0095-1137/05/$08.00⫹0 doi:10.1128/JCM.43.9.4336–4341.2005
Copyright © 2005, American Society for Microbiology. All Rights Reserved.
Use of Two Selective Media and a Broth Motility Test Can Aid
in Identification or Exclusion of
Bacillus anthracis
Vicki A. Luna,
1* K. Kealy Peak,
1William O. Veguilla,
1Frank Reeves,
2Lea Heberlein-Larson,
2Andrew C. Cannons,
1Phil Amuso,
1,2and Jacqueline Cattani
1Center for Biological Defense, College of Public Health, University of South Florida,1and Florida Department of Health, Bureau of Laboratories,2Tampa, Florida
Received 21 March 2005/Returned for modification 6 May 2005/Accepted 14 June 2005
During the anthrax attack of 2001, the Florida Department of Health (FDOH) Bureau of Laboratories in
Tampa received hundreds of isolates suspected of beingBacillus anthracis. None were confirmed to be B.
anthracis since most isolates were motile and not even in the Bacillus cereusgroup. Although the sentinel laboratories now send fewer isolates to FDOH laboratories, should another attack occur the number of isolates submitted would likely increase dramatically, and this upsurge would seriously challenge personnel who are expected to be busy examining an increased number of environmental samples. We examined two selective and differential growth media and alternative motility methods that could be used to streamline the processing of suspicious isolates. Of 60 isolates previously sent to the FDOH laboratory, 56 were endospore-forming
gram-positive rods and only 7 grew on mannitol-egg yolk-polymyxin B agar and/or theAnthracischromogenic
agar. Microscopic observation of early-log-phase growth (2 to 3 h) in a shaking broth was the best method to detect motility in 40 isolates that appeared nonmotile in the motility media investigated. One of these growth
media and microscopic examination of shaken broth cultures can be used to show that an isolate is notB.
anthracis before expensive molecular and antibody-based tests are performed. By doing so, costs could be reduced and analysis time shortened.
In 2001, Bacillus anthracis, the etiologic agent of anthrax,
was used as a bioterrorism weapon. When the U.S. Postal
Service unwittingly distributed B. anthracis spores that had
been placed inside common envelopes to several locations, these spores infected a total of 22 people, not all of whom were the primary targets. Except for an elderly woman in Connect-icut and a child visiting the American Broadcasting Company, the remainder appeared to be victims who worked in offices and buildings that became contaminated when spore-laden envelopes were handled (8, 9). During this time, hundreds of thousands of specimens were sent to designated laboratories throughout the United States that were part of the National Laboratory Response Network (LRN), an integrated system of Sentinel, Reference, and National laboratories established by the Centers for Disease Control and Prevention (CDC) (2, 3). There were three Florida State Department of Health (FDOH) laboratories that had biosafety level 3 facilities des-ignated to receive these potentially dangerous specimens. The FDOH Tampa Laboratory received and analyzed 1046 envi-ronmental samples and at the same time received hundreds of
clinical isolates suspected of beingB. anthracis from sentinel
laboratories across west central Florida. The FDOH Tampa Laboratory evaluated these isolates for hemolysis and motility
and then performed tests specific for B. anthracis such as
gamma phage susceptibility, cell wall and capsule detection by direct fluorescence antibody tests (DFA) and amplification of
targeted DNA sequences. None of the isolates received since,
or during the 2001 event, has proven to beB. anthracis.
Since that time, sentinel laboratories continue to send iso-lates, although in lower numbers. Currently, the FDOH Tampa laboratory receives an average of two isolates per week. Still, it is easy to imagine how quickly the laboratory system could become overwhelmed should another real or perceived threat occur. Shortly after the anthrax attacks, the FDOH Tampa laboratory noticed that the majority of the isolates received from the sentinel laboratories were hemolytic and/or motile when retested by FDOH personnel or they were nega-tive for the confirmatory and specific tests. Because this prob-lem has persisted, we wanted to examine whether the use of two selective and differential media (mannitol-egg
yolk-poly-myxin B [MEP] agar and R&F Anthracischromogenic agar
[ChrA]), and/or alternative motility methods, could streamline the processing of suspicious isolates. If proven effective, the
reference laboratories could use these methods to rule outB.
anthracisbefore expensive molecular and antibody-based tests were performed. Although these modifications if used in sen-tinel laboratories could potentially reduce the numbers of iso-lates being sent to the LRN Reference laboratories, the sen-tinel laboratories should continue to forward all suspicious isolates until the CDC makes a recommendation.
We examined and further identified 60 non-B. anthracis
iso-lates that had previously been sent to the FDOH Tampa Lab-oratory with the request to “rule-in” or confirm that the isolate was B. anthracis. Colonies of these isolates were inoculated onto selective and differential growth media, tube motility me-dia, and broth media (for phase microscopy of early log phase growth). The information obtained leads us to offer some
sug-* Corresponding author. Mailing address: Center for Biological De-fense, University of South Florida, 3602 Spectrum Blvd., Tampa, FL 33612. Phone: (813) 974-3873. Fax: (813) 974-1479. E-mail: vluna @bt.usf.edu.
4336
on May 15, 2020 by guest
http://jcm.asm.org/
gestions to the CDC and LRN laboratories. We suggest that the reference laboratories consider adding one of these media and/or the modified motility testing in log-phase broth in order to potentially reduce their workload and the cost of testing, since fewer isolates would warrant further testing by the more expensive antibody- and amplification-based methods in cur-rent use. Any savings in both time and money would be most important in the event of another perceived or confirmed threat of anthrax bioterrorism.
MATERIALS AND METHODS
Bacteria.Sixty bacterial isolates suspected of beingB. anthraciswere sent from microbiology laboratories in west central Florida to the FDOH Tampa Labora-tory for confirmation. After it was determined by LRN protocols (gamma phage susceptibility tests, direct fluorescence antibody tests, and PCR analysis) that none wereB. anthracis, the isolates were shared with the University of South Florida Center for Biological Defense for further examination. Control organ-isms used were obtained from the American Type Culture Collection (ATCC), CDC, and the U.S. Department of Agriculture (USDA) Agricultural Research Service Culture Collection.
Growth conditions and biochemical identification of bacteria.Isolates were evaluated for cell morphology with Gram stain reaction (controlsStaphylococcus aureusATCC 25923 (⫹),Escherichia coliATCC 25922 (⫺), and colony mor-phology with special emphasis on the hemolytic activity on Trypticase soy agar supplemented with 5% sheep red blood cells (BA) (controlsBacillus cereus
ATCC 14579 [beta-hemolytic] andB. megateriumATCC 14581 [nonhemolytic]) (7, 12, 13). Catalase production was tested with 3% hydrogen peroxide (controls
S. aureusATCC 29213 (⫹);Enterococcus faecalisATCC 29212 (⫺)) (4, 6, 7). Endospore production was induced by incubation at 30°C for 48 h on nutrient agar with 5 mg of manganese sulfate/liter or on modified nutrient sporulation media with phosphate agar (NSMP Medium #1209; ATCC, Manassas, VA) (1). Phase-contrast microscopy was used to determine endospore shape, location, swelling of the cell, and the presence of parasporal crystals, bodies, or inclusions (controlsB. cereusATCC 14579 [no parasporal bodies],B. thuringiensisATCC 35646 [parasporal crystals],B. megateriumATCC 14581 (inclusions), andListeria monocytogenesATCC 9525 [non-spore-former]) (11). Fermentation and bio-chemical studies were conducted by using the API 50 CH panel with API CHB/E Medium (bioMerieux, Inc., Hazelwood, MO), as well as 12 tests from the bio-Merieux API 20E kit for the presence of-galactosidase (ONPG [o
-nitrophenyl--D-galactopyranoside]), arginine dihydrolase, lysine decarboxylase, ornithine decarboxylase, gelatinase, tryptophan deaminase, urease, citrate utilization, H2S
production, indole production, acetoin production, and nitrate reduction. These tests were performed and interpreted according to the manufacturer’s instruc-tions (controlsB. cereusATCC 14579,B. megateriumATCC 14581,B. subtilis
ATCC 23059, andE. coliATCC 25922), and the resultant profiles were analyzed with the APILAB Plus Identification Program V.3.3.3/3.0 (bioMerieux, Inc.).
Motility tests.Motility was determined by simultaneous stab inoculation into tubes of the classical formula of motility test medium with or without 2,3,5-triphenyltetrazolium chloride (TTC) dye (REMEL, Inc., Lenexa, KS). Single line stabs with a narrow gauge nichrome needle were used to control inoculum size and reduce oxygen intrusion into the semisolid medium. The efficacy of the two formulations to detect motility was observed and compared after 24 and 48 h at two incubation temperatures: 30 and 35°C. Interpretation of the test as positive or negative for motility followed the manufacturer’s technical guidelines. A positive test showed diffuse growth away from the stab line of inoculation, evidenced by turbidity, cloudiness, or feathery protuberances extending laterally throughout the medium. A negative motility was defined by growth confined to the stab line. Equivocal motility tests were confirmed by phase-contrast micros-copy of wet mounts from overnight stationary broth cultures or from the growth edge of young colonies incubated at 30°C for less than 24 h. Isolates that failed to demonstrate motility in more than 25 fields were retested by phase-contrast microscopy of log-phase cells grown in 4 ml of Trypticase soy broth (REMEL) in 50-ml conical tubes to facilitate aeration by shaking at 125 rpm for 2 to 3 h at 30°C. Coverslips on the wet mounts were not sealed in order to foster aeration at the edges. Each wet mount was observed immediately after preparation. Positive and negative motility controls were used with each set of tests:B. cereus
ATCC 14579 (motile),B. megateriumATCC 14581 (motile only from log-phase broth),B. thuringiensisATCC 35646 (motile),E. faecalisATCC 29212 (nonmo-tile), andE. coliATCC 25922 (motile).
Selective/differential media.The bacteria’s ability to grow in the presence of low concentrations of polymyxin B (10g/ml), ferment mannitol, and produce
lecithinase were evaluated on MEP agar (REMEL) after growth for 24 to 48 h at 30°C. Originally designed for the isolation ofB. cereusfrom food, MEP is a selective and differential medium that also selects for otherB. cereus group members, includingB. anthracis. A positive mannitol fermentation reaction pro-duces a yellow colony, and lecithinase production results in an opaque precipi-tate in the medium under and/or around a colony (10).B. anthraciscolonies are colorless and, having weak lecithinase production, have an opaque zone usually limited to beneath the colony (4, 6, 7, 12). Control strains wereB. cereusATCC 14579 andB. thuringiensisATCC 35646 (growth and lecithinase [⫹], mannitol fermentation [⫺]),B. anthracisPasteur CDC BC 3132 (growth [⫹], lecithinase weak [⫹], mannitol fermentation [⫺]), andB. megateriumATCC 14581 and“B. maroccanus” ATCC 25099 (no growth). Isolates were subsequently tested on a second selective and differential medium, ChrA (R&F Laboratories, Downers Grove, IL), at 35°C for 24 to 48 h. With this medium, the isolates were examined for growth in the presence of polymyxin B (12g/ml) and hydrolysis of 5-bromo-4-chloro-3-indoxyl-cholinephosphate by phosphatidylcholine-specific phospho-lipase C activity, also known as lecithinase C, the key bacterial enzyme in the hydrolysis of lecithin (10). Colonies that were positive for phospholipase C enzyme were teal blue with a cream rim at 24 to 48 h. According to the manu-facturer’s technical instructions, the width of the cream rim varied with the species (thick rim forB. anthracisand thin rim forB. cereusandB. thuringiensis). Colonies that were negative for phospholipase C were cream-colored through-out. Control strains wereB. anthracisPasteur CDC BC 3132 (growth and weak phospholipase C activity [⫹]);B. cereusATCC 14579,B. thuringiensisATCC 35646, andB. mycoidesATCC 6462 (growth and phospholipase C activity [⫹]);
“B. maroccanus” ATCC 25099 andB. megateriumATCC 14581 (no growth); and
B. circulansATCC 4513 andB. subtilisNRRL NRS 744 (USDA Agricultural Research Service Culture Collection) (growth [⫹] and phospholipase C activity [⫺]).
RESULTS
On media containing sheep blood, 5 of the 60 isolates (iso-lates 105, 208, 210, 504, 505) demonstrated the presence of hemolysins that produced a clear beta-hemolysis pattern (Ta-ble 1). An additional isolate (isolate 497) produced a greening (alpha-) hemolysis pattern. Gram stains revealed that two of the beta-hemolytic isolates (isolates 504 and 505) were gram-positive cocci in grape-like clusters, one nonhemolytic isolate was a gram-positive, branched nocardia-form rod (isolate 1-0926) and a second nonhemolytic isolate was a gram-nega-tive rod (isolate 2-0349). These four isolates (isolates 504, 505, 1-0926, and 2-0349) were taken out of the study. The remain-ing 56 isolates were all endospore-formremain-ing, catalase-positive, gram-positive rods. We included the isolates that displayed both alpha- and beta-hemolysis because they may not have appeared as such to the original laboratory worker who sent them to the reference laboratory. They would also act as con-trols in the testing of our methods.
Of the 56 isolates in the study, only five (8.9%) (isolates 105, 208, 210, 304, and 500) displayed motility in all of the motility
media, at both 30 and 35°C (Table 1). These were three B.
cereus, oneB. amyloliquefaciens, and oneBacillus isolate. Of the remaining 51 isolates, 9 (16.1%) demonstrated motility at only one temperature or in one type of the motility medium. Wet mounts of liquid broth cultures revealed a total of 34 (60.7%) additional isolates to be truly motile although three of these isolates did not show motility until the strains were grown in shaking broth and observed at early log phase growth. Only eight isolates (14.3%) (isolates 209, 247, 261, 264, 303, 499, 502, and 506) were nonmotile by all motility tests.
There was no distinct advantage for the presence or absence of the dye TTC in the motility media. TTC appeared to aid in the interpretation of motility for only three isolates (isolates 300, 497, and 498) and failed to resolve questionable motility for others. For five isolates (isolates 245, 246, 249, 263, and
on May 15, 2020 by guest
http://jcm.asm.org/
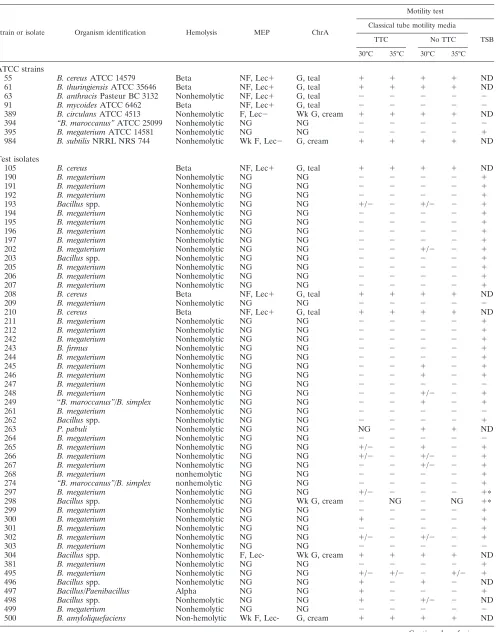
TABLE 1. Hemolysis pattern, growth on selective media, and motility tests of eight type strainBacillusspecies and 60 isolates received from sentinel laboratoriesa
Strain or isolate Organism identification Hemolysis MEP ChrA
Motility test
Classical tube motility media
TSB
TTC No TTC
30°C 35°C 30°C 35°C
ATCC strains
55 B. cereusATCC 14579 Beta NF, Lec⫹ G, teal ⫹ ⫹ ⫹ ⫹ ND
61 B. thuringiensisATCC 35646 Beta NF, Lec⫹ G, teal ⫹ ⫹ ⫹ ⫹ ND
63 B. anthracisPasteur BC 3132 Nonhemolytic NF, Lec⫹ G, teal ⫺ ⫺ ⫺ ⫺ ⫺
91 B. mycoidesATCC 6462 Beta NF, Lec⫹ G, teal ⫺ ⫺ ⫺ ⫺ ⫺
389 B. circulansATCC 4513 Nonhemolytic F, Lec⫺ Wk G, cream ⫹ ⫹ ⫹ ⫹ ND
394 “B. maroccanus”ATCC 25099 Nonhemolytic NG NG ⫺ ⫺ ⫺ ⫺ ⫺
395 B. megateriumATCC 14581 Nonhemolytic NG NG ⫺ ⫺ ⫺ ⫺ ⫹
984 B. subtilisNRRL NRS 744 Nonhemolytic Wk F, Lec⫺ G, cream ⫹ ⫹ ⫹ ⫹ ND
Test isolates
105 B. cereus Beta NF, Lec⫹ G, teal ⫹ ⫹ ⫹ ⫹ ND
190 B. megaterium Nonhemolytic NG NG ⫺ ⫺ ⫺ ⫺ ⫹
191 B. megaterium Nonhemolytic NG NG ⫺ ⫺ ⫺ ⫺ ⫹
192 B. megaterium Nonhemolytic NG NG ⫺ ⫺ ⫺ ⫺ ⫹
193 Bacillusspp. Nonhemolytic NG NG ⫹/⫺ ⫺ ⫹/⫺ ⫺ ⫹
194 B. megaterium Nonhemolytic NG NG ⫺ ⫺ ⫺ ⫺ ⫹
195 B. megaterium Nonhemolytic NG NG ⫺ ⫺ ⫺ ⫺ ⫹
196 B. megaterium Nonhemolytic NG NG ⫺ ⫺ ⫺ ⫺ ⫹
197 B. megaterium Nonhemolytic NG NG ⫺ ⫺ ⫺ ⫺ ⫹
202 B. megaterium Nonhemolytic NG NG ⫺ ⫺ ⫹/⫺ ⫺ ⫹
203 Bacillusspp. Nonhemolytic NG NG ⫺ ⫺ ⫺ ⫺ ⫹
205 B. megaterium Nonhemolytic NG NG ⫺ ⫺ ⫺ ⫺ ⫹
206 B. megaterium Nonhemolytic NG NG ⫺ ⫺ ⫺ ⫺ ⫹
207 B. megaterium Nonhemolytic NG NG ⫺ ⫺ ⫺ ⫺ ⫹
208 B. cereus Beta NF, Lec⫹ G, teal ⫹ ⫹ ⫹ ⫹ ND
209 B. megaterium Nonhemolytic NG NG ⫺ ⫺ ⫺ ⫺ ⫺
210 B. cereus Beta NF, Lec⫹ G, teal ⫹ ⫹ ⫹ ⫹ ND
211 B. megaterium Nonhemolytic NG NG ⫺ ⫺ ⫺ ⫺ ⫹
212 B. megaterium Nonhemolytic NG NG ⫺ ⫺ ⫺ ⫺ ⫹
242 B. megaterium Nonhemolytic NG NG ⫺ ⫺ ⫺ ⫺ ⫹
243 B. firmus Nonhemolytic NG NG ⫺ ⫺ ⫺ ⫺ ⫹
244 B. megaterium Nonhemolytic NG NG ⫺ ⫺ ⫺ ⫺ ⫹
245 B. megaterium Nonhemolytic NG NG ⫺ ⫺ ⫹ ⫺ ⫹
246 B. megaterium Nonhemolytic NG NG ⫺ ⫺ ⫹ ⫺ ⫹
247 B. megaterium Nonhemolytic NG NG ⫺ ⫺ ⫺ ⫺ ⫺
248 B. megaterium Nonhemolytic NG NG ⫺ ⫺ ⫹/⫺ ⫺ ⫹
249 “B. maroccanus”/B. simplex Nonhemolytic NG NG ⫺ ⫺ ⫹ ⫺ ⫹
261 B. megaterium Nonhemolytic NG NG ⫺ ⫺ ⫺ ⫺ ⫺
262 Bacillusspp. Nonhemolytic NG NG ⫺ ⫺ ⫺ ⫺ ⫹
263 P. pabuli Nonhemolytic NG NG NG ⫺ ⫹ ⫹ ND
264 B. megaterium Nonhemolytic NG NG ⫺ ⫺ ⫺ ⫺ ⫺
265 B. megaterium Nonhemolytic NG NG ⫹/⫺ ⫺ ⫹ ⫺ ⫹
266 B. megaterium Nonhemolytic NG NG ⫹/⫺ ⫺ ⫹/⫺ ⫺ ⫹
267 B. megaterium Nonhemolytic NG NG ⫺ ⫺ ⫹/⫺ ⫺ ⫹
268 B. megaterium nonhemolytic NG NG ⫺ ⫺ ⫺ ⫺ ⫹
274 “B. maroccanus”/B. simplex nonhemolytic NG NG ⫺ ⫺ ⫺ ⫺ ⫹
297 B. megaterium Nonhemolytic NG NG ⫹/⫺ ⫺ ⫺ ⫺ ⫹ⴱ
298 Bacillusspp. Nonhemolytic NG Wk G, cream ⫺ NG ⫺ NG ⫹ⴱ
299 B. megaterium Nonhemolytic NG NG ⫺ ⫺ ⫺ ⫺ ⫹
300 B. megaterium Nonhemolytic NG NG ⫹ ⫺ ⫺ ⫺ ⫹
301 B. megaterium Nonhemolytic NG NG ⫺ ⫺ ⫺ ⫺ ⫹
302 B. megaterium Nonhemolytic NG NG ⫹/⫺ ⫺ ⫹/⫺ ⫺ ⫹
303 B. megaterium Nonhemolytic NG NG ⫺ ⫺ ⫺ ⫺ ⫺
304 Bacillusspp. Nonhemolytic F, Lec- Wk G, cream ⫹ ⫹ ⫹ ⫹ ND
381 B. megaterium Nonhemolytic NG NG ⫺ ⫺ ⫺ ⫺ ⫹
495 B. megaterium Nonhemolytic NG NG ⫹/⫺ ⫹/⫺ ⫺ ⫹/⫺ ⫹
496 Bacillusspp. Nonhemolytic NG NG ⫹ ⫺ ⫹ ⫺ ND
497 Bacillus/Paenibacillus Alpha NG NG ⫹ ⫺ ⫺ ⫺ ⫹
498 Bacillusspp. Nonhemolytic NG NG ⫹ ⫺ ⫹/⫺ ⫺ ND
499 B. megaterium Nonhemolytic NG NG ⫺ ⫺ ⫺ ⫺ ⫺
500 B. amyloliquefaciens Non-hemolytic Wk F, Lec- G, cream ⫹ ⫹ ⫹ ⫹ ND
Continued on facing page
on May 15, 2020 by guest
http://jcm.asm.org/
265), the media without TTC demonstrated clearer motility than when TTC was present. However, the incubation temper-ature for the motility tests did seem to be important. As seen in Table 1, eight isolates (isolates 245, 246, 249, 263, 265, 300, 496, 497, and 498) that were nonmotile at 35°C expressed strong motility at 30°C.
The tube motility tests gave inaccurate motility results most of the time for most of the genera and species tested (Table 2).
One presumptiveB. cereus(isolate 503) was nonmotile in all of
the tube motility tests, and yet it displayed strong motility when grown in the broth media and visualized microscopically
(Ta-bles 1 and 2). Only 7 of the 39 isolates identified asB.
mega-teriumwere nonmotile by both the tube tests and the micro-scopic observation of broth suspensions.
When all 56 isolates were inoculated onto MEP and ChrA, 50 isolates did not grow on MEP and 49 did not grow on ChrA. One isolate (298) grew on ChrA but not on MEP (Table 1). It is important to note that on the ChrA media, 42 of the isolates
produced very few (ⱕ5) small (ⱕ2 mm) colonies at the initial
inoculation site after 24 to 48 h. No colonies were seen on the streaks in other quadrants of the media plates. These were
considered no growth and therefore not belonging to the B.
cereusgroup. Of the six isolates that grew on MEP, four iso-lates (isoiso-lates 105, 208, 210, and 503) did not ferment mannitol
and produced lecithinase and were identified as part of theB.
cereusgroup. Three of these (isolates 105, 208, and 210) were the beta-hemolytic strains that displayed strong motility and
were identified asB. cereus. The fourth isolate (isolate 503) was
nonhemolytic but motile only when examined after growing in
broth media and was identified as a presumptiveB. cereus. Two
isolates (isolates 304 and 500) fermented mannitol and did not produce lecithinase.
Of the seven isolates that grew on ChrA, three B. cereus
(isolates 105, 208, and 210) and one presumptive B. cereus
(isolate 503) grew luxuriantly as teal blue colonies (5 to 10 mm) with a cream rim at the edge after 24 h and with more intense teal blue at 48 h. The other three isolates (isolates 298, 304, and 500) grew more slowly, producing smaller (0.5 to 3 mm) cream-colored colonies at 24 and 48 h. These were identified as
B. amyloliquefaciensand Bacillus species not of theB. cereus
group (Table 1).
Further biochemical tests showed that 39 (69.6%) of the 56
isolates were identified asB. megaterium, while the remaining
17 (30.4%) were as follows: oneB. amyloliquefaciens, threeB.
[image:4.585.45.542.79.226.2]cereus, one presumptiveB. cereus, oneB. firmus, two “B. ma-roccanus”/B. simplex(5), onePaenibacillus pabuli, one Paeni-bacillus/Bacillusgroup, and sevenBacillusspp., not of the B. cereus group. Of these, the four B. cereus group, one B.
TABLE 1—Continued
Strain or isolate Organism identification Hemolysis MEP ChrA
Motility test
Classical tube motility media
TSB
TTC No TTC
30°C 35°C 30°C 35°C
501 B. megaterium Nonhemolytic NG NG ⫺ ⫺ ⫺ ⫺ ⫹ⴱ
502 B. megaterium Nonhemolytic NG NG ⫺ ⫺ ⫺ ⫺ ⫺
503 B. cereuspresumptive Nonhemolytic NF, Lec⫹ G, teal ⫺ ⫺ ⫺ ⫺ ⫹
506 B. megaterium Nonhemolytic NG NG ⫺ ⫺ ⫺ ⫺ ⫺
507 B. megaterium Nonhemolytic NG NG ⫺ ⫺ ⫺ ⫺ ⫹
1-0946 Nocardia-like rod Nonhemolytic ND ND ND ND ND ND ND
2-0349 Gram-negative rod Nonhemolytic ND ND ND ND ND ND ND
504 Gram-positive coccus Beta ND ND ND ND ND ND ND
505 Gram-positive coccus Beta ND ND ND ND ND ND ND
a
The hemolysis column shows the hemolytic pattern seen on Trypticase soy agar supplemented with 5% sheep red blood cells.⫹, Positive reaction;⫺, negative reaction; NF, nonfermenter of mannitol; F, fermenter; Wk F, weak fermenter; Lec, lecithinase; teal, phospholipase C activity; cream, no phospholipase C; Wk G, weak growth; G, growth; NG, no growth; ND, not done.ⴱ, This isolate had positive motility only after examination of early-log-phase growth in broth aerated with shaking.
TABLE 2. Analysis of growth and motility of 56 cultures of gram-positive rods on various media
Strain n
No. (%) of isolates with growth:
No. (%) of isolates positive for motilitya:
With TTCb
Without TTC
On MEP On ChrA 30°C 35°C 30°C 35°C
B. amyloliquefaciens 1 1 (100) 1 (100) 1 (100) 1 (100) 1 (100) 1 (100)
B. cereus 4 4 (100) 4 (100) 3 (75) 3 (75) 3 (75) 3 (75)
B. firmus 1 0 (0) 0 (0) 0 (0) 0 (0) 0 (0) 0 (0)
B. megaterium 39 0 (0) 0 (0) 1 (2.6) 0 (0) 3 (7.8) 0 (0)
“B. maroccanus”/B. simplex 2 0 (0) 0 (0) 0 (0) 0 (0) 1 (50) 0 (0)
Bacillusspp. (notB. cereusgroup) 7 1 (14.3) 2 (28.6) 3 (42) 1 (14) 2 (28) 1 (14)
Paenibacillus pabuli 1 0 (0) 0 (0) 0 (0) 0 (0) 1 (100) 1 (100)
Paenibacillus/Bacillusspp. 1 0 (0) 0 (0) 1 (100) 0 (0) 0 (0) 0 (0)
Total 56 6 (10.7) 7 (12.5) 9 (16) 5 (9) 11 (19.6) 6 (11)
aResults for microscopic examination of wet mounts from broth cultures are not shown.
on May 15, 2020 by guest
http://jcm.asm.org/
[image:4.585.43.543.575.716.2]amyloliquefaciens, and one Bacillus sp. (notB. cereus group) isolates grew on both MEP and ChrA. Subsequent
identifica-tion of an isolate as belonging to theB. cereusgroup or not
correlated well with their respective growth or lack of growth on the media and the ensuing reactions when growth did occur.
DISCUSSION
Of 60 isolates the FDOH Tampa LRN reference laboratory received from sentinel laboratories, only 52 needed further
examination by FDOH personnel to rule outB. anthracis.
The LRN protocols for the reference laboratory include Gram stain, gamma phage susceptibility, DFA for cell wall and capsule production, and molecular PCR assays for DNA
tar-gets specific forB. anthracis(2, 3, 13). Following this scheme,
the Gram stain screened out the two gram-positive cocci, the nocardia-form rod, and the gram-negative rod and precluded any further work on these isolates. The beta- and alpha-hemo-lytic patterns observed on BA plates for four additional isolates prevented these four from being further tested by the LRN Reference laboratory, although we retained these latter four in our study as controls for the two media and motility tests. These four may have appeared nonhemolytic and nonmotile to the originating laboratory. The LRN tests (gamma phage sus-ceptibility, DFA, and PCR) were performed on the remaining
52 isolates to rule outB. anthracisprior to our receiving them
from FDOH and including them in our study.
Laboratory error in the sentinel laboratories can explain the fact that eight nonbacillus or nonhemolytic isolates were in-correctly sent to the LRN reference laboratory. The addition of a selective and differential media such as MEP or ChrA media may have reduced the number of isolates that would need to be sent to the reference lab. Used in the reference lab, these media would be an aid in identifying or ruling out truly
suspicious colonies. All fourB. cereusgroup isolates grew on
the two media and displayed the characteristic reactions of opaqueness (lecithinase positive) and lack of color (mannitol fermentation negative) on MEP or a dark teal color with a cream rim (phospholipase C activity) on ChrA (Tables 1 and 2). Although the manufacturer of ChrA states that one can
distinguish the species of the B. cereus group based on the
intensity of the teal blue pigment and the width of the cream-colored colony margin, we could not comfortably differentiate
amongB. anthracisPasteur CDC BC 3132,B. mycoidesATCC
6462, andB. thuringiensisATCC 35646 on the media. Before
any biochemical identification was attempted, oneB.
amyloliq-uefaciens and two Bacillus spp. (not B. cereus group) were
identified as not beingB. anthracisbased upon their
appear-ance on the media (observed mannitol fermentation, no
leci-thinase reaction, and smaller colonies). Significantly, all 39B.
megaterium(the most common nonpathogenic species
submit-ted to the laboratory) and two “B. maroccanus”/B. simplex
isolates either failed to thrive or did not grow on both media. Although no growth was the norm for the isolates on MEP,
strains ofB. megateriumand “B. maroccanus”/B. simplex
grow-ing on ChrA frequently produced one to five colonies of lim-ited size (ca. 2 mm at 24 h) restricted to the inoculation site.
The constrainedB. megateriumcolonies produced a cream or,
infrequently, a light teal green color, whereas those presumed
to be “B. maroccanus”/B. simplexproduced a dark teal blue
colony, a finding consistent with lecithinase production by the species (12). It is doubtful that the severe restriction of growth
for this species on ChrA would result in “B. maroccanus”/B.
simplexbeing mistaken forB. cereusgroup provided the man-ufacturer’s instructions are closely followed. Misleading results could potentially occur if a heavy inoculum was used to make a single short streak, or if the plates were used beyond the manufacturer’s suggested shelf-life (60 days at 4°C). The MEP and ChrA media can be read and interpreted at 24 and 48 h and could easily fit into the LRN protocol either in the sentinel or reference labs. Both media are more expensive than normal TSA or blood agar plates, but they are much less expensive than antibody-based and molecular tests such as DFA and PCR. As revealed in Tables 1 and 2, the tube motility tests are very difficult to interpret even in our hands. TTC is incorporated into motility medium to minimize errors in discerning growth-driven turbidity. The tetrazolium salt is colorless, but as the cells grow, the dye is incorporated into the organism and is reduced to an insoluble red pigment. Most of the isolates appeared to be nonmotile regardless of whether TTC dye was included in the medium or whether the tests were incubated at 30 or 35°C. In data not presented, the size of the inoculum as well as the diameter and type of needle used affected the appearance of motility and generated both false and inconsis-tent interpretations. Inoculation with a narrow gauge
(⬍0.5-mm diameter) wire needle facilitated the introduction of
a single, discrete stab line, as well as a consistently reduced inoculum size. Stabs made with disposable, broad-gauge
(⬎1.0-mm diameter) plastic needles delivered larger and less
consistently sized inocula that frequently resulted in significant secondary growth at the interface between the wall of the tube and the surface of the column of medium as well as the occur-rence of billowing lobes of growth from the stab lines. After 48 h of incubation, the confluence of dense turbidity at the interface with the presence of apparent growth extending away from otherwise discrete stab lines made interpretation prob-lematic because it appeared to be oxygen-limited, arrested
motility. Because many Bacillus are aerobic, growth occurs
mainly at the top of the media and can be very difficult to discern deeper along the stab line. This pattern can be falsely interpreted as a negative motility. The failure to thrive and the failure to demonstrate motility in reduced oxygen tension are
characteristics of B. megaterium. These two characteristics
most likely contribute to the number of isolates of this species being referred to the LRN reference laboratory. In addition,
other species ofBacillusmay not grow in the motility media,
causing needless worry or confusion for personnel.
Wet mounts of cultures grown in aerated broth media, es-pecially when examined during the early log phase of growth, allowed one to easily see motility. During early log phase, motility was easily observed when the cells were single or in chains of only two or three. In contrast, for later-log-phase bacilli, the motility was obscured and infrequently observed or
sluggish, especially for B. megaterium strains that produced
moderate to lengthy chains. Because we could observe motility in three isolates only in the early log phase and not later, we switched to performing the wet mount exclusively from aerated broth cultures in the first 2 to 3 h of growth. In our laboratory, the coverslips were not sealed, and the areas close to the edges of the slips (areas with expected greater oxygen tension) were
on May 15, 2020 by guest
http://jcm.asm.org/
observed for motile cells. We feel that these tests were the equivalent of using the hanging drop method favored by some laboratories.
Of the 60 isolates that the FDOH laboratory originally re-ceived and shared with us, none of the isolates would have warranted further testing by the FDOH personnel if both the modified motility test and polymyxin-containing media had been incorporated in the LRN protocol. With the sole addition of the modified motility tests, only eight (13%) of these isolates were truly nonmotile (isolates 209, 247, 261, 264, 303, 499, 502, and 506). Using only the MEP or ChrA media, only one isolate (isolate 503) would have needed further testing by the refer-ence lab since the others that grew on the media were either beta-hemolytic on blood plates (isolates 105, 208, and 210) or demonstrated mannitol fermentation and negative lecithinase activity (isolates 298, 304, and 500).
If a reference laboratory used both the motility and one of the media (i.e., MEP), the MEP plate could be streaked the same day as blood plates are currently done. Both plates would be read at 24 h, and if there was no growth on the MEP plate or growth of a lecithinase-negative mannitol fermenting
col-ony, no further tests would be needed to rule outB. anthracis.
If the MEP plates grew nonfermenting and lecithinase-positive colonies, the modified motility test could then be set up and read the same day. A positive motility would preclude the continuation of tests (DFA, PCR, etc.).
When the sentinel laboratories first recognize the presence
of an isolate that looks like a potentialB. anthracis, they test
for beta-hemolysis and motility. If the modified motility test was set up immediately, it could be viewed microscopically the same day. If the motility was negative, the special medium plate (i.e., MEP) could be streaked, incubated overnight, and examined for growth the next day, along with the media used for transport (usually a blood plate) to the LRN reference lab. No growth on the MEP plate would show that the suspicious
colony was notB. anthracisand would not need to be sent to
the reference laboratory. Only growth that indicated that the isolate was nonfermenting and lecithinase positive would then be sent to the reference laboratory.
Based on our findings, we suggest that LRN reference lab-oratories consider adding either MEP or ChrA and the micro-scopic observation of motility, as described above, to the LRN protocol. The reference laboratories would benefit since they would have fewer specimens that would need confirmation by the more expensive confirmatory tests. The LRN confirmatory tests are expensive and can demand much hands-on technical time. When only one or two isolates are received, this may not be a problem. However, when many isolates are received, or when the laboratory has a high workload of other demanding responsibilities, time and expense do become important issues. Sentinel laboratories may realize cost savings if the LRN would recommend the modified motility test of log-phase growth and the use of one of these two selective media as supplements to
the current protocol for “ruling out”B. anthracis. There would
potentially be fewer suspicious isolates that would need to be sent to the reference laboratories, thus reducing transportation costs. We did not compute all possible costs or savings, and this probably could best be addressed in a full analysis by the LRN, although we did note that the MEP plates cost half as much as the ChrA plates.
The sentinel laboratories should be able to send any and all suspicious or questionable isolates to the LRN reference lab-oratories. Yet it is important that the system and personnel at the reference labs not be overburdened with samples that do
not qualify for confirmatory testing as B. anthracis. This is
especially important during a perceived or real threat because the reference laboratories are also responsible for testing all environmental samples. Experience tells us that the LRN ref-erence laboratories will have an increase in all types of speci-mens submitted to them when an event occurs. Therefore, the addition of either MEP or ChrA and the described motility test could be very beneficial to the whole LRN system.
ACKNOWLEDGMENT
This study was supported by Department of Defense grant DAAD-13-01-C-0043.
REFERENCES
1.Atlas, R. M.1993. NSMP, modified, p. 665–666.InL. C. Parks (ed.), Hand-book of microbiological media. CRC Press, Inc., Boca Raton, Fla. 2.Centers for Disease Control and Prevention.2001. Update: investigation of
anthrax associated with intentional exposure and interim public health guidelines, October 2001. Morbid. Mortal. Wkly. Rep.50:889–893. 3.Centers for Disease Control and Prevention.2001. Investigation of
bioter-rorism-related anthrax and interim guidelines for exposure management and antimicrobial therapy, October 2001. Morb. Mortal. Wkly. Rep.50:909–919. 4.Gibson, T., and R. E. Gordon.1974. Endospore-forming rods and cocci, p. 529–575.InR. E. Buchanan and N. E. Gibbons (ed.), Bergey’s manual of determinative bacteriology, 8th ed. The Williams & Wilkins Company, Bal-timore, Md.
5.Heyrman, J., N. A. Logan, M. Rodriquez-Diaz, P. Scheldeman, L. Lebbe, J. Swings, M. Heyndricks, and P. de Vos.2005. Study of mural painting isolates, leading to the transfer of “Bacillus maroccanus” and “Bacillus carotarum” to
Bacillus simplex, re-examination of the strains previously attributed to “ Ba-cillus macroides” and description ofBacillus muralissp. nov. Int. J. Syst. Evol. Microbiol.55:119–131.
6.Logan, N. A., and P. C. B. Turnbull.1999.Bacillusand recently derived genera, p. 357–369. InP. R. Murray, E. J. Baron, M. A. Pfaller, F. C. Tenover, and R. H. Yolken (ed.), Manual of clinical microbiology, 7th ed. American Society for Microbiology, Washington, D.C.
7.Logan, N. A., and P. C. B. Turnbull.2003.Bacillus and other aerobic endospore-forming bacteria, p. 445–460.InP. R. Murray, E. J. Baron, J. H. Jorgensen, M. A. Pfaller, and R. H. Yolken (ed.), Manual of clinical micro-biology, 8th ed. American Society for Micromicro-biology, Washington, D.C. 8.Jernigan, J. A., D. S. Stephens, D. A. Ashford, C. Omenaca, M. S. Topiel, M.
Galbraith, M. Tapper, T. L. Fisk, S. Zaki, T. Popovic, R. F. Meyer, C. P. Quinn, S. A. Harper, S. K., Fridkin, J. J. Sejvar, C. W. Shepard, M. Mc-Connell, J. Guarner, W. Shieh, J. M. Malecki, J. L. Gerberding, J. M. Hughes, B. Perkins, et al.2001. Bioterrorism-related inhalational anthrax: the first 10 cases reported in the United States. Emerg. Infect. Dis.7:933–944. 9.Jernigan, J. A., P. L. Raghunathan, B. P. Bell, R. B. Bresnitz, J. C. Butler, M. Cetron, M. Cohen, R. Doyle, C. Greene, K. S. Griffith, J. Guarner, J. Hadler, J. A. Hayslett, R. Meyer, L. R. Petersen, M. Phillips, R. Pinner, T. Popovic, C. P. Quinn, J. Reefhuis, D. Reissman, N. Rosenstein, A. Schuchat, W. Shieh, L. Siegal, D. L. Swerdlow, F. C. Tenover, M. Traeger, J. W. Ward, I. Weisfuse, S. Wiersma, K. Yeskey, S. Zaki, D. A. Ashford, B. A. Perkins, S. Ostroff, J. Hughes, D. Fleming, J. P. Koplan, J. L. Gerberding, et al.2002. Investigation of bioterrorism-related anthrax, United States, 2001: epidemi-ologic findings. Emerg. Infect. Dis.8:1019–1028.
10.MacFaddin, J. F.2000. Lecithinase test, p. 273–281.InL. McGrew (ed.), Biochemical tests for identification of medical bacteria, 3rd ed. Lippincott/ The Williams & Wilkins, Philadelphia, Pa.
11.Smirnoff, W. A.1962. A staining method for differentiating spores, crystals, and cells ofBacillus thuringiensis(Berliner). J. Insect Pathol.4:384–386. 12.Sneath, P. H. A.1986. Endospore-forming gram-positive rods and cocci, p.
1104–1139.InP. H. A. Sneath, N. S. Mair, M. E. Sharpe, and J. G. Holt (ed.), Bergey’s manual of systematic bacteriology, vol. 2. The Williams & Wilkins Company, Baltimore, Md.
13.Weyant, R. S., J. W. Ezzell, and T. Popovic.2003. Level B. Laboratory procedure for the identification ofBacillus anthracis, p. 47–64.InK. Q. Lindsey and S. A. Morse (ed.), Laboratory protocols for bioterrorism re-sponse laboratories for the identification ofBacillus anthracis. [Online.] www.lrnb.cdc.gov.