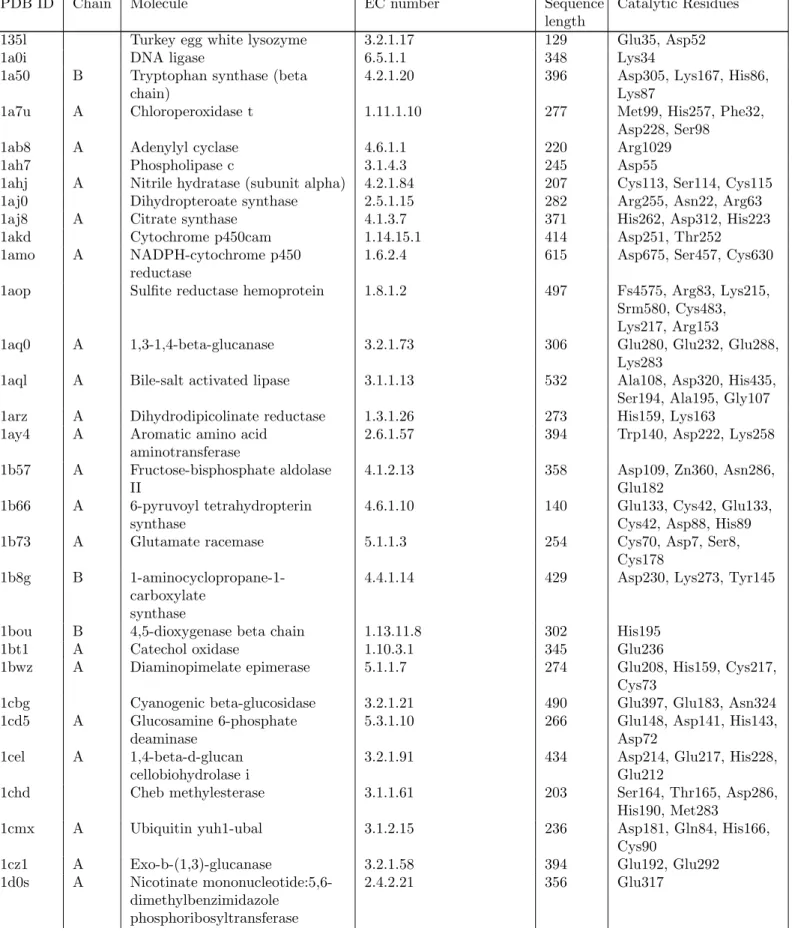
Catalytic Residues
6
0
0
Full text
(2)
(3)
(4)
(5)
(6)
Figure
Related documents
• Phua KH, Towards a Comparative Analysis of Health Systems Reforms in the Asia-Pacific Region, Asia-Pacific Journal of Public Health 14(1): 9-16, 2002. • Phua KH, The
Shares of Big effectively transferred to Small 40 Multiply by: Fair value per share of Big’s shares 480 Fair value of consideration effectively transferred 19,200 Goodwill (gain
In weighing the evidence, the Panel took into account that consumption of non-digestible carbohydrates results in reduced post-prandial blood glucose (and
This paper produces annual aggregate and industry productivity series for the market sector of the New Zealand economy for the period 1988 to 2002, using index number techniques and
craniostenosis, meaning a structured or narrowed skull, but was convinced by Sear 47 to instead call this entity Craniosynostosis, the premature closure of cranial sutures, has
5 From DWS to Prediction •Reporting •Case Mgt •Return Scoring •Enforcement Historical Returns •Current Returns •Collections •Case Management.. DWS – Existing
Tissue damage, typically to the feet or hands that occurs when exposure to cold damages beds of capillary or small blood vessels in the skin. Chilblains Signs
Another problem with adaptive algorithms applied to ad-hoc networks arises when changes in the network occur too frequently to allow routing updates to propagate throughout all
