GENETIC ANALYSIS
OF
THE DEVELOPMENTAL PROCESSES DURING
GERMINATION AND OUTGROWTH OF BACZLLUS SUBTZLZS
SPORES WITH TEMPERATURE-SENSITIVE MUTANTS
JUN-ICHI NUKUSHINA AND YONOSUKE IKEDA
The Institute of Applied Microbiology, University of Tokyo, Tokyo, Japan
Received December 19, 1968
HE
developmental processes in bacteriophage
T4
have been studied exten-
Tsively with temperature-sensitive mutants and amber mutants
(EDGAR,
DEN-
HARDT
and
EPSTEIN
1964; EDGAR
and
LIELAUSIS
1964).
In
Bacillus subtilis,
the
sporulation process
has
been studied with asporogenous mutants
(
RYTER,
SCHAEF-
FER
and
IONESCO
1966; TAKAHASHI
1965).
We examined the developmental processes involved
in the transition from
bacterial spores to vegetative cells with temperature-sensitive
( t s )mutants
o f
B. subtilis.
When incubated at a high temperature
(48"C),
the mutant spores
discontinued their development at various stages of outgrowth. The mutant loci
were mapped on the chromosome
by
transformation and transduction techniques.
The results will be discussed from the aspect of sequential expression
of
genetic
information during the course of germination and outgrowth
of
spores.
MATERIALS A N D METHODS
Strains: Mutants were derived from a tryptophan-requiring strain (160 t r y ) of Bacillus subtilis Marburg. The strain grows normally and forms colonies at 50°C. Other strains used for mapping are shown in Table 1. Phage PBSl was supplied by Dr.
I.
TAKAHASHI of McMaster University, Hamilton, Ont., Canada.Media: Mutants were isolated on NB agar medium (0.8% Difco Nutrient Broth and 1.5% Difco Bacto-agar)
.
Spores were germinated in MW (modified WOESE'S) medium. The medium contained, in 1,000 ml distilled water, glucose, 9 g; Na L-glutamate, 3 g; L-asparagine, 0.3 g; L-alanine, 100 mg; L-arginine.
HC1, 100 mg; casamino acids (Difco), 0.5 g; NH,Cl, 0.5 g; KCI, 70 mg; Na,SO,, 120 mg; MgC1,. 6H,O, 0.5 g; Ca acetate, 20 mg; MnC1,. 4H,O, 5 mg; NaH,PO, 2H,O, 156 mg; L-tryptophan, 30 mg; and tris(hydroxmethyl)aminomethane, 3 g (pH 7.2). Spores were prepared on potato extract agar slants as described by SPIZIZEN (1958).Dilution buffer used for the preparation of phage PBSl suspension consisted of NaCI, 5 g; MgC1, .6H,O, 0.2 g; CaCI,, 0.1 g; tris(hydroxymethyl)aminom&ane, 1.2 g (pH 7.4); and 1,000 ml distilled water.
Chemicals. 2-aminopurine and 5-bromodeoxyuridine were purchased from the California Biochem. Corp. and Sigma Chem. Co., respectively.
Straining: The Wirtz staining method (SCHAEFFER and FULTON 1933) using 5% malachite
green and 0.5% safranin was employed. Dormant spores were stained green and germinated spores red.
Isohtion of ts mutants: Spores of strain 160 try suspended in sterile 0.85% NaCl solution were exposed to gamma rays (241) kr) from a 6"Co source o r ultraviolet light (UV) (12,000 to 15,000 erg/",) from a 20-W germicidal lamp. Vegetative cells were cultured in minimal medium containing 2-aminopurine. (1 mg/ml) or 5-bromodeoxyuridine (0.4 mg/ml) overnight at 37°C.
64.
JUN-ICHI NUKUSHINA AND YONOSUKE IKEDATABLE 1
List of strains
Strain W23 sfr 23 ade met
160
101 Mu8u5u6 Mu8u5u5 flat A26 fla+ SP25 BC9 6160 M26
35
Genotypic designation 23 str-r
168 sul-r
23 ade met
160 try-3
168 purB6 leu-101 metB5
168 thrd leu-8 metB5 mot
168 ura-26 mot
160 argA15 phoP17
160 phoR9 try-3
168 purB6 leu-101 try-3 metB5
168 leu-8 ura-26 lys-21 metB5
160 argA15 hisB31
Source N. SUEOKA S. ZAMENHOF
T.
IIJIMA
P.
R.
BURKHOLDER and N.H.
GILES N. SUEOKAJ. MARMUR
J. MARMUR
T. MIKI
T. MIKI
J. NUKUSHINA
J.
NUKUSHINA
N. MUNAKATA
Abbreviations used: for nutritional requirements: pur, adenine, or guanine requirement; thr,
threonine; try, tryptophan; met, methionine; his, histidine; for resistance markers: str, strepto- mycin; sul, sulfonamide; mot, motility; phoP, and phoR, structural and regulatory genes, respectively, of alkaline phosphatase; fla
+
,
flagellated.After the exposure or treatment, survivors were plated on NB plates. Among the replica-plated colonies, those which failed to grow a t 48°C but grew a t 37°C were selected as ts mutants. Yields were about 0.5%, O.l%, and O.Ol%, respectively, when treated with
UV,
gamma rays, and 2-aminopurine. Ninety-seven t s mutants were obtained.Transformation procedures: Transformation experiments were conducted as described by
TANOOKA and SAKAKIBARA (1968). When ts+ transformants were to be scored, a transformed
culture was plated on SPIZIZEN’S minimal medium ( SPIZIZEN 1958) supplemented with L-trypto- phan (50 pg/ml )and L-arginine (50 pg/ml), and the plates were incubated first a t 37°C for 4
hr and then a t 48°C for 44)
hr.
L-arginine was added to all the plates except when arg+ trans- formants were to be scored. This addition minimized the error due to unequal growth of trans- formants. The procedure of preincubation at 37°C for 4 h r was used to help the full expression of ts+ gene function (MCDONALD and MATNEY 1963).Measurement of marker frequencies: Genetic mapping was carried out by the marker fre- quency method originally described by YOSHIKAWA and SUEOKA (1963). As the DNA donor, strain W23, kindly supplied by Dr. N. SUEOKA of Princeton University, was used. The log phase DNA was prepared by the p H 9 phenol method of SAITO and MIURA (1963) from the cells of W23 grown to a cell concentration of 7
x
107 cells/ml in the standard medium. The standard medium consisted of the components of SPIZIZEN’S minimal medium, 0.05% Difco casamino acids, and 50 pg/ml of L-tryptophan. The stationary phase DNA was prepared from cells grown for 17 h r i n the same medium. The marker frequencies were calculated from the number of trans- formants appearing on four platesGERMINATION MUTANTS IN
B.
subtilis
65
in Penassay broth. Later procedures were the same as described above. About 4-6
x
1010 viable phages were prepared from one plate. PBSl was titrated on NB plates by mix-plating with spores of 23 a& met.Transduction procedures; Transduction experiments were carried out principally as described by TAKAHASHI (1961). When cotransduction of a t s marker or alkaline phosphatase marker and a n auxotrophic marker was to be studied, transductants for a n auxotrophic marker were trans- ferred with sterile toothpicks to fresh selective plates, and colonies appearing on the plates were replica-plated onto appropriate media to examine unselected markers. The t s character introduced into derivatives of B . subtilis 168 was checked on arginine-supplemented media, because the re- cipient strain required arginine for growth at 48°C. Derivatives of B . subtilis 160 showed no re- quirement for arginine at 48°C. Alkaline phosphatase markers were studied as described by MIKI, MINAMI, and IKEDA (1965). At least two single colonies were purified from each CO-
transductant and their characters were examined in detail.
RESULTS
Germination and outgrowth
of
t smutants at 48'C:
Figure
1
shows
optical
2.0
0
(0 (0
n
a,
1.5
0
>
a
a,
K
.-
-w-
1
.o
0.5
3
7°C- 48°C
P
1
2
3
Time (hr)
66
JUN-ICHI NUKUSHINA AND YONOSUKE IKEDAdensity curves of parental spores incubated in
MW
medium
at
37°C and
48°C.
I n one lot, the incubation temperature was raised from 37°C to 48°C after a n
incubation of 25 min. The spores incubated throughout at 37°C developed into
vegetative cells passing through the stages of germinated spores, swollen spores,
elongated cells, and septate cells. The optical density curve showed normal germi-
nation and outgrowth. The spores incubated throughout at
48°C
did not change
in their optical densities, but the spores which had been incubated first at 37°C
€or 25 min and then the temperature raised to 48°C showed normal germination
and outgrowth. Thus, the first step (germination) is temperature-sensitive in the
parental strain.
It
was for this reason that the procedure of preincubation at 37°C
for 25 min was adopted in later experiments. Figure
2 shows optical density curves
of spores
of
ts28,
ts57, tsl15, and the parental strain in
MW
medium. The mutant
spores decreased in their optical densities during the incubation at 37°C for
25
min and the low level was maintained for a long period during further incubation
at 48°C. Figure 5-A shows cell morphology of tsl15 harvested from a
4
hr
culture
and stained with malachite green and safranin. The morphology is that of germi-
2.0
0 (0 (0
n
$
1 . 5
0
.-
C I
m
a, E
-
I
1.
0.
0
5
0
1
2
3
4
T i m e
(hr)
0
1
2
3
4
T i m e
(hr)
GERMINATION MUTANTS IN
B.
subtilis
67
nated spores. Spores of
ts28
and
ts57
also gave the appearance
of
germinated
spores, and then developed into swollen spores very slowly. These mutants
will
be called “swellingts mutants” in the sense that the swelling process is temper-
ature sensitive.
A
group of
ts
mutants including
ts2, ts98, ts107,
and
tsl 1
1
failed to develop into
elongated cells. Figure
3
shows optical density curves of spores of these mutants
in M W or
NB
medium. Spores of
ts2
were sensitive to high temperature in
NB
medium, but not in
MW
medium. Further information about this mutant will
be
given later. Cell morphology of
tsl
1 1
harvested from a
4
hr
culture is shown
in
Figure
5-B.
Swollen spores, but not elongated cells, are observed. Mutants of
this
kind will be called “elongationts mutants”.
The third group of mutants involving
t s l , ts12, ts76, ts103, t s l l 0 , ts112,
and
t s l l 4
did not septate at 48°C. Optical density curves
of
spores of these mutants
in
M W medium are shown in Figure
4.
Optical densities increased to a considerably
high level and microscopic observation demonstrated that the cells were not able
0
U) U)
n
a,
1.5
0
>
(-3 a,
f
x
.-
c,-
1
.o
0.5
48°C
- I
fs
5 0
d
-
II
,fs
107,
1
2
3
4
Time (hr)
68
JUN-ICHI NUKUSHINA A N D YONOSUKE IKEDA2
.o
0
(0 W
n
Q)
1 . 5
0
>
a
a, E
.-
CI-
I
I
48°C
I
I
I
It s 1 1 4
t s
103
0
.
5
2
0
1
2
3
4
Time
(hr)
FIGURE
4.-Germination and outgrowth of septationts mutants. Spores were incubated as tlescribed in Figure 2.to
septate (Figure 5-C) . These mutants will be called “septationts mutants”. There
was another group of
ts
mutants which developed to the stage of dividing cells in
M W medium but did not
form any visible colony on NB plate (Figure
5-D).
Optical density curves of these mutant spores in M W medium resembled those
of the parental strain.
Among the
30 ts
mutants examined, 4 belonged to the swellingts, 9 to the elon-
gationts, and 10 to the septationtS group of mutants. The remaining 7
ts
mutants
belonged to the last group.
Vegetatiue
growth
of
ts
mutants
at
48’C:
About
3
x
10’ vegetative cells were
inoculated into
1
ml of NB or M W medium at 48°C and incubated throughout at
the same temperature. At intervals during the incubation, the cells were examined
under a microscope. Cell lysis was apparent in some of the elongationts mutants;
ts2
cells lysed when incubated in NB or M W medium supplemented with 0.1
%
FIGURE
5.-Photomicrographs
of
2s
cells
harwstc4
from
4
hr
culture
ilt
48°C.
The
scales
represent
10
p.
Ihnniuit
slmi.cs
in
the
photographs
are
the
spores
not
germinated
before
transfer
to
48°C.
A;
ts115
(swellingtS)
B;
tslll
(elongation's)
C;
is12
(septation'")
D;
70
JUN-ICHI NUKUSHINA A N D YONOSUKE IKEDATABLE 2
Marker frequency of try to m e a *
Log phase DNA Stationary phase DNA
Experiment - Marker
No. try+ meiB+ (try/metB) try+ meiB+ (try/metB) frequency 1 55300 54100 1.02 78600 76300 1.03 1
.oo
2 7910 7720 1.02 8120 7270 1.12 0.91
3 25400 24000 1.06 84000 65600 1.28 0.83
4 8400 7500 1.12 16400 1MOO 1.14 0.98
5 6230 5250 1.19 3560 3540 1.00 1.19
average 0.98 map position 1.02**
* Strains W23 and 6160
(purB6 leu-I01 try-3 metB5) were used, respectively, as the donor and recipient in these experiments. Transformability is expressed as the number of transfonn- ants/O.l ml.** Map position was calculated from the equation presented by
YOSHIKAWA and SUEOKA (1963).the synthesis of cell wall components might be temperature sensitive in these
mutants.
Mutants
of
the third
group formed filamentous cells even when started from
vegetative cells. The cells synthesized as much DNA and RNA as the parental
strain at
48’C,
suggesting that their defectiveness in septation is probably not due
to the secondary effect of inhibition of DNA synthesis.
Mapping
of
standard markers:
As all the
ts
mutants isolated carried the
try
marker in common, map position of
try
was studied first.
A
strain
(6160)
carry-
ing
markers
purB6,leu-101, try-3, metB5
was prepared from strain
101
(purB6
leu-101 metB5)
by transformation with DNA prepared from strain
160
try-3,
using the penicillin screening technique. Table
2
shows the marker frequency of
try
to
m t B .
This result suggests that the
try
locus
(1.02)
is located close to the
metB
(l.OO),
as has been reported by other workers (O’SULLIVAN
and SUEOKA
1967;
DUBNAU,
GOLDTHWAITE,
SMITH and MARMUR
1967).
Marker frequency
of
purB
to
m t B
was also studied (Table
3 ) .
From a mean value of
1.73,
the map
TABLE 3
Marker frequency of purB to metB*
Log phase DNA Experiment
No. purB+ meiBf (purB/metB)
Stationary phase DNA
Marker
purB+ meiB+ (purB/metB) frequency
1 5530 2280 2.42
2 3910 2210 1.77
3 28400 15600 1.82 4 21600 10910 1.98
5 1950 580 3.36
6 10220 3470 2.95
12710 10150 1.25 2.04 26100 24100 1.08 1.64 36000 32900 1.10 1.65 31890 24750 1.29 1.53 2080 1110 1.87 1.80 17400 10310 1.71 1.73 average 1.73 map position 0.21
GERMINATION MUTANTS IN
B. subtilis
TABLE
4Marker frequency of ts markers to try*
71
Group Strain
Swellingts ElongationtS Septationts
is115 is57 is2 is98 tsl ts12 ts76
Experiment 1 1.72 1.21 1.31 1.26 1.10 1.11 0.95 2 1.55 1.33 1.23 1.15 1.10 1.10 0.85
Marker 3 . . . 1.37 1.33 1.30 1.16 1.16 1.04
frequency 4 . . . 1.42 . . . 1.25 , . . . . . 0.88
average 1.64 1.33 1.29 1.25 1.12 1.12 0.93
Map position 0.29 0.59 0.63 0.68 0.84 0.84 1.10
*
Tryptophan-requiring and temperature-sensitive mutants were used as recipients. The donor was W23. Marker frequency of each mutant and the map position were determined as described in Table 2.position of
purB
was calculated to be
0.21.
The map position of
purB
had been
reported as
0.06
by O'SULLIVAN
and SUEOKA
(1967). purB,
however, was not
cotransduced with either
str
(about
0.06)
or
thr (0.27)
by phage
PBSl
(DUBNAU
et
al. 1967).
Mapping
of
ts
m r k m s :
Marker frequencies of each
ts
locus relative to
try
were
studied with seven
ts
mutants that exhibited low back-mutation frequencies and
high competence. The results are shown in Table
4
and Figure
6.
The swellingts
loci mapped on the left half (0 to
0.6
of the chromosome), the elongationts loci
in the middle
(0.6
to
0.7),
and the septationts loci on the right half of the chromo-
some
(0.7
to
1.10).
Confirmation of
mapping b y cotransduction:
Auxotrophic mutants listed in
Table
1
were treated with a phage
PBSl
suspension prepared from a
ts
mutant.
Cotransductants were screened as described in
MATERIALS AND METHODS.The
following sets of genes were transduced together (Table
5
and Figure
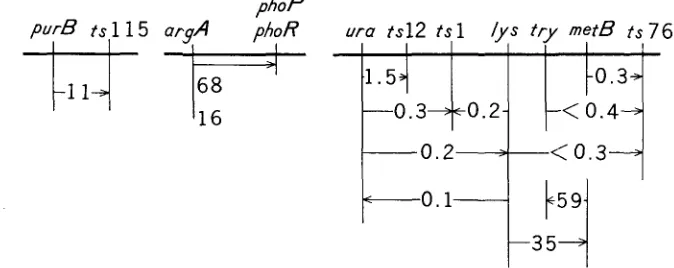
7 ) : purB
and
ts115, ura
and
ts12,
Zys
and
t s l ,
and
m t B
and
ts76.
Linkage was also demon-
strated between
metB
and
t r y
(and
Zys),
ura
and
Zys,
and
argA
and
phoP
(and
try
P U r B
0.29
0.59
0.63 0.68
0.84
m e t B1 . 1 0
t
II
/ I\
I
I0
o.21
t s 1 1 5
t s 5 7
t s 2ts98
t ~ l , t s 1 2
1.00
t s 7 6
I
t s
Swelling
S e p t a t e
cell
+
Dormant
spore
spore
spore
cell
Germinated
I
S w
o l l e n l Elongated
I ,
72
J U N - I C K I NUKUSHINA A N D Y O N O S U K E IKED.4TABLE 5
Results of cotransduction experiments with phage PBSI
Donor Recipient ts115 101
ts 1 M26
2s 1 M26
ts12 M26
ts76 M26
ts76 M26
sfr ts76 tr y
sfr ts76 try
tsl15 SP25
BC9 35
sfr M26
sfr M26
sfr M26
Mu8u5u5 fla
+
160Selected marker
Donor Number of Donor type Frequency' marker colonies recombinant (percent)
purB+ ura
+
ura
+
metB+
t r y+
ts76+
argA
+
argA+
lys+ ura
+
lys+ lys+lys+
metB
+
ts115 tsl tsl ts12 ts76 ts76 ts76+ try+ phoP+ phoR+
ura
+
metB+ lys+ try+ 198 288 486 541 384 298 270 1600 100 67 864 1222 99 27 22 1 1 8 1 0 0 0 68 11 1 3 35 16 11 0.3 0.2 1.5 0.3
<
0.3 <0.4 <O.l 68 16 0.1 0.2 35 59*
Cotransduction frequency(%).
phoR).
The
phoP
and
phoR
are structural and regulatory genes, respectively, of
alkaline phosphatase (MIKI, MINAMI,
and
IKEDA
1965). DUBNAU
et
al.
(1967)
stated that the
ura
and
lys
were not transduced together. I n the present work,
however, these two genes were cotransduced at a frequency of 0.2%.
DISCUSSION
Germination
of
bacterial spores is considered to be a process that can progress
without macromolecular synthesis
(
DEMAIN
and NEWKIRK
1960). The germi-
nated spores, however,
do
synthesize RNA, protein, and DNA in this sequence
phoP
GERMINATION MUTANTS I N
B. subtilis
73
during the course of outgrowth (WOESE
and
FORRO
1960; SAKAKIBARA,
SAITO
and
IKEDA
1965). The mutants described in this paper are vegetative
ts
mutants.
Spores
of
most of these vegetative
ts
mutants fail to develop into swollen spores,
elongated cells, or septate cells when incubated at
48°C.
This fact suggests that the
ts
genes may express their information at different stages of outgrowth.
The mapping data in Figure 6 are of interest. First, swellingts, elongationts,
and septationts groups seem to have their own territories on the chromosome.
Second, the
t sgroups are arranged on the chromosome in the sequence
of develop-
mental processes of bacterial spores.
As
the probability for getting these results by
chance is quite low (less than
0.3%),
we presume that the gene arrangement on
the chromosome may be in some way related to the sequential expression of gene
function during the course of outgrowth.
Do1 and
IGARASHI
(1964) studied messenger
RNA
fractions from three growth
phases of
B. subtilis;
namely, sporulation, germination, and step-down transition.
They indicated that
RNA's
are transcribed from unique genetic loci during mor-
phogenesis. MASTERS
and
PARDEE
(1965) described that enzyme synthesis in
synchronous cultures
of
B.
subtilis
occurred in sequence such that genetic and
enzyme maps were colinear. It was also reported by
TEVETHIA
and MANDEL
(1967) that the sequential replication of the
B. subtilis
chromosome might impose
a sequential character on the transcription process, and by STEINBERG
and
HAL-
VORSON(1968a, b) that gene expression during the outgrowth of
B. cereus
spores
might be controlled at the level of transcription. These observations are com-
patible with our hypothesis. We do not know, however, whether the sequential
expression is controlled at the level of transcription, as the above named investi-
gators have suggested, or at the level of translation. Studies along this line are
in
progress.
We are grateful to Drs. H. SAITO and
H.
HIROKAWA for valuable discussions and suggestions, and to Drs.I.
TAKAHASHI, N. SUEOKA, and J. MARMUR for the gift of the phage and bacterial strains employed.SUM MARY
Developmental processes from spores to vegetative cells in bacteria were studied
with temperature-sensitive mutants of
Bactillus subtilis
Marburg strain 160
try-3.
When mutant spores were incubated at a high temperature, they discontinued
their development
at
various stages of outgrowth, and were classified into four
groups; swellingt8 mutants, elongationts mutants, septationt8 mutants, and an-
other kind of mutant. Seven
ts
mutants that had been chosen at random from
the first three groups were mapped on the chromosome by the marker-frequency
method and by co'transduction with phage PBSI. The mapping data suggested
that the swellingts, elongationts, and septationts mutants might have their
own
74
JUN-ICHI NUKUSHINA AND YONOSUKE IKEDALITERATURE CITED
DEMAIN, A.
L.,
and J. F. NEWKIRK, 1960 Dissociation of spore germination from outgrowth byuse of auxotrophic mutants of Bacillus subtilis. J. Bacteriol. 79: 783-788.
DOI, R. H., and R. T. IGARASHI, 1964 Genetic transcription during morphogenesis. Proc. Natl. Acad. Sci.
U.
S. 52 : 755-762.DUBNAU, D. C., C. GOLDTHWAITE, I. SMITH and J. MARMUR, 1967 Genetic mapping i n Bacillus subtilis. J. Mol. Biol. 27: 163-185.
EDGAR, R. S., G.
H.
DENHART and R.H.
EPSTEIN, 1964 A comparative genetic study of condi- tional lethal mutations of bacteriophage T4D. Genetics 49 : 635-648.EDGAR, R. S., and I. LIELAUSIS, 1964 Temperature-sensitive mutants of bacteriophage T4D; their isolation and genetic characterization. Genetics 49 : 649-662.
MCDONALD, W., and T. S. MATNEY, 1963 Genetic transfer of the ability to grow a t 55°C in Bacillus subtilis. J. Bacteriol. 8 5 : 218-220.
MASTERS, M., and A. B. PARDEE, 1965 Sequence of enzyme synthesis and gene replication dur- ing the cell cycle of Bacillus subtilis. Proc. Natl. Acad. Sci. U. S. 54: 64-70.
MIKI, T., Z. MINAMI and
Y.
IKEDA, 1965 The genetics of alkaline phosphatase formation in Bacillus subtilis. Genetics 52: 1093-1 100.~'SULLIVAN, A., and N. SUEOKA, 1967 Sequential replication of the Bacillus subtilis chromo- some IV. Genetic mapping by density transfer experiment. J. Mol. Biol. 27: 349-368. RYTER, A., P. SCHAEFFER and H. IONESCO, 1966 Classification cytologique, par leur stade de
blocage, des mutants de sporulation de Bacillus subtilis Marburg. Ann. Inst. Pasteur (Paris) 110: 305-315.
Preparation of transforming deoxyribonucleic acid by phenol treatment. Biochim. Biophys, Acta 72 : 619-629.
Incorporation of radioactive amino acids and bases into nucleic acid and protein fractions of germinating spores of Bacillus subtilis. J. Gen. Appl. Microbiol. 11 : 243-254.
SCHAEFFER, A. B., and D. FULTON, 1933 A simplified method of staining endospores. Science 77: 194.
STEINBERG, W., and H. 0.
HALVORSON,
1968a Timing of enzyme synthesis during outgrowth of spores of Bacillus cereusI.
Ordered enzyme synthesis. J. Bacteriol. 95 : 469478.-
1968b Timing of enzyme synthesis during outgrowth of spores of Bacillus cereus11.
Rela- tionship between ordered enzyme synthesis and deoxyribonucleic acid replication. J. Bac- teriol. 95 : 479-489.Transformation of biochemically deficient strains of Bacillus subtilis by deoxyribonucleate. Proc. Natl. Acad. Sci. U. S. 44: 1072-1078.
Genetic transduction in Bacillus subtilis. Biochem. Biophys. Res. Comm. 5: 171-175. - 1965 Localization of spore markers on the chromosome of Bacillus subtilis. J. Bacteriol. 89: 1065-1067.
TANOOKA,
H.,
and Y. SAKABIBARA, 1968 Radioresistant nature of the transforming activity of DNA in bacterial spores. Biochim. Biophys. Acta 150: 130-142.TEVETHIA, M. J., and M. MANDEL, 1967 Transcription of the aligned chromosome of Bacillus subtilis W23. Proc. Natl. Acad. Sci. U. S. 5 8 : 1174-1181.
WOESE, C. R., and J. R. FORRO, 1960 Correlations between ribonucleic acid and deoxyribonucleic acid metabolism during spore germination. J. Bacteriol. 80: 81 1-81 7.
YOSHIKAWA, H., and N. SUEOKA, 1963 Sequential expression of Bacillus subtilis chromosome.
I. Comparison of marker frequencies in exponential and stationary growth phases. Proc. Natl. Acad. Sci. U. S. 49: 559-565.
SAITO, H., and K. MIURA, 1963
SAKAKIBARA, Y.,
H.
SAITO andY.
IKEDA, 1965SPIZIZEN, J., 1958