R E S E A R C H
Open Access
Differential myofiber-type transduction
preference of adeno-associated virus
serotypes 6 and 9
Muhammad Riaz
1*, Yotam Raz
1, Elizabeth B. Moloney
2, Maaike van Putten
1, Yvonne D. Krom
1,
Silvere M. van der Maarel
1, Joost Verhaagen
2,3and Vered Raz
1Abstract
Background:Gene therapy strategies are promising therapeutic options for monogenic muscular dystrophies, with several currently underways. The adeno-associated viral (AAV) vector is among the most effective gene delivery systems. However, transduction efficiency in skeletal muscles varies between AAV serotypes, with the underlying factors poorly understood. We hypothesized that myofiber-specific tropism differs between AAV serotypes.
Methods:We developed a quantitative histology procedure and generated myofiber pattern maps for four myosin heavy chain (MyHC) isotypes. We compared myofiber pattern maps between AAV6 or AAV9 injected tibialis anterior muscle in mice. We correlated MyHC expression with AAV-derived green fluorescence protein (GFP) expression using statistical models.
Results:We found that MyHC-2x expressing myofibers display a significantly higher preference for AAV transduction, whereas MyHC-2b expressing myofibers negatively correlated with AAV transduction. In addition, we show that AAV9-mediated transduction is enriched in myofibers expressing MyHC-1 and MyHC-1/2a. Moreover, AAV9-mediated transduction can predominantly be predicted by the expression of MyHC isotypes. In contrast, AAV6 transduction can be predicted by myofiber size but not by myofiber types.
Conclusions:Our findings identify differences between AAV6 and AAV9 for myofiber-type preferences, which could be an underlying factor for mosaic transduction of skeletal muscle. Adjusting AAV serotype for specific muscle conditions can therefore improve transduction efficacy in clinical applications.
Keywords:Skeletal muscle, Adeno-associated viral vectors, AAV serotypes, Gene therapy, Myofiber types
Background
Adeno-associated viruses (AAV) have been successfully engineered as gene-delivery vectors for efficient transduc-tion of post-mitotic cells including skeletal muscles [1]. AAV-derived transgene expression is sustainable, consist-ent over time, and safe in preclinical applications [2]. Ongoing innovations in AAV vector engineering have further improved their suitability for application in clinical trials. Over the past decade, several AAV serotypes with variable tropism have been developed for superior trans-duction [3, 4], and most of these have been suggested for
use in skeletal muscle transduction [5]. However, three AAV serotypes, AAV1, AAV6, and AAV9, are mostly used in pre-clinical studies [2]. Broad tropism, safety, and high transduction efficiency are among the main considerations for serotype utilization. However, this is not fully exploited and results are not always consistent between studies. For example, in dogs, AAV1 tropism is broader and not specific to skeletal muscles as compared to AAV9 [6], but AAV9 tropism in mice was suggested to be broader than AAV6 or AAV1 [2, 4]. Furthermore, in mice, AAV6 shows more resistance to proteasome-mediated degradation compared with AAV1 [7]. Elucidating tissue-specific trop-ism of AAV serotypes is therefore necessary to understand transduction efficiency and targeted application in the changing environment of a tissue like skeletal muscle. * Correspondence:[email protected]
1
Department of Human Genetics, Leiden University Medical Center, Building 2, Room R3-17, Einthovenweg 20, 2333 ZC Leiden, The Netherlands Full list of author information is available at the end of the article
Muscle tissue is composed of contractile fibers (also known as myofibers) that can be generally subdivided into four interchangeable types, marked by the expres-sion of four myosin heavy chain (MyHC) isotypes: MyHC-2b, MyHC-2x, MyHC-2a, and MyHC-1 [8–10]. The distribution of the myofiber types changes dynamic-ally to accommodate alterations in muscle function and metabolism [11]. Moreover, in muscular disorders, a switch in myofiber types is accompanied by muscle disuse and atrophic conditions [12, 13]. However, prefer-ence of AAVs for myofiber types and its contribution to overall transduction efficiency has not been thoroughly investigated. We studied myofiber-type preference in mouse tibialis anterior (TA) muscle following AAV6 or AAV9 intramuscular administration. We found that while AAV6 transduced smaller myofibers with a higher preference for MyHC-2x expressing myofibers, AAV9 transduction can be determined by MyHC expression. We therefore suggest that myofiber types of skeletal muscle can play a role in the selection of AAV serotypes for effective transduction and treatment.
Methods
AAV vectors
The AAV expression pTRCGW vector containing
inverted terminal repeats (ITRs) of AAV2, EGFP (named here green fluorescence protein (GFP)) cDNA under control of the cytomegalovirus (CMV) promoter and poly-adenylation signal of the simian virus 40, and the wood-chuck post-transcriptional regulatory element (WPRE) was used for AAV production. AAV6 and AAV9 particles expressing the same vector were generated and purified as described before [14]. Briefly, for each AAV serotype, six 15-cm petri dishes containing 12.5 × 106HEK 293 T cells were transfected with branched polyethyleneimine (Sigma, St Louis, MO, USA). For AAV6, pTRCGW was co-transfected with the serotype-specific helper plasmid (ratio 3:1, total DNA 50μg/plate). For AAV9, pTRCGW was co-transfected with the helper plasmid pAdΔF6 and the serotype-specific helper plasmid (ratio 2:2:1, total DNA 62.5μg/plate). Helper plasmids for AAV6 were kindly pro-vided by JA Kleinsschmidt [15] and for AAV9 by JM Wilson [16]. Transfected cells were grown in 20 ml Iscove’s modified Eagle’s medium with 10 % fetal calf serum (FCS), glutamate, and penicillin/streptomycin (Invi-trogen, Carlsbad, CA, USA) for 2 days. Cells were lysed and virus particles were harvested and purified using a density gradient of Iodixanol as published before [17] with slight modifications. Briefly, cells containing virus particles were harvested into phosphate-buffered solution (PBS) and lysed by doing freeze-thaw in dry ice/ethonal for three times in the presence of DNAse (10 mg/ml). The lysate was incubated for an hour at 37 °C and centrifuged at 4000gfor 30 min. The supernatant containing AAVs was
then underlaid with a gradient of 15, 25, 40, and 60 % Iodixanol in water (Nycomed Pharma AS, Oslo, Norway) in Beckman Quick-Seal Polyallomer tube using a pasteur pip-ette. The tube was sealed and placed into a NVT90 rotor (Beckman Instruments) and centrifuged at 69,000 rpm for 70 min at 16 °C. Fractions of approximately 3 ml of the Iodixanol gradient (1 ml of 60 % layer and 2 ml of 40 % layer) were collected from the bottom of the tube. The AAV derived from the Iodixanol gradient was further diluted 10 times with PBS, pH 7.5, to reduce the viscosity of the Iodixanol, and was subsequently concentrated using Amicon Ultra-15 centrifugal filter units (Milli-pore, Amsterdam, The Netherlands). The removal of cellular impurities in AAV stocks were further con-firmed using protein gel electrophoresis and electron microscopy as described before [17]. Virus titers were determined by quantitative PCR using a primer-set targeting the WPRE sequence of the expression cas-sette and indicated as genomic copies per unit vol-ume (gc/ml) [14]. Genomic copies therefore represent only particles containing the expression cassette. All AAV stocks were kept at −80 °C prior to injections.
Mouse strain and AAV particles injection
Male C57BL/6Jico mice of 7–8-week old were purchased from Jackson laboratories. After 1 week of acclimatization, AAV6 or AAV9 particles (2.13 × 1010gc in 50μl PBS; indi-cated in the text 2 × 1010 gc) were intramuscular injected into either left or right tibialis anterior muscles. Addition-ally, AAV9 (2.13 × 1011gc; indicated in the text 2 × 1011gc) and PBS control were injected into right or left TA muscles, respectively. Five mice were used per injection set. Injec-tions were carried out under general anesthesia using 2 % isoflurane (Pharmachemie BV, Haarlem, The Netherlands). Mice were housed in ventilated cages with sterile bedding, water, rodent food, and air in DM-III containment level. Experiments were carried out in accordance with the Animal Research: Reporting of In Vivo Experiments (AR-RIVE) guidelines [18]. An animal research protocol [#13113] was approved by the Institutional Animal Ethical Committee (DEC), Leiden University Medical Center, Leiden, the Netherlands.
In vivo fluorescence imaging, muscle collection, in vitro imaging, and image quantification
Four-week post-injection mice were sacrificed by cervical dislocation, and TA muscles were collected and immersed in liquid nitrogen cold isopentane for about 30–45 s and then stored at−80 °C for further processing. Transverse cryosections of 10-μm thick were made with a cryostat (Leica CM3050S) and pasted on superfrost plus glass slides (Menzel-Gläser; Thermo scientific) from four alternating parts across the muscle (Additional file 1: Figure S1). Additional tissue from the intervening areas was used for RNA isolation, ensuring representation of the whole muscle during data collection. Histological ana-lyses of the muscle sections included hematoxylin and eosin staining [19] and immunofluorescence for myofiber typing. The untreated cryosections were directly incubated with a primary antibody to Laminin (1:1000; ab11575, Abcam). When indicated, a supernatant of hybridoma 6H1 (1:5, Developmental Studies Hybridoma Bank (DSHB), USA), detecting MyHC-2x was co-incubated. These anti-bodies were detected with secondary anti-rabbit-alexa-647 and anti-mouse-alexa-546 (Molecular Probes, Invitrogen). A mixture of monoclonal antibodies to 1, MyHC-2a, and MyHC-2b isotypes (hybridoma BA-D5, SC-71 and BF-F3, respectively (DSHB)), conjugated with alexa-350, alexa-594, and alexa-488, respectively was generated as describe before [8]. Antibody production from hybridomas and fluorophore conjugation was carried out as described earlier [10]. All antibody incubations were carried out in PBS containing 0.05 % tween (PBST) and 5 % dry milk, and washes were carried out with PBST. Slides were mounted with Aqua Polymount (Polyscience). Incubation with only anti-mouse conjugated secondary was carried out to ex-clude nonspecific binding to GFP positive myofibers. Im-aging of GFP was carried out in untreated sections that were directly mounted with Polymount containing 4′ ,6-dia-midino-2-phenylindole (DAPI). Imaging was carried out with either DM5500 or TCS-SP5 (confocal) microscopes (Leica) using LAS AF software versions: 2.3.6 (DM5500) and 2.5.1.6757 (TCS-SP5), respectively.
Image quantification was carried out with ImageJ, ver-sion 1.48 [20]. Per myofiber mean fluorescence intensity (MFI) and cross-sectional area (CSA) were documented after segmenting sections based on Laminin staining. Subsequently, MFI values were corrected for background and normalized for a fluorophore constant, which was determined from unbound fluorochrome per microscope and camera combination: (DM 5500: 430 nm = 0.735; 488 nm = 0.067; 596 nm = 0.172 and SP5: 488 nm = 0.003; 546 nm = 3.90).
Quantitative RT-PCR
RNA isolation and RT-qPCR were performed as de-scribed previously [21]. Messenger RNA (mRNA) fold changes were calculated after normalizing to the average CT values of Hprt and Gapdh housekeeping genes and
to the mean of PBS-injected TA muscles. Primers used in this study are listed in Additional file 1: Table S1.
Statistical analyses
Fold changes and statistical significance of mRNA were determined with the Student’st test in GraphPad Prism version 6.02.P values <0.05 were considered statistically significant. Statistical analyses of myofibers were carried out on natural log transformed values of the MFI (normalized to the fluorophore constant). A correlation between MFI of GFP and MyHC isotypes was assessed in a multivariate linear regression model with GFP MFI as a dependent variable and MyHC MFIs as independent variables. In a second multivariate model, CSA was added into the first model as an independent variable. Both models were stratified based on AAV serotypes. In addition, Pearson correlation was used to assess correla-tions between CSA, GFP, and the MFI of each MyHC isotype. Analyses were performed with IBM SPSS Statis-tics version 20.0.
Results
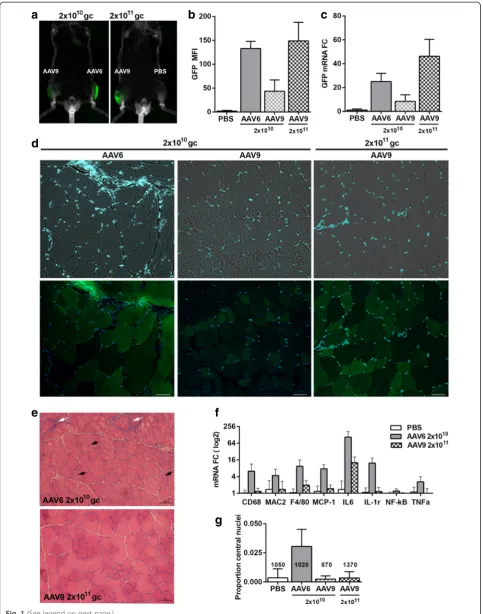
Enhanced transduction efficiency and inflammatory response by AAV6
a
d
b
e
f
g
c
IL6 (Fig. 1f), suggesting that inflammation in TA muscle is not activated by AAV9 administration.
Inflammation in muscles is associated with damaged muscles as characterized by leaky membranes and cen-tral nuclei [11]. In the AAV6-injected muscles, we found an increase in myofibers containing central nuclei (Fig. 1e and Additional file 1: Figure S4). About 3 % of myofibers in AAV6-injected TA muscles have central nuclei, but in the AAV9-injected muscles, the percentage of central nuclei was not different from the PBS injected muscles (Fig. 1g). Moreover, in the AAV6 but not in AAV9-injected muscles, we found regions with damaged myofi-bers and dispersed GFP fluorescence across myofimyofi-bers (Additional file 1: Figure S5). This dispersed GFP fluor-escence differed from a fiber-restricted localization of GFP in the AAV9-injected muscles (Fig. 1d).
Preferential transduction of specific myofiber types by AAV6 and AAV9
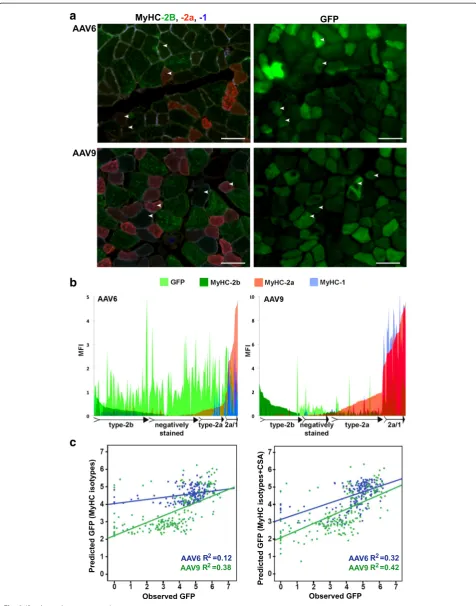
Histological analysis revealed mosaic GFP expression in both AAV6- and AAV9-injected muscles. We investigated whether myofiber types could underpin differences in muscle transduction. We first assessed a simple correlation between GFP MFI and myofiber CSA. In both AAV6- and AAV9-transduced muscles, we found a negative correlation between GFP and myofiber CSA (Additional file 1: Table S2A). This negative correlation could partly be explained by higher concentration of GFP fluorescence in smaller myofibers. However, the correlation between CSA and GFP was much lower in AAV9-injected muscles compared with that of AAV6 (−0.39 and −0.55, respectively). Myofiber CSA was also correlated with the expression levels of MyHC isotypes (Additional file 1: Table S2A). We therefore further assessed whether AAV-mediated gene expression is affected by myofiber types; we correlated GFP expression per myofiber with the expression levels of myofiber types (Additional file 1: Figure S6). MFI of both GFP and each of MyHC fluorophores were measured per myofiber (Fig. 2a). Using a Pearson correlation, we found a negative correl-ation between MyHC-2b expression and GFP in both AAV6- and AAV9-injected muscles, whereas MyHC-2a expression positively correlated with GFP. The expression of MyHC-1 positively correlated with GFP in only
AAV9-transduced muscles (Additional file 1: Table S2B). Plotting MFI of all four fluorophores per myofiber against GFP MFI of corresponding myofibers showed a negative preference of GFP expression in MyHC-2b expressing myofibers (Fig. 2b). Furthermore, we found a strong correlation be-tween MyHC-2a and MyHC-1 expression in these muscles, but not between MyHC-2b and either MyHC-2a or MyHC-1 (Additional file 1: Table S2C). These analyses sug-gest hybrid myofibers co-expressing 1 and MyHC-2a; however, hybrid myofibers expressing MyHC-2b and either MyHC-2a or MyHC-1 were not identified in TA muscles.
Since we found hybrid myofibers co-expressing MyHC isotypes (Fig. 2b), we next assessed the contribution of each MyHC isotype to GFP expression using a multivariate model. In the AAV6-transduced muscles, a weak positive correlation was found between MyHC-2a expression and GFP (regression coefficient (β) = 0.25; P= 0.007; Table 1, Model 1). In the AAV9-transduced muscles, however, MyHC-2a expression negatively correlated with GFP whereas MyHC-1 positively correlated with GFP (β=−0.38,
P< 0.001; β= 0.80, P< 0.001; respectively, Table 1, Model 1). The negative correlation between MyHC-2b and GFP remained unchanged in the multivariate model (Table 1, Model 1). As we found a correlation between GFP and CSA (Additional file 1: Table S2A), we then added CSA as an additional variable into the multivariate model. Surpris-ingly, in the AAV6-transduced muscles, only the CSA remained significant (Table 1, Model 2) whereas, in the AAV9-transduced muscles, a strong correlation between GFP expression and the aforementioned MyHC isotypes persisted (Table 1, Model 2). Scatter plots between the observed GFP and the GFP calculated from the multivariate models (predicted GFP) showed that in the AAV6- but not AAV9-transduced muscles, GFP MFI prediction was im-proved by threefold when CSA was added to the model (Table 1 and Fig. 2c). Together, these data suggest that AAV9-mediated transduction is determined by myofiber types whereas AAV6 transduction is determined primarily by myofiber size.
The MFI distribution plots (Fig. 2b) suggest that GFP positive myofibers are enriched in the negatively stained population of myofibers. In this staining protocol, the (See figure on previous page.)
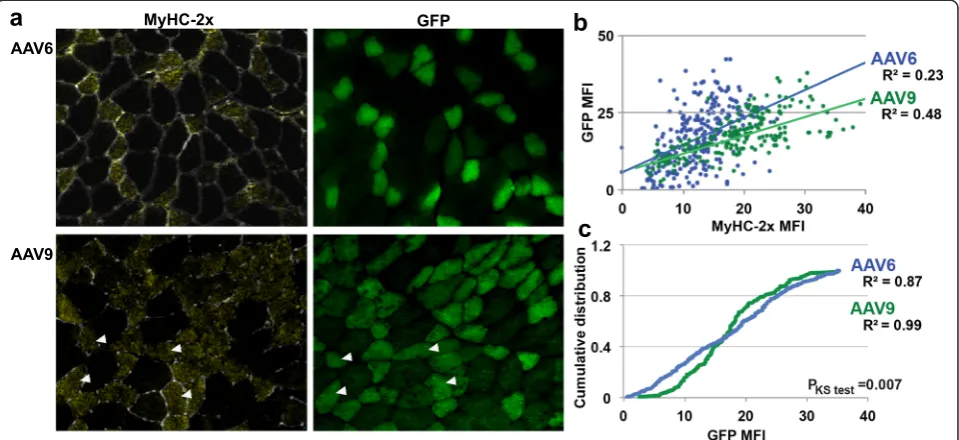
a
b
c
negatively stained myofibers are regarded as MyHC-2x isotype expressing myofibers [8]. To assess whether GFP positive myofibers are indeed MyHC-2x expressing myo-fibers, we stained muscle sections with an antibody to MyHC-2x isotype (Fig. 3a). Primary antibody missing control was performed to exclude a possibility of prefer-ential adherence of the conjugated secondary antibody alexa-546 with GFP positive myofibers (Additional file 1: Figure S7). A strong positive correlation was found between MyHC-2x expression and GFP in myofibers from AAV6- or AAV9-transduced muscles (Pearson cor-relation:P< 0.001) (Fig. 3b). Interestingly, a better fitness (>2-fold) to the linear regression line was found in the AAV9 muscles as compared with AAV6 (R2= 0.48 or 0.23, respectively) (Fig. 3b). Commutative correlation plot of GFP MFI also confirmed better fitness to the regression line (R2) in AAV9-transduced myofiber com-pared with AAV6 fibers (Fig. 3c). These data suggest that both AAV6 and AAV9 show transduction preference for MyHC-2x expressing myofibers with a higher preference for AAV9.
Discussion
In the last couple of decades, several AAV serotypes have been developed for effective gene delivery into various tissues including skeletal muscle with AAV vectors emer-ging as promising vehicles in clinical trials for monogenic musculoskeletal diseases including Duchenne and limb-girdle muscular dystrophy [23, 24]. Muscle tissue is com-posed of different but interchangeable myofibers, which determine muscle contraction and tissue stabilization. Here, we show that the transduction of myofiber types differs between AAV serotypes, using AAV6 and AAV9 as
an example. The same expression vector was packaged in AAV6 and AAV9, and a standard protocol of virus parti-cles generation and purification was used for both sero-types. This ensures that all observed differences are attributed to differences in AAV serotype. Consistent with previous studies, we found that AAV transduction of skel-etal muscle results in a mosaic pattern [25–27]. Here, we found that the preferential transduction of myofiber types may explain the observed mosaic transduction by AAVs. A previous study has assessed correlation of AAV trans-duction with myofiber types using CMV-mediatedlacZ as a reporter gene. Although this study found preferential transduction of the TA muscle over soleus, correlation of reporter gene expression with MyHC isotypes was un-clear. The authors suggested that their procedure was limited by quantitative measurements [25]. We employed a quantitative histology procedure for myofiber typing by combining the antibodies to four MyHC isotypes. More-over, in contrast to previous studies that used visual assessment for myofiber types [25, 26], our assessment of myofiber types is based on measurement of the MFI of each myofiber type. This quantification of MyHC isotypes expression per myofiber enables us to apply prediction models and determine the contribution of each myofiber type to transgene expression. We found significant differ-ences between AAV6 and AAV9 for their transduction preferences to myofiber types. Myofibers expressing MyHC-2b isotype poorly express GFP that was derived by either AAV6 or AAV9. In our cassette, GFP was expressed under CMV promoter; thus, we cannot exclude the possi-bility of suboptimal activity of CMV promoter in MyHC-2b myofibers. However, previous studies showed that the CMV promoter does not discriminate between myofiber
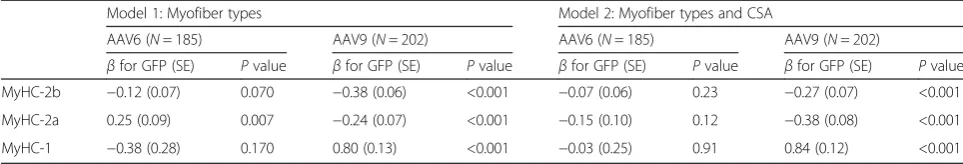
Table 1MyHC protein expression correlates with AAV9- but not with AAV6-mediated transduction
Model 1: Myofiber types Model 2: Myofiber types and CSA
AAV6 (N= 185) AAV9 (N= 202) AAV6 (N= 185) AAV9 (N= 202)
βfor GFP (SE) Pvalue βfor GFP (SE) Pvalue βfor GFP (SE) Pvalue βfor GFP (SE) Pvalue
MyHC-2b −0.12 (0.07) 0.070 −0.38 (0.06) <0.001 −0.07 (0.06) 0.23 −0.27 (0.07) <0.001
MyHC-2a 0.25 (0.09) 0.007 −0.24 (0.07) <0.001 −0.15 (0.10) 0.12 −0.38 (0.08) <0.001
MyHC-1 −0.38 (0.28) 0.170 0.80 (0.13) <0.001 −0.03 (0.25) 0.91 0.84 (0.12) <0.001
Linear regression analyses were performed including mean fluorescent intensities (MFI) for GFP as a dependent variable. In model 1, MFIs of myofibers expressing MyHC-2b, MyHC-2a, and MyHC-1 isotypes were included as independent variables. In model 2, cross-sectional area (CSA) of the myofibers was additionally included as an independent variable. Models were stratified for AAV6 and AAV9. Beta of the regression analysis and standard errors (SE) are provided
(See figure on previous page.)
Fig. 2Analysis of GFP and myofiber-type correlation in AAV6- or AAV9-transduced muscles. aImages of representative sections after immunohistochemistry with four antibodies to MyHC isotypes (staining of each MyHC isotype separately is shown in Additional file 1: Figure S5) and GFP fluorescence in a consecutive section. Examples of matching myofibers are marked witharrowheads. Scale bar in 50μm.bPlotsshow mean fluorescent intensity (MFI) distribution of MyHC isotypes and GFP within a myofiber (NAAV6= 188 fibers;NAAV9 = 202 fibers); GFP inlight green, MyHC-2b indark green, MyHC-2a inred, and MyHC-1 inblue. Myofiber types that predominantly express MyHC-2b, MyHC-2a or MyHC-2a/1, and negatively stained myofibers are schematically indicated.c Scatter plotsshow the distribution of observed GFP MFI versus predicted GFP in AAV6 (ingreen) or AAV9 (in blue) conditions. Linear regression and fitness (R2) are depicted in
types [28, 29]. Additional studies with different promoters and reporters could clarify this potential concern. We sug-gest that not all myofiber types are readily permissive for AAV. In agreement with this hypothesis, a recent study demonstrated that AAV does not transduce satellite cells, the muscle stem cells within a muscle tissue [30]. In con-trast to myofibers expressing MyHC-2b, myofibers ex-pressing MyHC-2x isotype were preferentially transduced by both AAV6 and AAV9. Analysis of MyHC-2x was ab-sent from a study that investigated the effect of AAV transduction between myofiber types [25]. Although, some receptors for AAV9 have been identified [31–33], but the molecular basis for AAV transduction and cell type speci-ficity is still poorly understood. Elucidation of AAV sero-type and myofiber specificity should be addressed in future studies, which should include screening of add-itional muscles expressing MyHC isotypes at different ra-tios and hybrid myofibers in different disease conditions. This will hopefully lead to develop more effective gene therapy strategies using AAV vectors.
Using the prediction model that includes myofiber types and CSA, we show that a correlation between GFP MFI and myofiber types was insignificant in the AAV6-transduced muscles but remained significant in the AAV9-transduced muscles. Importantly, addition of CSA to this model significantly improved the prediction of GFP MFI in AAV6- but not in AAV9-transduced mus-cles. Together, this suggests that AAV9-mediated trans-duction is predominantly determined by myofiber types
whereas AAV6 transduction is determined primarily by myofiber size. Variations in transduction efficiency be-tween AAV serotypes and different muscles are not fully understood. The TA muscle is more effectively trans-duced by AAV9 vector compared to the soleus muscle [26]. However, AAV6 is more effective for soleus trans-duction over AAV2 [25]. AAV1 transtrans-duction in dog models for muscular diseases seems to be more effective compared to mice [6]. Recently, AAV8 has also been suggested as an improved vehicle for transduction of muscle tissues [34]. Future studies should include testing of transduction efficiency of various serotypes in differ-ent muscles of multiple species, and this could result in an atlas of AAV serotypes and myofiber specificity for more effective utility of AAV-mediated gene therapy.
Conclusions
myofiber type, whereas transduction by AAV6 serotype can be explained by myofiber size, but not by myofiber type. Future studies should expand these results in muscu-lar disease models, and such studies may open a new dis-cussion to the choice of AAV serotype and cell type specificity for AAV-mediated transduction in clinical ap-plications. We suggest that understanding of AAV tropism at myofiber type levels will improve transduction efficacy and specificity for clinically interesting muscles.
Additional file
Additional file 1:A pdf file containing two supplementary tables (S1–2) and five supplementary figures (S1–5) are included as an Additional file 1. Table S1contains primer sequences used in this study.Table S2contains myofiber-based univariate correlations.Figure S1shows schematic summary of methodology.Figure S2shows GFP fluorescence in AAV-injected TA muscles.Figure S3shows GFP fluorescence in all 10 AAV- or PBS-injected mice at 4 week post-injection.Figure S4
indicates myofiber damage in AAV6-transduced TA muscles.Figure S5shows immunohistochemistry with antibodies to MyHC type-2b, MyHC type-2a, and MyHC type-1 and to laminin in AAV6- and AAV9-injected animals.
Abbreviations
AAV:adeno-associated virus; ARRIVE: Animal Research: Reporting of In Vivo Experiments; CMV: cytomegalovirus; CSA: cross-sectional area; CT: cycle of threshold; DAPI: 4′,6-diamidino-2-phenylindole; DEC: Dier Experimenten Commissie (Animal Ethical Committee); DSHB: developmental studies hybridoma bank; EGFP: enhanced green fluorescence protein; FCS: fetal calf serum; Gapdh: glyceraldehyde-3 phosphate dehydrogenase; GFP: green fluorescence protein; HEK cells: human embryonic kidney cells; Hprt: hypoxanthine phosphoribosyltransferase; ITRs: inverted terminal repeats; LAS AF: leica application suite advanced fluorescence; MD: muscular dystrophies; MyHC: myosin heavy chain; MFI: mean fluorescence intensity; PBS: phosphate-buffered solution; PBT: PBS containing 0.05 % tween; RT-qPCR: reverse transcriptase quantitative polymerase chain reaction; SE: standard error; TA: tibialis anterior; WPRE: woodchuck post-transcriptional regulatory element.
Competing interests
The authors declare that they have no competing interests.
Authors’contributions
MR, SM, VR, and JV designed the study; EBM generated AAV particles; MR and YDK performed animal experiments; YR and VR performed statistical analysis; MR, VR, and MVP performed imaging and image quantification; MR and VR wrote the manuscript. All authors have read and approved the final version of the manuscript.
Acknowledgements
This work is funded in part by the Centre for Medical Systems Biology within the framework of the Netherlands.
Author details
1
Department of Human Genetics, Leiden University Medical Center, Building 2, Room R3-17, Einthovenweg 20, 2333 ZC Leiden, The Netherlands. 2
Department of Regeneration of Sensorimotor Systems, Netherlands Institute for Neuroscience, Royal Dutch Academy of Sciences, Amsterdam, The Netherlands.3Department of Molecular and Cellular Neurobiology, Center for Neurogenomics and Cognition Research, Vrije Universiteit Amsterdam, Amsterdam, The Netherlands.
Received: 29 June 2015 Accepted: 27 October 2015
References
1. Kotterman MA, Schaffer DV. Engineering adeno-associated viruses for clinical gene therapy. Nat Rev Genet. 2014;15(7):445–51.
2. Asokan A, Schaffer DV, Samulski RJ. The AAV vector toolkit: poised at the clinical crossroads. Mol Ther. 2012;20(4):699–708.
3. Chamberlain JS. Gene therapy of muscular dystrophy. Hum Mol Genet. 2002;11(20):2355–62.
4. Zincarelli C, Soltys S, Rengo G, Rabinowitz JE. Analysis of AAV serotypes 1–9 mediated gene expression and tropism in mice after systemic injection. Mol Ther. 2008;16(6):1073–80.
5. Wang Z, Tapscott SJ, Chamberlain JS, Storb R. Immunity and AAV-mediated gene therapy for muscular dystrophies in large animal models and human trials. Front Microbiol. 2011;2:201.
6. Hakim CH, Yue Y, Shin JH, Williams RR, Zhang K, Smith BF, et al. Systemic gene transfer reveals distinctive muscle transduction profile of tyrosine mutant AAV-1, -6, and -9 in neonatal dogs. Mol Ther Methods Clin Dev. 2014;1:14002. 7. Li W, Zhang L, Wu Z, Pickles RJ, Samulski RJ. AAV-6 mediated efficient
transduction of mouse lower airways. Virology. 2011;417(2):327–33. 8. Gregorevic P, Meznarich NA, Blankinship MJ, Crawford RW, Chamberlain JS.
Fluorophore-labeled myosin-specific antibodies simplify muscle-fiber phenotyping. Muscle Nerve. 2008;37(1):104–6.
9. Schiaffino S, Reggiani C. Fiber types in mammalian skeletal muscles. Physiol Rev. 2011;91(4):1447–531.
10. Bloemberg D, Quadrilatero J. Rapid determination of myosin heavy chain expression in rat, mouse, and human skeletal muscle using multicolor immunofluorescence analysis. PLoS One. 2012;7(4), e35273.
11. Zierath JR, Hawley JA. Skeletal muscle fiber type: influence on contractile and metabolic properties. PLoS Biol. 2004;2(10), e348.
12. Ciciliot S, Rossi AC, Dyar KA, Blaauw B, Schiaffino S. Muscle type and fiber type specificity in muscle wasting. Int J Biochem Cell Biol. 2013;45(10):2191–9. 13. Wang Y, Pessin JE. Mechanisms for fiber-type specificity of skeletal muscle
atrophy. Curr Opin Clin Nutr Metab Care. 2013;16(3):243–50.
14. Alves JN, Muir EM, Andrews MR, Ward A, Michelmore N, Dasgupta D, et al. AAV vector-mediated secretion of chondroitinase provides a sensitive tracer for axonal arborisations. J Neurosci Methods. 2014;227:107–20.
15. Grimm D, Kay MA, Kleinschmidt JA. Helper virus-free, optically controllable, and two-plasmid-based production of adeno-associated virus vectors of serotypes 1 to 6. Mol Ther. 2003;7(6):839–50.
16. Gao G-P, Alvira MR, Wang L, Calcedo R, Johnston J, Wilson JM. Novel adeno-associated viruses from rhesus monkeys as vectors for human gene therapy. Proc Natl Acad Sci U S A. 2002;99(18):11854–9.
17. Hermens WT, ter Brake O, Dijkhuizen PA, Sonnemans MA, Grimm D, Kleinschmidt JA, et al. Purification of recombinant adeno-associated virus by iodixanol gradient ultracentrifugation allows rapid and reproducible preparation of vector stocks for gene transfer in the nervous system. Hum Gene Ther. 1999;10(11):1885–91.
18. Kilkenny C, Browne WJ, Cuthill IC, Emerson M, Altman DG. Improving bioscience research reporting: the ARRIVE guidelines for reporting animal research. PLoS Biol. 2010;8(6), e1000412.
19. Histopathology in hematoxylin & eosin stained muscle sections. [http:// www.treat-nmd.eu/downloads/file/sops/cmd/MDC1A_M.1.2.004.pdf]. Accessed on 14-04-2015.
20. Schneider CA, Rasband WS, Eliceiri KW. NIH Image to ImageJ: 25 years of image analysis. Nat Methods. 2012;9(7):671–5.
21. Trollet C, Anvar SY, Venema A, Hargreaves IP, Foster K, Vignaud A, et al. Molecular and phenotypic characterization of a mouse model of oculopharyngeal muscular dystrophy reveals severe muscular atrophy restricted to fast glycolytic fibres. Hum Mol Genet. 2010;19(11):2191–207. 22. Arnett AL, Garikipati D, Wang Z, Tapscott S, Chamberlain JS. Immune
responses to rAAV6: the influence of canine parvovirus vaccination and neonatal administration of viral vector. Front Microbiol. 2011;2:220. 23. Mingozzi F, High KA. Therapeutic in vivo gene transfer for genetic disease
using AAV: progress and challenges. Nat Rev Genet. 2011;12(5):341–55. 24. Hastie E, Samulski RJ. AAV at 50: a golden anniversary of discovery, research,
and gene therapy success, a personal perspective. Hum Gene Ther. 2015. 25. Blankinship MJ, Gregorevic P, Allen JM, Harper SQ, Harper H, Halbert CL, et
al. Efficient transduction of skeletal muscle using vectors based on adeno-associated virus serotype 6. Mol Ther. 2004;10(4):671–8.
27. Bortolanza S, Nonis A, Sanvito F, Maciotta S, Sitia G, Wei J, et al. AAV6-mediated systemic shRNA delivery reverses disease in a mouse model of
facioscapulohumeral muscular dystrophy. Mol Ther. 2011;19(11):2055–64. 28. Hallauer PL, Hastings KE. Human cytomegalovirus IE1 promoter/enhancer
drives variable gene expression in all fiber types in transgenic mouse skeletal muscle. BMC Genet. 2000;1:1.
29. Robinson DA, Bremner SN, Sethi K, Shah SB, Sirsi SR, Lutz GJ. In vivo expression of myosin essential light chain using plasmid expression vectors in regenerating frog skeletal muscle. Gene Ther. 2005;12(4):347–57. 30. Arnett AL, Konieczny P, Ramos JN, Hall J, Odom G, Yablonka-Reuveni Z, et
al. Adeno-associated viral (AAV) vectors do not efficiently target muscle satellite cells. Mol Ther Methods Clin Dev. 2014;1:1-10.
31. Akache B, Grimm D, Pandey K, Yant SR, Xu H, Kay MA. The 37/67-kilodalton laminin receptor is a receptor for adeno-associated virus serotypes 8, 2, 3, and 9. J Virol. 2006;80(19):9831–6.
32. Wu Z, Miller E, Agbandje-McKenna M, Samulski RJ. Alpha2,3 and alpha2,6 N-linked sialic acids facilitate efficient binding and transduction by adeno-associated virus types 1 and 6. J Virol. 2006;80(18):9093–103. 33. Shen S, Bryant KD, Brown SM, Randell SH, Asokan A. Terminal N-linked
galactose is the primary receptor for adeno-associated virus 9. J Biol Chem. 2011;286(15):13532–40.
34. Charan RA, Niizawa G, Nakai H, Clemens PR. Adeno-associated virus serotype 8 (AAV8) delivery of recombinant A20 to skeletal muscle reduces pathological activation of nuclear factor (NF)-kappaB in muscle of mdx mice. Mol Med. 2012;18:1527–35.
Submit your next manuscript to BioMed Central and take full advantage of:
• Convenient online submission
• Thorough peer review
• No space constraints or color figure charges
• Immediate publication on acceptance
• Inclusion in PubMed, CAS, Scopus and Google Scholar
• Research which is freely available for redistribution