IL-6 Predicts p16
INK4aExpression, Intracellular
Tenofovir/Emtricitabine Exposure, and Endogenous
Nucleotide Pools in Adults with HIV
Honors Candidate:
Cynthia Lee
PharmD Candidate 2016
UNC Eshelman School of Pharmacy
University of North Carolina at Chapel Hill
Advisor:
Julie B. Dumond, PharmD, MS, BCPS, AAHIVP
Assistant Professor
Division of Pharmacotherapy and Experimental Therapeutics
UNC Eshelman School of Pharmacy
Preface
I worked with Dr. Julie Dumond and colleagues to study the function of cytokines to
aging, antiretroviral drugs, and endogenous nucleotide exposures in HIV-infected
subjects. My involvements in the study include participating in subject recruitment and
visit, assisting in AUC calculations, conducting literature searches, and contributing to
manuscript write-up. The manuscript fully describes the study and is prepared for
publication in HIV Medicine journal.
Abstract
Objectives: It was recently found that p16INK4a, a marker for aging and cellular
senescence, is associated with lower intracellular concentrations of endogenous
nucleotides and nucleos(t)ide reverse transcriptase inhibitors (NRTIs). This study
expands on these findings by determining whether markers of inflammation are
predictive of p16INK4a expression, intracellular metabolite exposure, or endogenous
nucleotide concentrations.
Methods: Samples from HIV-infected adults receiving daily tenofovir/emtricitabine
(TFV/FTC) with either efavirenz (EFV) or atazanavir/ritonavir (ATV/r) were tested for
p16INK4a expression, plasma cytokine concentrations, and intracellular drug
concentrations. Elastic net regression was used to identify cytokines that were
predictive of p16INK4a expression and intracellular metabolite/endogenous nucleotide
exposures. These outcomes were each compared between groups with detectable
Results: Enrolled participants had a median age of 48 years (range 23 - 73). Results of
the elastic net regression determined that individuals with detectable interleukin-6 (IL-6)
concentrations were predicted to have elevated p16INK4a expression and lower exposure
to tenofovir diphosphate (TFV-dp), emtricitabine triphosphate (FTC-tp), and their
respective endogenous nucleotides, deoxyadenosine triphosphate (dATP) and
deoxycytidine triphosphate (dCTP).
Conclusions: Elevated concentrations of IL-6 have been associated with frailty,
morbidity, and mortality in individuals with HIV. Our findings suggest that IL-6 affects
cellular senescence, NRTI pharmacokinetics, and endogenous nucleotide pools in an
aging HIV population.
Introduction
Despite recent advances in antiretroviral (ARV) therapy, HIV-infected patients are at
heightened risk for developing complications typically associated with increased age (1).
Markers of inflammation are chronically elevated in HIV-infected patients and have been
associated with increases in non-AIDS defining morbidity and all-cause mortality (2,3).
Given these consequences, understanding the role inflammation plays in the
pharmacokinetics of ARVs and the progression of HIV is critical to improving outcomes
in aging patients.
Tenofovir and emtricitabine are nucleos(t)ide reverse transcriptase inhibitors (NRTIs)
and are recommended in first-line combination ARV therapies. In order to exert their
their respective metabolites, tenofovir diphosphate (TFV-dp) and emtricitabine
triphosphate (FTC-tp). They work by competing with endogenous nucleotides
(deoxyadenosine triphosphate [dATP] for TFV-dp; deoxycytidine triphosphate [dCTP]
for FTC-tp) for incorporation into the DNA of infected cells, leading to chain termination.
By virtue of this mechanism, it is thought that the ratio of intracellular metabolite to
endogenous nucleotide has a greater influence on antiviral efficacy and toxicity than
intracellular drug concentrations alone (4).
We previously demonstrated that increased expression of p16INK4a, a biomarker for
aging, is associated with lower concentrations of TFV/FTC intracellular metabolites and
endogenous nucleotides (5). The work herein extends upon that study and aims to
determine whether markers of inflammation previously associated with aging and HIV
morbidity / mortality are predictive of p16INK4a expression, intracellular metabolite
exposure, or endogenous nucleotide concentrations.
Methods
Clinical Trial Design
A detailed description of the trial design and eligibility criteria has been previously
published (5). In short, HIV-infected adults were recruited from UNC HealthCare
Infectious Diseases Clinic (Chapel Hill, NC) and the Cone Health Regional Center for
Infectious Diseases (Greensboro, NC). All participants received daily TFV/FTC 300/200
mg with either efavirenz 600mg or atazanavir/ritonavir 300/200mg for at least 2 weeks.
study protocol was approved by the Institutional Review Boards of both institutions
(Clinicaltrials.gov NCT01180075).
Cytokines
At one of the above pharmacokinetic time points, an additional EDTA tube was
collected, centrifuged at 3000 RPMs for 10 minutes, and stored at -80ºC for biomarker
profiling. Analysis was performed in the Duke Regional Biocontainment Laboratory
(RBL) Immunology Unit (Durham, NC) under the direction of Dr. Gregory D. Sempowski
using MILLIPLEX® MAP Human Cytokine/Chemokine Premixed 39 Plex bead-based
assay kit (EMD Millipore Corporation, Billerica, Massachusetts). Due to the large
number of potential predictor variables, a subset of cytokines that have been associated
with HIV and aging was included in the analysis: TNFα, IFNγ, 1ra, 6, 12P40,
IL-12P70, IL-17α, MCP-1, MIP-1α, MIP-1β, MCP-3, MDC, GRO, sCD40L, fractalkine, and
eotaxin (6–8).
Pharmacokinetics & p16INK4a Expression
TFV-dp, FTC-tp, dATP, and dCTP concentrations in peripheral blood mononuclear cells
(PBMCs) were measured using LC-MS/MS in the UNC Center for AIDS Research
Clinical Pharmacology and Analytical Chemistry Laboratory (5). Drug exposure was
measured as area under the curve (AUC) using non-compartmental analysis in Phoenix
Win Nonlin 6.3 (Pharsight, A Certara Company, St. Louis, MO); the linear up/log down
trapezoidal method was used to calculate AUC over the dosing interval. Expression of
p16INK4a was determined using validated PCR-based methods (9) and final values were
Statistical Analysis
The primary goal for this analysis was to identify cytokines that are predictive of p16INK4a
expression and intracellular metabolite/endogenous nucleotide AUCs. Predictor
variables were selected using a general linear model elastic net algorithm, a penalized
regression technique (10) capable of selecting variables that are most predictive
amongst a large number of correlated potential predictors. Tuning parameters for the
elastic net algorithm were chosen via 5-fold cross-validation with the optimal tuning
parameter values chosen to minimize predicted residual sum of squares (CVPRESS).
The cytokine levels were dichotomized as detectable (1) or undetectable (0) for
analysis. Chronological age in years was included as a potential predictor variable. An
exact Wilcoxon rank-sum test was used to assess whether p16INK4a expression and
intracellular metabolite/endogenous nucleotide exposures were different between
groups with detectable and undetectable concentrations of predictive cytokines.
Analyses were conducted in SAS version 9.4 (SAS Institute, Cary, NC) using the
GLMSELECT procedure or R 3.1.2 (r-project.org). A p-value of < 0.05 was considered
statistically significant. These analyses were exploratory in nature, so no adjustment
Results
Study Participants
The study enrolled 79 participants receiving TFV/FTC. Of those, 54 were receiving EFV
and 25 were receiving ATV/r. The median age was 48 years (range 23 - 73) and the
median duration of HIV infection was 10 years. Sixty-one percent of participants were
African-American. Detailed demographics of the study population have been published
(5). One enrolled participant did not have a sample available for cytokine profiling. Of
the remaining 78, five did not provide a sample for p16INK4a measurement and six did
not provide adequate samples for accurate calculation of intracellular
metabolite/endogenous nucleotide exposures. These individuals were excluded from
analysis.
Elastic Net Analysis
P16INK4a expression and AUC measurements were available for 73 and 72 participants,
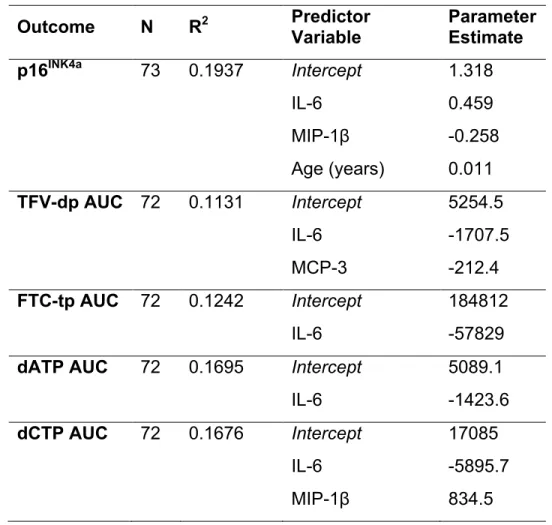
respectively. Table 1 presents the elastic net results for p16INK4a expression and the four
AUC measures using dichotomized cytokine predictor variables. Detectable
concentrations of interleukin-6 (IL-6) were found to be associated with higher p16INK4a
expression and lower AUC of FTC-tp, TFV-dp, dATP, and dCTP. Macrophage
inflammatory protein-1β (MIP-1β) was negatively associated with p16INK4a expression
and positively associated with dCTP exposure. Monocyte chemoattractant protein 3
(MCP-3) was associated with lower TFV-dp exposure. Age positively predicted p16INK4a
Cytokines were dichotomized based on detectability because a sizeable proportion of
raw concentration values were left censored. The R2 values for the optimal model are
presented for each outcome and represent the proportion of variation explained by
predictor variables in the model. The penalized parameter estimates for the selected
variables describe the direction of prediction. The magnitude does not provide for
meaningful comparisons between models and significance testing cannot be applied to
the model parameters as in ordinary least squares (10).
Outcomes of interest were then compared between groups with detectable and
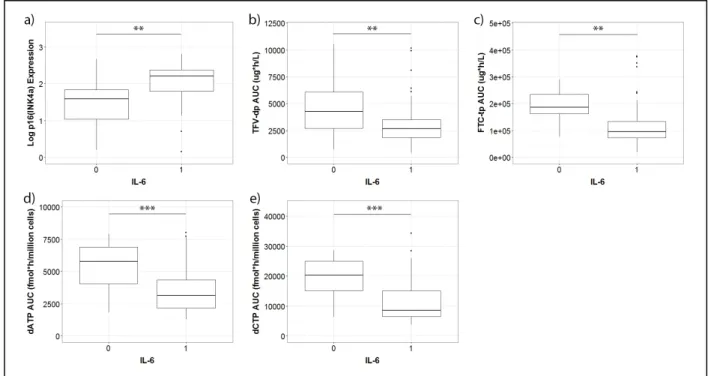
undetectable concentrations of IL-6. Figure 1 shows that participants with detectable
concentrations of IL-6 had significantly higher p16INK4a expression and lower TFV-dp,
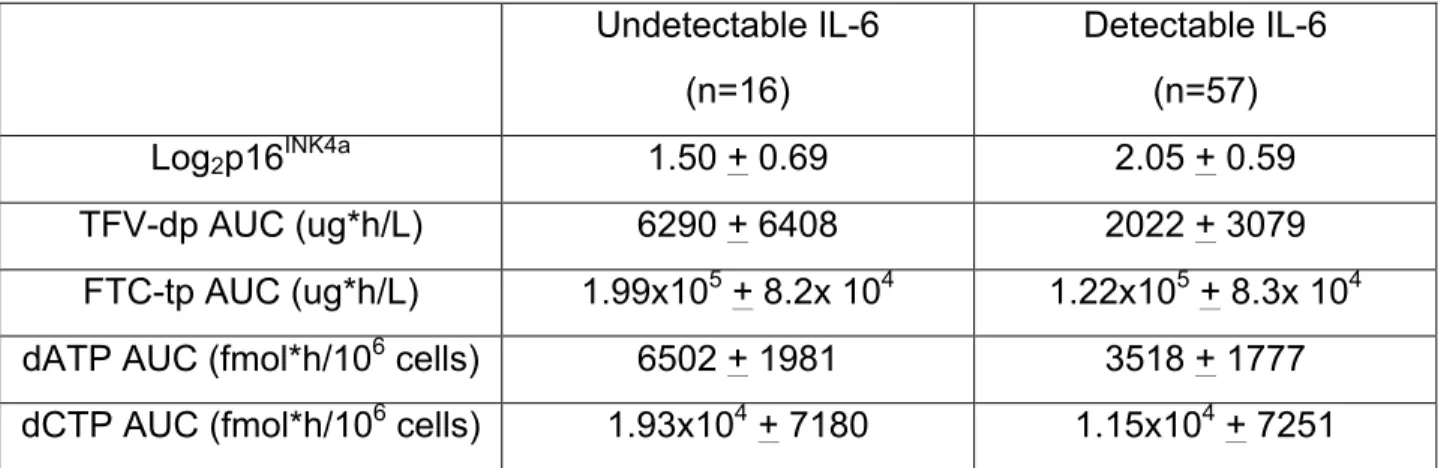
FTC-tp, dATP, and dCTP exposures in PBMCs. Means and standard deviations are
provided in Table 2. Comparisons between groups with detectable and undetectable
concentrations of MIP-1β and MCP-3 were not statistically significant (Wilcoxon p >
0.05).
Discussion
Results from the elastic net regression showed detectable concentrations of IL-6
predicted increased expression of p16INK4a. PIM-1 is a proto-oncogene expressed in the
hematopoietic and lymphoid system. It encodes for a serine/threonine protein kinase
that regulates cell cycle progression and apoptosis (12). IL-6 mediates PIM-1
expression through signal transducer and activator of transcription-3 (STAT3), an
PIM-1 increases markers of aging and cellular senescence, including p16INK4a. The
relationship between IL-6 and p16INK4a seen in our HIV positive cohort could be
explained by through this mechanism.
It was also observed that participants with detectable IL-6 had decreased exposure to
TFV-dp and FTC-tp. This finding could be explained by membrane transporter
modulation. TFV is a known substrate for efflux transporters P-glycoprotein (P-gp) and
breast cancer resistance protein (BCRP), and FTC is transported by multidrug
resistance-associated protein-1 (MRP-1) (14). IL-6 has been shown to directly increase
P-gp and BCRP cell-surface expression through the PIM-1 pathway (15) and MRP-1
expression through unknown mechanisms in human cells lines (16). Modulation of
membrane transporters is already known to affect intracellular accumulation of protease
inhibitors (17). Increased expression of these efflux transporters on PBMCs could result
in decreased intracellular NRTI concentrations.
Detectable concentrations of IL-6 were also predictive of lower amounts of dATP and
dCTP. Reduced exposure to these nucleotides may be related to cell cycle arrest in
senescent cells. Demand for nucleotide biosynthesis varies during the cell cycle and is
highest during S-phase (Bayes, et al. 2014). To meet this heightened demand, enzymes
involved in the synthesis of nucleotides and deoxynucleotides, such as phosphoribosyl
pyrophosphate (PRPP) and ribonucleotide reductase (RNR), are upregulated (18). IL-6
has been described to induce premature senescence in human fibroblasts (19). Since
senescent cells are arrested in the G-1 phase and are no longer dividing, demand for
endogenous nucleotides, such as dATP and dCTP, remains low and their synthesis is
MIP-1β was selected as a predictor of p16INK4a expression and dCTP AUC. MCP-3 was
predictive of TFV-dp exposure. However, it was decided to focus on IL-6, as it was
consistently predictive of all outcomes of interest.
The results from this work are consistent with our earlier findings. P16INK4a was found to
be negatively associated with drug metabolite and endogenous nucleotide exposures.
Our analysis demonstrated that detectable concentrations of IL-6 are positively
predictive of p16INK4a expression, while negatively predictive of intracellular drug
metabolite and nucleotide exposures. This relationship suggests that cytokines, such as
IL-6, could play a role in the inverse association between p16INK4a expression and
drug/nucleotide exposure previously seen.
Also, although a relationship between IL-6 and intracellular metabolite/endogenous
nucleotide exposures is evident, our study lacks the ability to identity a clear causal
mechanism for the relationships being seen. Future in vitro and in vivo studies are
planned to confirm these relationships and explore potential mechanisms.
HIV-infected patients exhibit significantly higher plasma concentrations of IL-6
compared to uninfected controls (20) and IL-6 has been associated with increased
all-cause mortality in this population (2). The results of this study suggest that IL-6 affects
cellular senescence, NRTI pharmacokinetics, and endogenous nucleotide pools. These
findings emphasize the importance that markers of inflammation play in the treatment of
HIV in an aging population, and further support the idea of cytokines as potential drug
Acknowledgements
The authors wish to thank study participants, as well as the UNC Clinical and
Translational Research Center (UL1RR02574), the UNC Health Care Infectious
Diseases Clinic, and the Cone Health Regional Center for Infectious Diseases staff for
their contribution.
Biomarker profiling was performed in the Duke Regional Biocontainment Laboratory
(RBL) Immunology Unit under the direction of Dr. Gregory D. Sempowski. Phoenix Win
Nonlin software is generously provided to the Division of Pharmacotherapy and
Experimental Therapeutics in the UNC Eshelman School of Pharmacy through the
Certara Center of Excellence program. Dr. Angela Kashuba graciously provided
oversight of drug concentration analysis in the UNC Center for AIDS Research Clinical
Pharmacology and Analytical Chemistry Laboratory. We also thank Jordan Messer and
Jingxian Chen for assisting with AUC calculations.
JBD is supported by K23AI093156; funding for this work was provided in part by
UL1RR02574.
OF, MC, HMAP, KRM, CS, NW, SM, MGH are supported in part by the UNC Center for
AIDS Research (P30AI050410).
MC is supported by T32-GM086330.
CT and NES are supported by R01-AG024379-10.
Disclosures
KRM previously received research support from grants awarded to UNC from Merck,
References
1. Horvath S, Levine a. J. HIV-1 Infection Accelerates Age According to the Epigenetic Clock. J Infect Dis [Internet]. 2015;212. Available from: http://jid.oxfordjournals.org/lookup/doi/10.1093/infdis/jiv277
2. Kuller LH, Tracy R, Belloso W, Wit S De, Drummond F, Lane HC, et al. Inflammatory and Coagulation Biomarkers and Mortality in Patients with HIV Infection. PLoS Med [Internet]. 2008;5(10):e203. Available from: http://www.plosmedicine.org/article/info:doi/10.1371/journal.pmed.0050203
3. Tenorio AR, Zheng Y, Bosch RJ, Krishnan S, Rodriguez B, Hunt PW, et al. Soluble Markers of Inflammation and Coagulation but Not T-Cell Activation Predict Non-AIDS-Defining Morbid Events During Suppressive Antiretroviral Treatment. J Infect Dis [Internet]. 2014;210:1–12. Available from: http://www.ncbi.nlm.nih.gov/pubmed/24795473
4. Goicoechea M, Jain S, Bi L, Kemper C, Daar ES, Diamond C, et al. Abacavir and tenofovir disoproxil fumarate co-administration results in a nonadditive antiviral effect in HIV-1-infected patients. AIDS. 2010;24(5):707–16.
5. Dumond JB, Francis O, Cottrell M, Trezza C, Prince HM, Mollan K, et al. Tenofovir/emtricitabine metabolites and endogenous nucleotide exposures are associated with p16INK4a expression in subjects on combination therapy. Antivir
Ther [Internet]. 2016; Available from:
http://www.intmedpress.com/journals/avt/abstract.cfm?id=3017&pid=48
6. Nixon DE, Landay AL. Biomarkers of immune dysfunction in HIV. Curr Opin HIV AIDS. 2010;5(6):498–503.
7. Paiardini M, Müller-Trutwin M. HIV-associated chronic immune activation. Immunol Rev. 2013;254(1):78–101.
8. Abbondanzo SJ, Chang SL. HIV-1 Transgenic Rats Display Alterations in Immunophenotype and Cellular Responses Associated with Aging. PLoS One
[Internet]. 2014;9(8):e105256. Available from:
http://dx.plos.org/10.1371/journal.pone.0105256
9. Nelson J a E, Krishnamurthy J, Menezes P, Liu Y, Hudgens MG, Sharpless NE, et al. Expression of p16(INK4a) as a biomarker of T-cell aging in HIV-infected patients prior to and during antiretroviral therapy. Aging Cell [Internet].
2012;11(5):916–8. Available from:
http://www.pubmedcentral.nih.gov/articlerender.fcgi?artid=3697001&tool=pmcentr ez&rendertype=abstract
10. Zou H, Hastie T. Regularization and Variable Selection via the Elastic Net. J R Stat Soc Ser B. 2005;67(2):301–20.
[Internet]. 2009;8(4):439–48. Available from: http://www.pubmedcentral.nih.gov/articlerender.fcgi?artid=2752333&tool=pmcentr ez&rendertype=abstract
12. Bachmann M, Möröy T. The serine/threonine kinase Pim-1. Int J Biochem Cell
Biol [Internet]. 2005;37(4):726–30. Available from:
http://linkinghub.elsevier.com/retrieve/pii/S1357272504003991
13. Jin B, Wang Y, Wu CL, Liu KY, Chen H, Mao Z Bin. PIM-1 modulates cellular senescence and links IL-6 signaling to heterochromatin formation. Aging Cell. 2014;13(5):879–89.
14. Kis O, Robillard K, Chan GNY, Bendayan R. The complexities of antiretroviral drug – drug interactions : role of ABC and SLC transporters. 2009;(December). 15. Natarajan K, Bhullar J, Shukla S, Burcu M, Chen ZS, Ambudkar S V., et al. The
Pim kinase inhibitor SGI-1776 decreases cell surface expression of P-glycoprotein (ABCB1) and breast cancer resistance protein (ABCG2) and drug transport by Pim-1-dependent and -independent mechanisms. Biochem Pharmacol [Internet]. Elsevier Inc.; 2013;85(4):514–24. Available from: http://dx.doi.org/10.1016/j.bcp.2012.12.006
16. Lee G, Piquette-Miller M. Influence of IL-6 on MDR and MRP-mediated multidrug resistance in human hepatoma cells. CanJ Physiol Pharmacol. 2001;79(10):876– 84.
17. Hoggard PG, Back DJ. Intracellular pharmacology of nucleoside analogues and protease inhibitors: role of transporter molecules. Curr Opin Infect Dis [Internet].
2002;15(1):3–8. Available from:
http://www.ncbi.nlm.nih.gov/entrez/query.fcgi?cmd=Retrieve&db=PubMed&dopt= Citation&list_uids=11964899
18. Lane a. N, Fan TW-M. Regulation of mammalian nucleotide metabolism and biosynthesis. Nucleic Acids Res [Internet]. 2015;43(4):2466–85. Available from: http://nar.oxfordjournals.org/lookup/doi/10.1093/nar/gkv047
19. Kojima H, Inoue T, Kunimoto H, Nakajima K. IL-6-STAT3 signaling and premature senescence. Jak-Stat [Internet]. 2013;2(4):e25763. Available from: http://www.pubmedcentral.nih.gov/articlerender.fcgi?artid=3876432&tool=pmcentr ez&rendertype=abstract
20. Borges ÁH, O’Connor JL, Phillips AN, Rönsholt FF, Pett S, Vjecha MJ, et al. Factors Associated With Plasma IL-6 Levels During HIV Infection. J Infect Dis
[Internet]. 2015;212(4):585–95. Available from:
Figure and Tables
Figure 1: Boxplots showing the distributions of a) p16INK4a expression, b) TFV-dp AUC, c) FTC-tp AUC, d) dATP AUC, and e) dCTP AUC between participants with undetectable (0) and detectable (1) levels of IL-6.
Each box corresponds to the inter-quartile range (middle 50% of observations) for the group. The median is indicated by the horizontal line within the box. The distance between the upper and lower whiskers represents the range of the data that are not considered outliers. Outliers are denoted by a closed circle (•). Participants with detectable concentrations of IL-6 exhibited significantly higher expression of p16INK4a than those with undetectable concentrations (a). Conversely, participants with detectable IL-6 concentrations had a lower TFV-dp AUC (b), FTC-tp AUC (c), dATP AUC (d), and dCTP AUC (e). ** p <0.01, *** p < 0.001
Outcome N R2 Predictor
Variable
Parameter Estimate
p16INK4a 73 0.1937 Intercept 1.318
IL-6 0.459
MIP-1β -0.258
Age (years) 0.011
TFV-dp AUC 72 0.1131 Intercept 5254.5
IL-6 -1707.5
MCP-3 -212.4
FTC-tp AUC 72 0.1242 Intercept 184812
IL-6 -57829
dATP AUC 72 0.1695 Intercept 5089.1
IL-6 -1423.6
dCTP AUC 72 0.1676 Intercept 17085
IL-6 -5895.7
MIP-1β 834.5
Table 1: Elastic net results using dichotomized cytokine values as predictor
variables for p16INK4a expression, TFV-dp AUC, FTC-tp AUC, dATP AUC, and dCTP AUC.
The R2 value is the proportion of variation explained by predictor variables in the model.
Variables chosen by the elastic net algorithm that best predict each outcome are shown with their corresponding penalized parameter estimates. The penalized parameter estimates show whether a variable is positively or negatively predictive of the outcome. Detectable concentrations of IL-6 are positively predictive of p16INK4a expression and negatively predictive of TFV-dp AUC, FTC-tp AUC, dATP AUC, and dCTP AUC. Detectable concentrations of MIP-1β predict p16INK4a expression and dCTP AUC.
Detectable MCP-3 is a predictor for TFV-dp AUC.
AUC, area under curve; TFV-dp tenofovir diphosphate; FTC-tp, emtricitabine
Undetectable IL-6
(n=16)
Detectable IL-6
(n=57)
Log2p16INK4a 1.50 + 0.69 2.05 + 0.59
TFV-dp AUC (ug*h/L) 6290 + 6408 2022 + 3079
FTC-tp AUC (ug*h/L) 1.99x105 + 8.2x 104 1.22x105 + 8.3x 104
dATP AUC (fmol*h/106 cells) 6502 + 1981 3518 + 1777
dCTP AUC (fmol*h/106 cells) 1.93x104 + 7180 1.15x104 + 7251
Table 2: Comparison of p16INK4a expression, TFV-dp AUC, FTC-tp AUC, dATP AUC, and dCTP AUC measurements between participants with undetectable and detectable concentrations of IL-6.
Values are reported as mean + standard deviation.