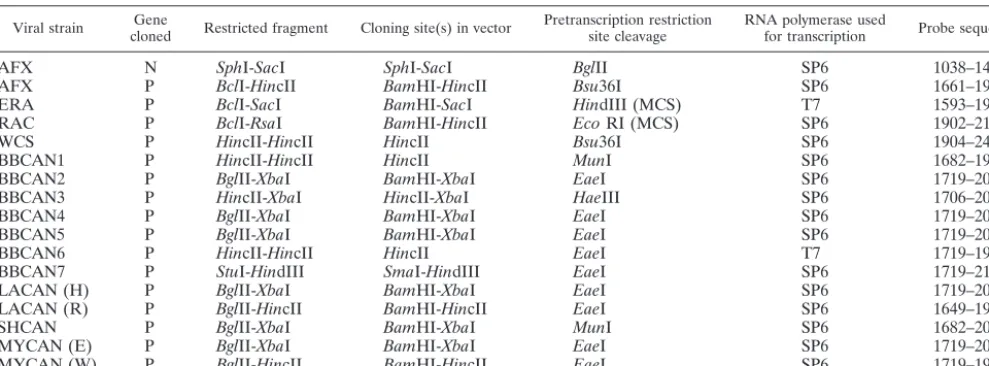
conditions recommended by the manufacturer, and the desired fragment was recovered by using the Wizard PCR purification kit after electrophoresis through low-melting-point agarose. Purified P-gene fragments were ligated into the pGEM3Zf(⫹) vector (Promega) previously cleaved by one or two restriction endonucleases contained within the multiple-cloning site (MCS). A complete description of the cloning strategy for all P-gene molecular clones used in these studies is provided in Table 2. In addition, a molecular clone of the N gene of the arctic fox strain (Table 2)—generated in a related vector, pGEM7Zf(⫹), from a clone described previously (19)—was used to generate a broadly cross-reactive probe that could be used as a positive control for the detection of all strains studied. Ligations were transformed intoEscherichia colistrain TG1 or JM109, and potentially positive clones were identified as white colonies by the
-galac-tosidase colorimetric assay with 5-bromo-4-chloro-3-indolyl--D
-galactopyrano-side substrate. In all cases the insertion fragment was confirmed by nucleotide sequencing across the cloning junction. Selected clones of each type were prop-agated in Luria-Bertani medium supplemented with 100g of ampicillin per ml, and plasmid stocks were prepared with a Wizard Midiprep kit (Promega).
[image:3.603.126.456.72.524.2]Preparation of cRNA probes. Plasmid DNA was cleaved by a restriction endonuclease at a site either within the P-gene insert or within the MCS to determine the size and specific sequence of the transcript. Following complete digestion, plasmid DNA was treated with proteinase K to remove any nuclease activity and was recovered by phenol-chloroform extraction and ethanol precip-itation. The DNA was dissolved in diethylpyrocarbonate (DEPC)-treated sterile water and maintained as a 1-mg/ml stock at⫺20°C.
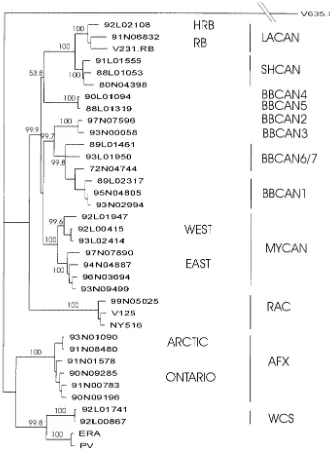
FIG. 1. Phylogeny of rabies virus strains in Canada. Nucleotide sequences for the coding region of the P genes for isolates representative of all rabies virus strains examined in these studies were aligned and used for phylogenetic prediction by using a neighbor joining algorithm with PHYLIP (version 3.5) software. Bootstrap values, determined by using 1,000 replicates of the data and shown as percentages, indicate the number of times that the clade to the right of the branch is predicted in the consensus tree. The antigenic groups represented by each branch are indicated to the right of the tree. A serotype 3 lyssavirus isolate, V635.S3 (22), was used as an outgroup; the distance of this isolate from all other specimens in the analysis has been reduced (indicated by the double reverse slashes) for illustrative purposes, but all other distances reflect the distance scale shown at the bottom left.
on May 15, 2020 by guest
http://jcm.asm.org/
Digoxigenin (DIG)-labeled cRNA probes were generated by in vitro transcrip-tion of 1g of linearized plasmid by using a RNA labeling kit according to the directions of the manufacturer (Roche Molecular Biochemicals) and either T7 or SP6 RNA polymerase, according to the orientation of the insert; all probes were of negative sense to detect mRNA sequences. After precipitation of the probe and recovery in DEPC-treated water, the probes were titrated for evaluation of labeling efficiency by spotting 1-l aliquots of 1-in-10 serial dilutions of the probe (1/100 to 1/100,000) onto a nitrocellulose membrane. The DIG ligand was de-tected colorimetrically by using an anti-DIG alkaline phosphatase (AP) conju-gate (Roche Molecular Biochemicals) diluted 1:5,000 and the substrate-chromo-gen solution containing 5-bromo-4-chloro-3-indolylphosphate (BCIP) and nitroblue tetrazolium (NBT) applied over a 90-min period. Only probes gener-ating a signal at a dilutionⱖ1/1,000 were used.
Southern and slot blotting.Southern blots were prepared by subjecting the RT-PCR products generated from the P genes of a variety of rabies viruses to electrophoresis through 1% agarose, followed by transfer to a nylon membrane by standard capillary methods. Slot blots were prepared by application of the RT-PCR products onto nitrocellulose by using a Vacu-Aid slot blot apparatus (Hybaid), as recommended by the manufacturer. The blots were prehybridized in prehybridization solution (5⫻SSC [1⫻SSC is 0.15 M NaCl plus 0.015 M sodium citrate], 1% casein, 0.1%N-lauroylsarcosine, 0.02% sodium dodecyl sulfate) at 70°C for 1 to 2 h. The probe was diluted 200- to 300-fold in hybridization solution (75% formamide, 10% dextran sulfate, 3⫻SSC, 1⫻Denhardt’s solution, 50 mM sodium phosphate buffer [pH 7.5], yeast tRNA carrier at 200g/ml), heated at 75°C for 3 min, and then immediately cooled on ice prior to incubation with the filter at 70°C overnight. After sequential washing in graded SSC solutions (see the description of the ISH procedure below), the blots were visualized by col-orimetric detection of the DIG ligand by using the anti-DIG AP conjugate and BCIP-NBT.
Preparation of rabies virus-infected reference tissues.Mice were inoculated intracerebrally with 10% suspensions of brain tissue recovered from animals infected with a predetermined strain of rabies virus. Animals were euthanized as soon as clinical signs were apparent. The brains were then removed, sliced into two halves, and fixed in 3% paraformaldehyde for 24 to 48 h at room tempera-ture before processing and embedding by standard methods at a temperatempera-ture not exceeding 60°C.
Section preparation.Embedded brains were cut into 5-m sections and mounted onto Superfrost Plus charged slides (Fisher) without the use of section adherents. The slides were baked at 60°C for 2 h, deparaffinized by using three 5-min changes in xylene, and cleared by using three 5-min changes in 100% ethanol (EtOH).
ISH. (i) Slide preparation and prehybridization.The general precautions taken to prevent RNase contamination of samples and reagents included baking of all glassware for 8 h at 180°C and DEPC treatment of all aqueous solutions, as recommended previously (25). The sections were hydrated by using one change of 70% EtOH and two changes of water and were then immersed in 0.2 N HCl at room temperature for 30 min. After the sections were rinsed in water
and then 1⫻PBS, sections were incubated in 1⫻PBS at 37°C for 20 min. The sections were incubated in a predetermined concentration of proteinase K (usu-ally 20g/ml for mouse brain tissues) in 50 mM Tris-HCl (pH 8.0)–2 mM CaCl2
buffer at 37°C for 30 min. After the slides were washed in three changes of water, they were incubated in 0.1 M triethanolamine (TEA) for 5 min and then in 0.1 M TEA containing 0.25% acetic anhydride for 10 min to block positively charged amino groups and to destroy any residual proteinase K activity. Finally, the slides were rinsed several times in water, incubated in 2⫻SSC at 70°C for 30 min, dehydrated in absolute EtOH, and dried.
(ii) Hybridization.Volumes of hybridization buffer in the range of 25 to 40l were used, depending on the size of the tissue section. After application of the probe (prepared as detailed above for Southern blotting), the liquid was covered by using sterile, siliconized, and baked glass coverslips; and the coverslips were sealed with rubber cement. The slides were sealed within a hybridization cham-ber humidified with 75% formamide and were then placed in an incubator set to 80°C. After 15 min, the temperature was reduced to 70°C for incubation over-night.
(iii) Posthybridization washing and conjugate application.The coverslips were removed; and the slides were washed sequentially in 2⫻, 1⫻, and 0.5⫻SSC for 15 min each and then in 0.1⫻SSC for 1 h at 80°C. The slides were incubated in blocking buffer (1% blocking reagent [Roche Molecular Biochemicals] in 100 mM Tris-HCl [pH 7.5], 150 mM NaCl) for 1 h at room temperature, and then 50
l of a 1:500 dilution of the anti-DIG AP conjugate was applied to each slide and the slides were incubated at room temperature for a minimum of 3 h or overnight at 4°C. Excess antibody was removed by using three 5-min changes of TBS buffer (10 mM Tris-HCl [pH 8.0], 15 mM NaCl) and the slides were equilibrated in AP developing buffer (10 mM Tris-HCl [pH 9.5], 10 mM NaCl, 5 mM MgCl2) for 10
min. Liquid was removed from the slide, and BCIP-NBT substrate was applied. The slides were monitored over several hours for the development of a purple precipitate in the positive controls. Once a clearly positive signal was observed for the positive controls, the reaction was stopped by removal of the substrate and washing of the slides in 1⫻TE (Tris-EDTA) buffer. A section was scored as positive if intracytoplasmic inclusions in the perikarya of neurons were stained; these inclusions could vary in size from fine dust to large polymorphic inclusion bodies. Any staining of other tissue elements was considered nonspecific and was scored as negative. Negative controls were monitored to ensure that they pre-sented minimal nonspecific staining and no staining of inclusion bodies in the perikarya of neurons anywhere on the section. Each hybridization was scored according to the intensity of specific neural cell staining from negative to weakly positive and very strongly positive; staining intensity was a measure of the degree of viral infectivity within the tissue section.
[image:4.603.46.541.81.264.2]IHC detection of rabies virus antigen.IHC detection of rabies virus antigen was performed with an avidin-biotin-peroxidase system as described previously (7). The criteria used for definition of positive and negative results for IHC were similar to those used for ISH. All mouse brains were confirmed to be rabies virus positive by IHC prior to their inclusion as controls for development of the ISH methodology. In addition, IHC was performed in parallel with ISH with all test
TABLE 2. Details of construction of rabies virus clones used during these studies
Viral strain clonedGene Restricted fragment Cloning site(s) in vector Pretranscription restrictionsite cleavage RNA polymerase usedfor transcription Probe sequencea
AFX N SphI-SacI SphI-SacI BglII SP6 1038–1425
AFX P BclI-HincII BamHI-HincII Bsu36I SP6 1661–1904
ERA P BclI-SacI BamHI-SacI HindIII (MCS) T7 1593–1967
RAC P BclI-RsaI BamHI-HincII EcoRI (MCS) SP6 1902–2149
WCS P HincII-HincII HincII Bsu36I SP6 1904–2460
BBCAN1 P HincII-HincII HincII MunI SP6 1682–1958
BBCAN2 P BglII-XbaI BamHI-XbaI EaeI SP6 1719–2062
BBCAN3 P HincII-XbaI HincII-XbaI HaeIII SP6 1706–2062
BBCAN4 P BglII-XbaI BamHI-XbaI EaeI SP6 1719–2062
BBCAN5 P BglII-XbaI BamHI-XbaI EaeI SP6 1719–2062
BBCAN6 P HincII-HincII HincII EaeI T7 1719–1958
BBCAN7 P StuI-HindIII SmaI-HindIII EaeI SP6 1719–2134
LACAN (H) P BglII-XbaI BamHI-XbaI EaeI SP6 1719–2062
LACAN (R) P BglII-HincII BamHI-HincII EaeI SP6 1649–1904
SHCAN P BglII-XbaI BamHI-XbaI MunI SP6 1682–2062
MYCAN (E) P BglII-XbaI BamHI-XbaI EaeI SP6 1719–2062
MYCAN (W) P BglII-HincII BamHI-HincII EaeI SP6 1719–1904
aBase numbers with reference to strain PV.
4346 NADIN-DAVIS ET AL. J. CLIN. MICROBIOL.
on May 15, 2020 by guest
http://jcm.asm.org/
sample tissues to define the location and the distribution of the virus within the tissue section.
Preparation of photomicrographs.Slides were viewed under an Olympus BX50 microscope and were photographed with a model DMCI Polaroid digital camera at either⫻10 or⫻20 magnification.
RESULTS
Genetic diversity of Canadian rabies viruses.To illustrate
the genetic diversity of the rabies viruses being targeted for discrimination by ISH, the results of phylogenetic analysis per-formed with the complete coding region of the P gene are shown (Fig. 1). In addition to all indigenous strains, we in-cluded in these studies the attenuated Evelyn-Rokitniki-Abels-eth (ERA) rabies virus strain, which is used as an oral vaccine for fox rabies control in Ontario (18). The residual pathoge-nicity of this strain does occasionally cause disease in wildlife and domestic species; indeed, in Ontario between 1991 and 1999 a total of nine cases of rabies diagnosed in two skunks, four red foxes, two raccoons, and one bovine were demon-strated by both antigenic and genetic typing methods to be a consequence of infection with the ERA strain (unpublished data). The tree presented in Fig. 1 illustrates that the P-gene locus sequence data allow the discrimination of the viral strains that were identified previously by both antigenic and genetic characterization at the N and G loci (21). The genetic distances illustrated in this tree were calculated from the complete P-gene sequences, but to maximize the discriminatory potential of strain-specific probes, the divergent central region of the P gene, corresponding to bases 1660 to 2060 of the PV reference strain of rabies virus (34), was targeted.
Generation of strain-specific probes.As described in
Mate-rials and Methods, a P-gene clone was generated from a
RT-PCR product produced from a representative of each viral type. A summary of the precise cloning details undertaken in each case is presented in Table 2; in addition, the bases of the rabies virus PV reference strain that correspond to the se-quence used to generate cRNA transcripts from each clone are shown. It was determined during the course of these studies that the inclusion of bases upstream of base 1600 (PV refer-ence strain numbering) was highly detrimental to the probe’s discriminatory capability, and hence, this region was avoided either by excluding it from the molecular clone or by cleavage of the plasmid with selected restriction endonucleases to pre-vent transcription through this region.
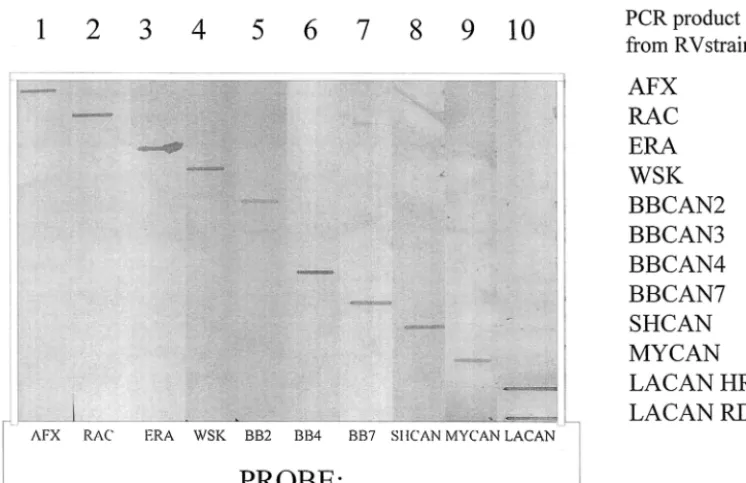
Testing of the specificities of these probes was initially per-formed by Southern and slot blotting hybridization. Prelimi-nary studies indicated the importance of the use of highly stringent hybridization conditions, obtained by use of a high formamide concentration, a high temperature, and a low salt concentration to achieve discriminatory conditions. Figure 2 illustrates the results of a slot blotting analysis of 10 of these probes and their interaction with the RT-PCR products gen-erated from one representative of each viral strain by using the optimized hybridization conditions described in Materials and Methods. Probes prepared from the three terrestrial strains and the ERA strain all hybridized only to their respective products, as did the probes prepared from the SHCAN and
Myotis virus types. However, the BBCAN2-specific probe
bound strongly to BBCAN2 sequences and more weakly, but consistently, to BBCAN3 sequences. Finally, theLasiurus ge-nus-specific probe bound equally well to the sequences of iso-lates from both red and hoary bats and faintly to the SHCAN sequence. The BBCAN1-specific probe bound specifically to products of the BBCAN1 antigenic type, while the BBCAN3-FIG. 2. Slot blotting analysis showing the specificities of selected P-gene-specific probes. Ten replicate slot blots onto which P-gene PCR products amplified from the viral RNA of the indicated strains had been applied were hybridized with P-gene-specific probes by using the conditions specified in Materials and Methods.
on May 15, 2020 by guest
http://jcm.asm.org/
[image:5.603.109.482.70.311.2]specific probe cross-reacted to sequences of both the BBCAN2 and the BBCAN3 strains of rabies virus as well as weakly to MYCAN strain sequences (data not shown). Probes specific for BBCAN4 and BBCAN5 strains recognized the PCR prod-ucts of these groups exclusively but could not discriminate between the two types, and similarly, the probes developed for BBCAN6 and BBCAN7 strains exhibited specificity for these types but did not distinguish them (data not shown).
The ability of each of these probes to recognize RT-PCR products of the rabies virus P gene generated from several viral isolates of each type was also verified by Southern blotting analysis. For this purpose, RT-PCR products were generated from the following collection of isolates: 10 AFX isolates, including representatives of all five genetic variants identified over the Canadian range of this strain as described previously (19); 5 RAC isolates from Pennsylvania, New York, and On-tario; 3 ERA isolates obtained from field specimens; 12 iso-lates of big brown bat origin, including at least 1 isolate of all antigenic variants (in some cases only small numbers of the different variants have been isolated); 5 SHCAN isolates; 6 isolates fromMyotisspecies of bats from both the eastern and the western parts of Canada; and 6 isolates from bats of the
Lasiurusgenus (3 from red bats and 3 from hoary bats). These
experiments demonstrated that the probes selected detected all isolates within the respective groups while maintaining group specificity (data not shown). It was concluded from these blotting analyses that this probe panel would allow discrimina-tion between the following rabies virus strains: ERA, AFX, WSK, RAC, BBCAN1, BBCAN2-BBCAN3, BBCAN4-BBCAN5, BBCAN6-BBCAN7, SHCAN, MYCAN, and LACAN. The identical reactivities of the two probes specific for LACAN (H) and LACAN (R) precluded discrimination of viruses within the LACAN antigenic type, although phyloge-netic examination placed hoary and red bat isolates on sepa-rate branches of the same clade (21). Moreover, the close genetic similarity (Fig. 1) of several of the BBCAN groups clearly precluded their discrimination by this technique.
Use of P-gene probes for ISH.To examine the possibility of
using these strain-specific probes for ISH, the probes were tested by using rabies virus-positive brain tissue prepared from mice experimentally inoculated with a selection of isolates representative of all strains under study. Additional brain tis-sues from uninoculated animals were also used as negative controls. During the course of this phase of the studies, a molecular clone with a highly conserved region of the rabies virus N gene (Tables 1 and 2) that could be transcribed into a broadly cross-reactive probe that recognized all strains under study was found to be an invaluable positive control. This N-gene clone was used to demonstrate that probes targeting a positive-sense (primarily mRNA) sequence were more sensi-tive than those targeting a negasensi-tive-sense (genomic) sequence. Furthermore, this probe was useful for determination of the precise tissue solubilization conditions (e.g., proteinase K treatment) required to optimize probe penetration and signal strength.
Figure 3 illustrates the staining patterns observed by using a selection of rabies virus-specific probes applied to mouse brain tissues infected with a variety of rabies virus strains. It was notable that in serially cut sections there was an excellent correspondence between the pattern of rabies virus antigen
detected by IHC (Fig. 3D, H, and L) and the pattern obtained by using both the N-gene probe (Fig. 3A, E, and I) and strain-specific P-gene probes (Fig. 3B, F, and J) for viral mRNA, although the antigenic signals obtained with the probes were always stronger than those achieved by ISH. In all cases stain-ing was restricted to the cytoplasms of selected neurons in a pattern consistent with the usual distribution of the virus in infected brain tissue. No signal was apparent in uninfected brain tissue, regardless of the probe used (data not shown). Only those tissues infected with a viral strain corresponding to the specificity of the P-gene probe applied showed strong pur-ple staining (Fig. 3B, F, and J); tissues hybridized with a probe that did not correspond to the variant present gave negative patterns (Fig. 3C, G, and K). It was thus concluded that these P-gene-specific probes can be used in an ISH protocol to dis-criminate between various rabies virus strains.
Evaluation of ISH with isolates also typed by traditional
methods.Two recently submitted specimens were used to
val-idate the utility of the ISH procedure for the typing of rabies viruses from other mammalian species. The first specimen was from a rabid cat (submission 03N00756) from Halifax, Nova Scotia, which was originally submitted as formalin-fixed mate-rial, but fresh brain material was subsequently located and provided. The second specimen was from a human from Van-couver, British Columbia, who was diagnosed with rabies post-mortem; both fixed and frozen brain materials were provided. Sections of fixed brain material from each case were evaluated by ISH, and the results are summarized in Table 3. Figure 4 illustrates some of the data for the human specimen only. In both cases the specimens yielded hybridization patterns con-sistent with the presence of a rabies virus of the MYCAN strain. Additional studies with these specimens from Nova Scotia and British Columbia by standard antigenic and genetic procedures, as detailed previously (21), also confirmed the presence of the MYCAN strain of the east and west varieties, respectively, in these tissues (data not shown). These observa-tions confirm the utility of the ISH method in correctly iden-tifying rabies virus variants in mammalian species other than mice.
Use of rabies virus strain-specific ISH to type archival
sam-ples.To explore the practical utility of the ISH method, it was
applied to two equine specimens both received at this labora-tory in 1992 for rabies diagnosis subsequent to brain fixation. One (specimen 92N08767) originated from Truro, Nova Sco-tia, and the second (specimen 92N10245) was from Stanley, New Brunswick. Both of these specimens had been found to be rabies virus positive by IHC, but both were refractory to all viral typing methods available at that time. However, since neither of these provinces was known to harbor any terrestrial rabies viruses during that period, it was presumed that the infections were likely a consequence of viral spillover from rabid bats.
During initial ISH studies with these brain specimens by using the N-gene-specific probe, it became evident that mod-ification of the standard ISH protocol was required. In partic-ular, a more vigorous proteinase K treatment (50 g/ml at 45°C for 30 min) was necessary to optimize probe penetration and thereby generate sufficiently strong signal intensities. Moreover, due to the sparsity of viral infection of the brain tissue received for specimen 92N10245 (from New Brunswick),
4348 NADIN-DAVIS ET AL. J. CLIN. MICROBIOL.
on May 15, 2020 by guest
http://jcm.asm.org/
FIG.
3.
Use
of
P-gene-speci
fic
probes
and
ISH
to
discriminate
between
rabies
virus
variants
used
to
infect
mouse
brains.
Sections
were
prepared
from
the
brain
tissues
of
mice
which
had
been
experimentally
infected
with
the
following
rabies
viruses:
BBCAN1
rabies
virus
variant
(A
to
D),
MYCAN
rabies
virus
variant
(E
to
H),
and
AFX
rabies
virus
variant
(I
to
L).
The
following
rabies
virus
P-gene-speci
fic
probes
were
used
in
the
ISH
protocol:
rabies
virus
N-gene-speci
fic
probe
(A,
E,
and
I),
BBCAN1-speci
fic
probe
(B,
G,
and
K),
MYCAN-speci
fic
probe
(C
and
F),
and
AFX-speci
fic
probe
(J).
Viral
antigen
was
detected
by
IHC
in
panels
D,
H,
and
L.
Magni
fications,
⫻
9(
At
oD
)a
nd
⫻
19
(E
to
L).
on May 15, 2020 by guest
http://jcm.asm.org/
segments of the spinal cord, which exhibited more extensive levels of viral infection, were also processed for ISH analysis. A summary of all results obtained from these studies is presented in Table 4. Note that only a subset of probes was selected after due consideration of prior knowledge of the geographical dis-tribution and frequencies of many of the variants circulating in chiropteran species; for example, the BBCAN4 and BBCAN5 strains have been identified only in British Columbia and BBCAN6 and BBCAN7 strains are rare. Specimen 92N08767 (from Nova Scotia) generated neuron-specific staining only with the N-gene- and SHCAN-specific probes. Specimen 92N10245 (from New Brunswick) yielded strong ISH signals
only with the N-gene- and BBCAN2-specific probes, with a weaker signal obtained with the BBCAN1-specific probe and some faint signal obtained with the AFX-specific probe; in particular, the BBCAN2-specific probe gave intense staining in the cytoplasm and axonal projections for a small cluster of neurons in the brain. Sequential sections of spinal cord from this animal showed similar staining patterns with the N-gene-and BBCAN2-specific probes (data not shown). All other probes tested failed to generate any signal with either of these specimens. ISH controls, which consisted of mouse brain ref-erence tissues analyzed alongside these unknowns, all gave the expected hybridization patterns. From these findings it was concluded that the SHCAN (for the specimen from Nova
[image:8.603.43.283.89.197.2]Sco-FIG. 4. Typing of rabies virus strain in human specimen from BC by ISH. Brain sections were assayed as follows: ISH with MYCAN-specific probe (A); ISH with N-gene-specific probe (B); IHC detection of rabies virus antigen (C); and ISH with AFX-specific probe (D), SHCAN-specific probe (E), and BBCAN4-specific probe (F). Magnification⫻9.
TABLE 3. Signal pattern generated for two rabies virus-positive specimens evaluated by ISH
Probe
Pattern for the following specimena:
03N00756
(cat, Nova Scotia) (human, British Columbia)03N01069
N gene (⫹⫹⫹) (⫹⫹⫹)
AFX (⫺) (⫺)
BBCAN2 (⫺) ND
BBCAN4 ND (⫺)
MYCAN (E) (⫹⫹⫹)b ND
MYCAN (W) ND (⫹⫹⫹)b
SHCAN (⫺) (⫺)
aEach of the two specimens was evaluated for binding of the indicated probe,
as detected by production of purple staining through the AP-BCIP-NBT color-imetric system (see Materials and Methods). Results were scored as follows: ND, not determined (some specimen-probe combinations were not tested because of the regionally limited distribution of the BBCAN2, BBCAN4, and MYCAN variant strains and the limited availability of tissues); (⫺), no staining; (⫹), low levels of rabies-specific staining; (⫹⫹⫹), very strong levels of rabies virus-specific staining.
[image:8.603.300.542.97.205.2]bThe strain identified as responsible for the infection.
TABLE 4. Signal pattern generated for two archival rabies virus-positive horse specimens evaluated by ISH with
strain-specific probes
Probe
Pattern for the following specimen:a
92N08767
(Nova Scotia) (New Brunswick)92N10245
N gene (⫹⫹⫹) (⫹⫹⫹)
AFX (⫺) (⫹)
BBCAN1 (⫺) (⫹)
BBCAN2 (⫺) (⫹⫹)b
LACAN (⫺) (⫺)
MYCAN (⫺) (⫺)
SHCAN (⫹⫹⫹)b (⫺)
aEach of the two specimens was evaluated for binding of the indicated probe,
as detected by production of purple staining through the AP-BCIP-NBT color-imetric system (see Materials and Methods). Results were scored as follows: (⫺), no staining; (⫹), low levels of rabies-specific staining; (⫹⫹) or (⫹⫹⫹), strong to very strong levels of rabies-specific staining.
bThe strain identified as responsible for the infection.
4350 NADIN-DAVIS ET AL. J. CLIN. MICROBIOL.
on May 15, 2020 by guest
http://jcm.asm.org/
[image:8.603.49.541.454.698.2]tia) and BBCAN2-BBCAN3 (for the specimen from New Brunswick) strains were responsible for these two cases of rabies.
DISCUSSION
Previously, ISH has been applied to rabies virus-infected tissues for tracing of the presence of rabies virus nucleic acids in mouse and human brains, primarily for diagnostic purposes (14, 36). In those previous studies, no attempt was made to discriminate the rabies virus strains responsible for the infec-tions; indeed, probes whose sequences were based on con-served regions of the rabies virus genome were used to permit detection of a broad range of both rabies viruses and rabies virus-related viruses (36). The use of ISH for rabies virus strain discrimination is thus unique to this study, although this meth-odology has been applied to the differentiation of distinct ge-notypes of other viruses; for example, Larochelle and Magar (16) developed probes to discriminate the North American and European porcine reproductive and respiratory syndrome vi-rus genotypes by ISH applied to formalin-fixed tissues.
Several parameters of the ISH technique must be optimized for its successful application to rabies virus strain typing. A critical factor in this regard is the choice of target sequence and the type of probe used. Our initial attempts with DIG-labeled synthetic oligonucleotides gave poor results, despite the tar-geting of short P-gene sequences of high intertypic variability. Such probes lacked specificity under hybridization conditions that permitted adequate signal strength and frequently yielded nonspecific nuclear staining. The use of DIG-labeled RNA probes for rabies virus detection by ISH by other investigators (13, 36) prompted investigation of this probe type. The RNA-specific probes thus developed targeted the divergent central section of the P gene that encompasses bases 1660 to 2060 of the PV reference strain (34). For the discrimination of viruses that are genetically quite distinct (e.g., AFX and bat strains), the precise choice of sequence included in the probe was not so critical, but to differentiate between closely related strains (e.g., those of big brown bats), probes lacking bases upstream of base 1680 were found to confer superior discriminatory properties. While the P gene proved to be a suitable target for viral typing, an N-gene-specific probe (developed from the sequence of strain AFX) which was broadly cross-reactive to all of the strains under study proved invaluable for optimiza-tion of various ISH parameters during method development. The relatively high strength of RNA-RNA hybrids permits the use of these cRNA probes under highly stringent conditions, a factor clearly critical to the method’s success. Optimization of the concentration of the applied probe was also important, and the lowest concentration which yielded sufficiently intense staining was used; higher concentrations were sometimes found to reduce probe specificity. Other factors significantly affecting the procedure’s success included probe length and sense. Probes of 200 to 400 bases in length gave optimal results by ISH with probes significantly larger than this, giving increas-ingly weaker signals, presumably due to poor tissue penetra-tion. The negative-sense sequence-specific probes routinely used here target the positive-sense sequence, which is predom-inantly mRNA. The typical rhabdovirus transcription pattern (3) allows the synthesis of relatively large amounts of mRNAs
for the 3⬘proximal N and P genes compared to the amounts obtained for genes located farther away from the 3⬘terminus of the genome, and thus, these genes provide the highest level of sensitivity for this approach. The use of positive-sense se-quence-specific probes for detection of genomic RNA was less sensitive, perhaps due to either the smaller amount of this molecule in infected cells or the significantly reduced accessi-bility of the probe to this target due to its encapsidation by the nucleoprotein, especially after tissue fixation and cross-linking of cell components. Nonspecific staining with the anti-DIG conjugate, particularly within endothelial cells lining blood ves-sels, was also observed occasionally, even in uninfected tissues, probably due to the presence of endogenous AP in these tis-sues. Careful evaluation of the cellular distribution of all stain-ing is thus essential to identify such artifacts.
Another critical step in the procedure is the optimization of the proteinase K treatment. There is a need to balance high levels of tissue permeabilization to allow probes to access their target and maintenance of RNA target integrity to maximize signal strength. Thus, the standardized proteinase K treatment identified for use with our mouse reference material, all of which was processed under similar conditions, was not appriate for the two archival equine specimens, and pretest teinase K titrations were necessary to optimize the ISH pro-cedure for use with the latter specimens. It is possible that these opposing needs could confound the application of this procedure to highly processed specimens; however, ISH was successfully applied to rabies virus typing in this instance and identified the strains responsible for infection as two distinct bat strains, SHCAN (for a specimen from Nova Scotia) and BBCAN2-BBCAN3 (for a specimen from New Brunswick). Furthermore, this ISH procedure was the only method that could be satisfactorily applied in these two cases. However, the inclusion of a large number of controls is essential when un-known specimens are examined. As demonstrated with our experience with the specimen from New Brunswick, extended incubation in the AP chromogen substrate during the final step of probe detection, which may sometimes be necessary to ob-tain clearly positive signals, can result in the development of weak signals as a result of the low-level residual cross-reactiv-ities of certain probes (data not shown). This could lead to an incorrect typing result if appropriate controls (comprising un-infected brain and mouse reference brains un-infected with strains for which each of the probes used is specific) are not included within each test.
We have demonstrated the utility of the ISH approach for the typing of Canadian rabies viruses which circulate in several geographically overlapping and temporally dynamic ranges. However, the development of additional probes appropriate for the detection of other strains of this virus would allow the application of this technology to many other areas of the world. Such an application might be of particular benefit in developed countries where human rabies is rare and is diagnosed only postmortem, but it might also be of considerable value in tropical countries where preservation of infected tissues by means other than formalin fixation can be a significant prob-lem. Indeed, many regions of Latin America harbor multiple rabies virus types in geographically overlapping host reservoirs (12, 22), and improved epidemiological information on these
on May 15, 2020 by guest
http://jcm.asm.org/
types would be a significant help with their control. Application of this ISH methodology could play some role in this endeavor.
ACKNOWLEDGMENTS
We thank F. Muldoon for technical assistance with the IHC analyses and passaging of rabies virus isolates in mice. We also acknowledge the provision of brain material from a human case of rabies by M. Petric (BCCDC) and I. Mackenzie (Vancouver General Hospital).
REFERENCES
1. Badrane, H., C. Bahloul, P. Perrin, and N. Tordo.2001. Evidence of two immuno-pathologically distinct lyssavirus phylogroups. J. Virol.75:3268– 3276.
2. Badrane, H., and N. Tordo.2001. Host-switching inLyssavirushistory from theChiropterato theCarnivoraorders. J. Virol.75:8096–8104.
3. Banerjee, A. K., and S. Barik.1992. Gene expression of vesicular stomatitis virus genome RNA. Virology188:417–428.
4. Bourhy, H., K. Bachir, and N. Tordo.1993. Molecular diversity of the Lyssavirus genus. Virology194:70–81.
5. Dean, D. J., M. K. Abelseth, and P. Atanasiu.1996. The fluorescent antibody test, p. 88–95.InF. X. Meslin, M. M. Kaplan, and H. Koprowski (ed.), Laboratory techniques in rabies, 4th ed. World Health Organization, Ge-neva, Switzerland.
6. Dries, V., I. von Both, M. Mu¨ller, G. Gerken, P. Schirmacher, M. Odenthal, R. Bartenschlager, U. Drebber, K. H. Meyer zum Bu¨schenfeld, and H. P. Dienes.1999. Detection of hepatitis C virus in paraffin-embedded liver bi-opsies of patients negative for viral RNA in serum. Hepatology29:223–229. 7. Fekadu, M., P. W. Greer, F. W. Chandler, and D. W. Sanderlin.1988. Use of the avidin-biotin peroxidase system to detect rabies antigen in formalin-fixed paraffin-embedded tissues. J. Virol. Methods19:91–96.
8. Felsenstein, J.1993. PHYLIP: phylogeny inference package, version 3.52c. University of Washington, Seattle.
9. Gruber, A. D., I. M. Greiser-Wilke, L. Haas, M. Hewicker-Trautwein, and V. Moennig.1993. Detection of bovine viral diarrhea virus RNA in formalin-fixed, paraffin-embedded brain tissue by nested polymerase chain reaction. J. Virol. Methods43:309–319.
10. Gruber, A. D., V. Moennig, M. Hewicker-Trautwein, and G. Trautwein.1994. Effect of formalin fixation and long-term storage on the detectability of bovine viral-diarrhoea-virus (BVDV) RNA in archival brain tissue using polymerase chain reaction. J. Vet. Med. Ser. B41:654–661.
11. Guerrero, R. B., K. P. Batts, D. J. Brandhagen, J. J. Germer, R. G. Perez, and D. H. Persing.1997. Effects of formalin fixation and prolonged block storage on detection of hepatitis C virus RNA in liver tissue. Diagn. Mol. Pathol. Am. J. Surg. Pathol.6(Pt. B):277–281.
12. Ito, M., Y. T. Arai, T. Itou, T. Sakai, F. H. Ito, T. Takasaki, and I. Kurane.
2001. Genetic characterization and geographic distribution of rabies virus isolates in Brazil: identification of two reservoirs, dogs and vampire bats. Virology284:214–222.
13. Jackson, A. C.1992. Detection of rabies virus mRNA in mouse brain by using in situ hybridisation with digoxigenin-labelled RNA probes. Mol. Cell. Probes6:131–136.
14. Jackson, A. C., and W. H. Wunner.1991. Detection of rabies virus genomic RNA and mRNA in mouse and human brains by using in situ hybridisation. J. Virol.65:2839–2844.
15. Kissi, B., N. Tordo, and H. Bourhy.1995. Genetic polymorphism in the rabies virus nucleoprotein gene. Virology209:526–537.
16. Larochelle, R., and R. Magar.1997. Differentiation of North American and European porcine reproductive and respiratory syndrome virus genotypes by in situ hybridisation. J. Virol. Methods68:161–168.
17. Le Mercier, P., Y. Jacob, and N. Tordo.1997. The complete Mokola virus genome sequence: structure of the RNA-dependent RNA polymerase. J. Gen. Virol.78:1571–1576.
18. MacInnes, C. D.1998. Eliminer la rage du Sud de l’Ontario. Med. Vet. Quebec28:115–118.
19. Nadin-Davis, S. A., G. A. Casey, and A. Wandeler.1993. Identification of regional variants of the rabies virus within the Canadian province of Ontario. J. Gen. Virol.74:829–837.
20. Nadin-Davis, S. A., M. Sheen, M. Abdel-Malik, L. Elmgren, J. Armstrong, and A. I. Wandeler.2000. A panel of monoclonal antibodies targeting the rabies virus phosphoprotein identifies a highly variable epitope of value for sensitive strain discrimination. J. Clin. Microbiol.38:1397–1403.
21. Nadin-Davis, S. A., W. Huang, J. Armstrong, G. A. Casey, C. Bahloul, N. Tordo, and A. I. Wandeler.2001. Antigenic and genetic divergence of rabies viruses from bat species indigenous to Canada. Virus Res.74:139–156. 22. Nadin-Davis, S. A., M. Abdel-Malik, J. Armstrong, and A. I. Wandeler.2002.
Lyssavirus P gene characterisation provides insights into the phylogeny of the genus and identifies structural similarities and diversity within the encoded phosphoprotein. Virology298:286–305.
23. Page, R. D. M.1996. TREEVIEW: an application to display phylogenetic trees on personal computers. Comput. Appl. Biosci.12:357–358. 24. Rupprecht, C. E., B. Dietzschold, W. H. Wunner, and H. Koprowski.1991.
Antigenic relationships of lyssaviruses, p. 69–100.InG. M. Baer (ed.), The natural history of rabies, 2nd ed. CRC Press, Inc., Boca Raton, Fla. 25. Sambrook, J., E. F. Fritsch, and T. Maniatis.1989. Molecular cloning: a
laboratory manual, 2nd ed., p. 7.3–7.5. Cold Spring Harbor Laboratory Press, Cold Spring Harbor, N.Y.
26. Schwarz, T. F., S. R. Zaki, S. Morzunov, C. J. Peters, and S. T. Nichol.1995. Detection and sequence confirmation of Sin Nombre virus RNA in paraffin-embedded human tissues using one-step RT-PCR. J. Virol. Methods51:349– 356.
27. Smith, J. S., and A. A. King.1996. Monoclonal antibodies for the identifi-cation of rabies and non-rabies lyssaviruses, p. 145–156.InF. X. Meslin, M. M. Kaplan, and H. Koprowski, (ed.) Laboratory techniques in rabies, 4th ed. World Health Organization, Geneva, Switzerland.
28. Smith, J. S., L. A. Orciari, P. A. Yager, H. D. Seidel, and C. K. Warner.1992. Epidemiologic and historical relationships among 87 rabies virus isolates as determined by limited sequence analysis. J. Infect. Dis.166:296–307. 29. Smith, J. S., L. A. Orciari, and P. A. Yager.1995. Molecular epidemiology of
rabies in the United States. Semin. Virol.6:387–400.
30. Soguero, C., E. Campo, T. Ribalta, J. M. Sa´nchez-Tapias, J. C. Sa´iz, and M. Bruguera.2000. Assessment of genotype and molecular evolution of hepa-titis C virus in formalin-fixed paraffin-embedded liver tissue from patients with chronic hepatitis C virus infection. Lab. Investig.80:851–856. 31. Thompson, J. D., T. J. Gibson, F. Plewniak, F. Jeanmougin, and D. G.
Higgins.1997. The ClustalX windows interface: flexible strategies for mul-tiple sequence alignment aided by quality analysis tools. Nucleic Acids Res.
24:4876–4882.
32. Tordo, N.1996. Characteristics and molecular biology of the rabies virus, p. 28–51.InF. X. Meslin, M. M. Kaplan, and H. Koprowski, (ed.) Laboratory techniques in rabies, 4th ed. World Health Organization, Geneva, Switzer-land.
33. Tordo, N., K. Charlton, and A. Wandeler.1998. Rhabdoviruses: rabies, p. 666–692.InL. H. Collier (ed.) Topley and Wilson’s microbiology and mi-crobial infections. Arnold Press, London, United Kingdom.
34. Tordo, N., O. Poch, A. Ermine, G. Keith, and F. Rougeon.1986. Walking along the rabies genome: is the large G-L intergenic region a remnant gene? Proc. Natl. Acad. Sci. USA83:3914–3918.
35. Wandeler, A. I., R. C. Rosatte, D. Williams, T. K. Lee, K. F. Gensheimer, J. T. Montero, C. V. Trimarchi, D. L. Morse, M. Eidson, P. F. Smith, J. L. Hunter, K. A. Smith, R. H. Johnson, S. R. Jenkins, and C. Berryman.2000. Update: raccoon rabies epizootic—United States and Canada, 1999. Morb. Mortal. Wkly. Rep.49:31–35.
36. Warner, C. K., S. G. Whitfield, M. Fekadu, and H. Ho.1997. Procedures for reproducible detection of rabies virus antigen mRNA and genome in situ in formalin-fixed tissues. J. Virol. Methods67:5–12.
4352 NADIN-DAVIS ET AL. J. CLIN. MICROBIOL.