Chlamydia felis
Genotyping: Comparison with Multilocus Sequence
Typing
Karine Laroucau,aAntonietta Di Francesco,bFabien Vorimore,aSimon Thierry,aJean Luc Pingret,cClaire Bertin,aHermann Willems,d Goran Bölske,eand Ross Harleyf
Bacterial Zoonoses Unit, French Agency for Food, Environmental & Occupational Health Safety (ANSES), Maisons-Alfort, Francea; Department of Veterinary Medical Sciences, University of Bologna, Bologna, Italyb; Scanelis, Colomiers, Francec; Clinic for Swine, Giessen, Germanyd; Department of Bacteriology, National Veterinary Institute (SVA), Uppsala, Swedene; and School of Clinical Veterinary Sciences, University of Bristol, Bristol, United Kingdomf
Chlamydia felisis an important ocular pathogen in cats worldwide. A multilocus variable-number tandem-repeat analysis (MLVA) system for the detection of tandem repeats across the whole genome ofC.felisstrain Fe/C-56 was developed. Nine se-lected genetic loci were tested by MLVA in 17C.felisisolates, including theC.felisBaker vaccine strain, and 122 clinical samples from different geographic origins. Analysis of the results identified 25 distinctC.felisMLVA patterns. In parallel, a recently de-scribed multilocus sequence typing scheme for the typing ofChlamydiawas applied to 13 clinical samples with 12 differentC.
felisMLVA patterns. Rare sequence differences were observed. Thus, the newly developed MLVA system provides a highly sensi-tive high-resolution test for the differentiation ofC.felisisolates from different origins that is suitable for molecular epidemio-logical studies.
C
hlamydiaceae spp. are obligate intracellular Gram-negativebacterial parasites that have a worldwide distribution and cause various diseases in animals, including humans. According to the most recent taxonomy, the familyChlamydiaceaewith its sin-gle genus,Chlamydia, contains nine species (18). Among them,
Chlamydia felisis an important pathogen of cats (10).
Conjuncti-val epithelium appears to be the major target ofC.felis, although the microorganism has been detected in a variety of tissues and anatomical locations in cats (reviewed in reference36).C. felis
causes acute-to-chronic conjunctivitis, particularly in young cats, and is the most common cause of conjunctivitis in cats, detected in approximately 30% of clinical cases (7,11,28,34,35,41). Feline chlamydiosis is typically characterized by unilateral or bilateral acute conjunctivitis and excretion of high levels ofC.felis, peaking approximately 1 to 2 weeks postinfection (5,23,37,40). Thereaf-ter, chronic conjunctivitis and lower levels ofC.felis excretion often persist for up to about 2 months postinfection prior to the resolution of clinical signs and cessation of shedding (23,40). However, some cats may remain persistently infected for longer time periods and may represent a population of asymptomatic carriers (23,36,40). Among other clinical signs,C.felisinfection has been associated with vaginal discharge and excretion in exper-imentally infected cats and the organism has been suspected as a cause of abortion and reproductive disease in queens; however, the limited evidence for this is largely circumstantial (36).
Chlamydiosis is relatively common in the domestic cat popu-lation, and the infection may become endemic, especially in cat colonies (6). Transmission ofC.felisbetween cats occurs primar-ily by direct contact with infective ocular secretions. Rescue shel-ters may therefore represent high-risk environments for the trans-mission of chlamydiosis, as a result of large numbers of animals being housed in limited spaces and the periodic introduction of new, susceptible cats, particularly kittens.
The zoonotic potential of this bacterium appears to be low; however, exposure toC.felismay occur when handling infected
cats. Hence, cat owners and professionals who work with cats (veterinarians, employees at catteries, and cat breeders) are at in-creased risk, especially where there are insufficient hygienic con-ditions. The risk of zoonotic transmission is presumably greater for immunocompromised individuals.C.felisinfection has been associated with conjunctivitis and/or respiratory tract disease (4,
15), community-acquired pneumonia (21), hepatosplenomegaly, glomerulonephritis, and endocarditis in humans (31).
Until now, genotyping studies conducted withC. felis have shown low genetic diversity among isolates of this species, with a substantial degree of rRNA andompAgene conservation (29,33). The 16S rRNA genes ofC.felisstrains that have been sequenced to date differ by less than 0.6% (8). However, heterogeneity of feline strains related to geographic origin was demonstrated by restric-tion fragment length polymorphism of thegroELgene (9). Also, analysis by random amplification of polymorphic DNA revealed at least two different genetic fingerprints among the sixC. felis
strains examined (30).
With the increasing availability of whole bacterial genome se-quences, new molecular tools based on genome-wide screens are currently being developed. The typing method termed multiple-locus variable-number tandem-repeat (VNTR) analysis (MLVA) is based on the detection of tandem-repeat polymorphisms (38) and has been used successfully for the typing of many pathogens, includingChlamydia trachomatis (27),Chlamydia psittaci(19),
andChlamydia abortus(20). The typing method termed
multilo-Received14 February 2012 Returned for modification7 March 2012 Accepted2 April 2012
Published ahead of print11 April 2012
Address correspondence to Karine Laroucau, [email protected].
Copyright © 2012, American Society for Microbiology. All Rights Reserved.
doi:10.1128/JCM.00417-12
on May 16, 2020 by guest
http://jcm.asm.org/
cus sequence typing (MLST) is often based on the sequence anal-ysis of internal fragments of seven housekeeping genes. A previ-ously described MLST scheme has been successfully used to understand the population genetic structures ofC.trachomatis,C.
pneumonia, andC.psittaci(24,25).
The aims of the present study were to develop a novel MLVA scheme specific forC.felisgenotyping and to compare it with the previously described MLST typing method.
MATERIALS AND METHODS
Bacterial isolates and clinical samples.DNA extracted from 122 clinical samples (121 conjunctival swabs and 1 placental swab) and 17C.felis
isolates (16 field isolates and the Baker vaccine strain) (Table 1) were obtained from ANSES (Maisons-Alfort, France), Scanelis (Colomiers, France), the Department of Veterinary Medical Sciences, University of Bologna (Ozzano dell’ Emilia, Italy), the Clinic for Swine (Giessen, Ger-many), the National Veterinary Institute of Sweden (Uppsala, Sweden), and the University of Bristol (Bristol, United Kingdom). Samples were collected from both eyes of some Italian cats and analyzed.
Italian clinical samples were collected in 2000 and 2001 at three differ-ent catteries (6). German, Swedish, and United Kingdom (13) samples were collected at various times from 2005 to 2007, and French samples were collected in 2011 as part of routine diagnostic analysis.
Both clinical samples andC.felisisolates had been previously subjected to DNA extraction, and all of the DNA included in this study tested positive with theC.felis-specific real-time PCR assay described by Pantchev et al. (26). The protocol included primers CpfOMP1-F (5=-TCGGATTGATTGGTCTTGC A-3=) and CpfOMP1-R (5=-GCTCTACAATGCCTTGAGAAATTTC-3=) and probe CpfOMP1-S (6-carboxyfluorescein–5=-ACTGATTTCGCCA ATCAGCGTCCAA-3=– 6-carboxytetramethylrhodamine). Each reaction mixture (20l) contained 2l of sample template DNA, 10l of 2⫻ Universal Master Mix (Applied Biosystems), a 0.625M final concentra-tion of each primer, a 0.1M final concentration of the probe, and 5l of deionized water. The temperature-time profile was 95°C for 10 min, fol-lowed by 45 cycles at 95°C for 15 s and 60°C for 60 s.
MLVA genotyping method.Tandem-repeat sequences within the whole genome sequence ofC.felisstrain Fe/C-56 (GenBank accession no.
AP006861) (1) were determined using the Tandem Repeats Finder pro-gram (http://tandem.bu.edu/trf/trf.html) (3). Nine tandem repeats with a repeat unit length ofⱖ12 bp were identified, and primers for PCR ampli-fication on both sides of the repeats were chosen (Table 2). VNTR size was determined in order to allow efficient differentiation on agarose gel.
All 17 isolates (including the Baker vaccine strain) and 122 clinical samples were subjected to MLVA. For VNTR amplification, a PCR was performed in a total volume of 15l containing 5 to 10 ng of DNA, 1⫻ PCR buffer, 1 U of HotStarTaq DNA polymerase (Qiagen), a 200M final concentration of each deoxynucleotide, and a 0.3M final concentration of each flanking primer. The initial denaturation step of 95°C for 15 min was followed by 40 cycles consisting of denaturation at 95°C for 30 s, primer annealing at 58°C for 30 s, and elongation at 72°C for 45 s. The final extension step was 72°C for 10 min. Five microliters of amplification product was loaded onto a 3% agarose gel. Gels stained with ethidium bromide were visualized under UV light and photographed. The size marker used was a 100-bp ladder (MBI-Fermentas–Euromedex, Souffel-weyersheim, France).
Amplicon size was determined using the Quantity One software pack-age version 4.6.7 (Bio-Rad Laboratories, Inc., Hercules, CA). The number of motifs in each allele was derived from the amplicon size. For each discriminant VNTR marker, representative patterns were sequenced (MWG Biotech France, Roissy, France).
MLST genotyping method.Based on previous work (25), we ampli-fied and sequenced seven housekeeping genes (enoA,fumC,gatA,gidA,
hemN,hflX, andoppA) from 13 randomly selected French clinicalC.felis
samples (2, 3, 4, 5, 6, 8, 9,
11E3-10, 11E3-11, 11E3-13, 11E3-19, 11E3-22, 11E3-38) (Table 1, samples in bold) and from theC.felisBaker vaccine strain.
Cell culture.For cell culture, yolk sacs of 7-day-old embryonated eggs were inoculated with 0.2 ml of theC.felissuspension per egg and five eggs were used per sample. For each inoculation set, three eggs were inoculated
withC.psittacistrain Loth as a positive control and three other eggs were
kept separately as noninfected controls. Eggs were incubated at 38°C and observed daily. Vitellus membranes were collected from dead embryos and submitted to 10 serial passages in chicken eggs. DNA was extracted from collected vitellus membranes and analyzed by PCR and then MLVA as described above.
Nucleotide sequence accession numbers.The sequences determined by MLST have been deposited in the GenBank database under accession numbersHE795868toHE795965, and those determined by MLVA have been deposited under accession numbersHE795966toHE795983.
RESULTS
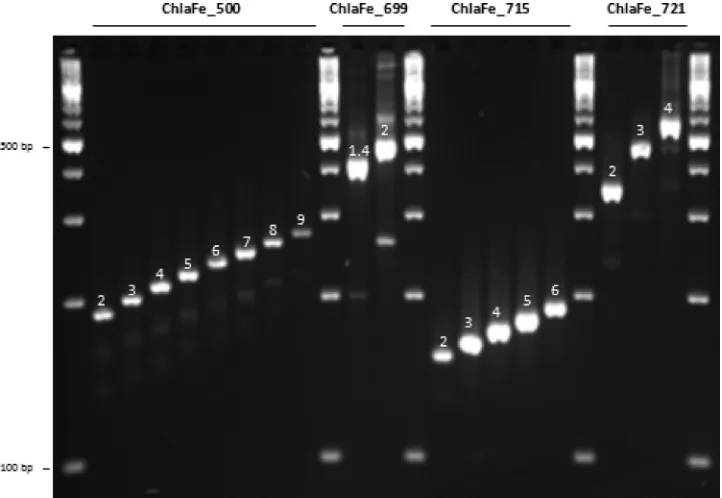
Selection of VNTRs for MLVA.Fragment size analysis of the am-plified PCR products showed that four of the tandem repeats, namely, ChlaFe_500, ChlaFe_699, ChlaFe_715, and ChlaFe_721, were polymorphic, with eight, two, five, and three different pat-terns, respectively (Fig. 1; Table 1). Based on these four VNTR markers,C.felisisolates and/or samples were subdivided into 25 genotypes termed genotypes 1 through 25 (Table 1). Expected tandem-repeat sizes were observed, except for the marker ChlaFe_715, for which a 9-bp tandem-repeat unit was discovered rather than the 18-bp repeat identified by the software, which was set to detect tandem-repeat units with a minimum size of 12 bp. No geographic link between isolates or samples from different European countries was found (data not shown).
The stability of the four discriminant markers was assessed by comparison of the MLVA patterns of two strains (the Baker vac-cine strain and the 99-9537 isolate) after 10 serial passages in chicken eggs. MLVA typing results were identical to those ob-tained initially (data not shown).
Focus on samples collected in three catteries.Clinical samples from four, six, and nine cats belonging to three different Italian catteries were investigated. Samples were harvested during an ep-idemiological study performed in 2000 and 2001 (6). Most of the cats belonging to the catteries sampled had conjunctivitis for sev-eral months, and all of the clinical samples analyzed in the present study were from cats showing ophthalmic problems at the time of sampling.
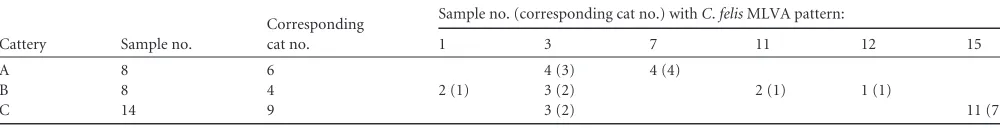
Up to four different genotypes were identified within the same cattery. In particular, in cattery A, genotypes 3 and 7 were detected, whereas genotypes 1, 3, 11, and 12 were detected in cattery B and genotypes 3 and 15 were identified in cattery C (Tables 1and3).
In cattery A, two cats harbored genotype 3, three cats had ge-notype 7, and one cat had both gege-notypes 3 and 7. In cattery B, one cat harbored genotype 1, one cat had genotype 3, one cat had genotype 11, and one cat had both genotypes 3 and 12. In cattery C, two cats harbored genotype 3 and seven cats had genotype 15. From a clinical sample from cattery A, one isolate (M) with MLVA genotype 3 was obtained. This is consistent with the find-ing of clinical samples with the same MLVA pattern from this cattery (Table 3).
Of the cats from both of whose eyes samples were collected, nine had the same genotype in both samples. In contrast, in two cats from two different catteries, a different genotype was identi-fied in each eye. One of these cats (number 2), in cattery A, had
on May 16, 2020 by guest
http://jcm.asm.org/
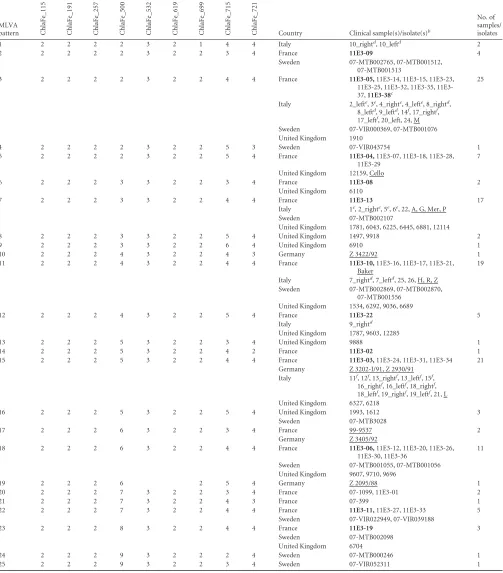
TABLE 1C.felisisolates and samples organized by MLVA genotypea
MLVA pattern
No. of different patterns
Country Clinical sample(s)/isolate(s)b
No. of samples/ isolates ChlaFe_115 ChlaFe_191 ChlaFe_257 ChlaFe_500 ChlaFe_532 ChlaFe_619 ChlaFe_699 ChlaFe_715 ChlaFe_721
1 2 2 2 2 3 2 1 4 4 Italy 10_rightd
, 10_leftd
2
2 2 2 2 2 3 2 2 3 4 France 11E3-09 4
Sweden 07-MTB002765, 07-MTB001512,
07-MTB001513
3 2 2 2 2 3 2 2 4 4 France 11E3-05,11E3-14, 11E3-15, 11E3-23,
25, 32, 35, 11E3-37,11E3-38c
25
Italy 2_lefte
, 3e
, 4_righte
, 4_lefte
, 8_rightd
, 8_leftd
, 9_leftd
, 14f
, 17_rightf
, 17_leftf
, 20_left, 24, M
Sweden 07-VIR000369, 07-MTB001076
United Kingdom 1910
4 2 2 2 2 3 2 2 5 3 Sweden 07-VIR043754 1
5 2 2 2 2 3 2 2 5 4 France 11E3-04,11E3-07, 11E3-18, 11E3-28,
11E3-29
7
United Kingdom 12159, Cello
6 2 2 2 3 3 2 2 3 4 France 11E3-08 2
United Kingdom 6110
7 2 2 2 3 3 2 2 4 4 France 11E3-13 17
Italy 1e, 2_righte, 5e, 6e, 22, A, G, Mer, P
Sweden 07-MTB002107
United Kingdom 1781, 6043, 6225, 6445, 6881, 12114
8 2 2 2 3 3 2 2 5 4 United Kingdom 1497, 9918 2
9 2 2 2 3 3 2 2 6 4 United Kingdom 6910 1
10 2 2 2 4 3 2 2 4 3 Germany Z 3422/92 1
11 2 2 2 4 3 2 2 4 4 France 11E3-10,11E3-16, 11E3-17, 11E3-21,
Baker
19
Italy 7_rightd
, 7_leftd
, 25, 26, H, R, Z
Sweden 07-MTB002869, 07-MTB002870,
07-MTB001556 United Kingdom 1534, 6292, 9036, 6689
12 2 2 2 4 3 2 2 5 4 France 11E3-22 5
Italy 9_rightd
United Kingdom 1787, 9603, 12285
13 2 2 2 5 3 2 2 3 4 United Kingdom 9888 1
14 2 2 2 5 3 2 2 4 2 France 11E3-02 1
15 2 2 2 5 3 2 2 4 4 France 11E3-03,11E3-24, 11E3-31, 11E3-34 21
Germany Z 3202-I/91, Z 2930/91
Italy 11f
, 12f
, 13_rightf
, 13_leftf
, 15f
, 16_rightf
, 16_leftf
, 18_rightf
, 18_leftf
, 19_rightf
, 19_leftf
, 21, L
United Kingdom 6327, 6218
16 2 2 2 5 3 2 2 5 4 United Kingdom 1993, 1612 3
Sweden 07-MTB3028
17 2 2 2 6 3 2 2 3 4 France 99-9537 2
Germany Z 3405/92
18 2 2 2 6 3 2 2 4 4 France 11E3-06,11E3-12, 11E3-20, 11E3-26,
11E3-30, 11E3-36
11
Sweden 07-MTB001055, 07-MTB001056
United Kingdom 9607, 9710, 9696
19 2 2 2 6 2 5 4 Germany Z 2095/88 1
20 2 2 2 7 3 2 2 3 4 France 07-1099, 11E3-01 2
21 2 2 2 7 3 2 2 4 3 France 07-399 1
22 2 2 2 7 3 2 2 4 4 France 11E3-11,11E3-27, 11E3-33 5
Sweden 07-VIR022949, 07-VIR039188
23 2 2 2 8 3 2 2 4 4 France 11E3-19 3
Sweden 07-MTB002098
United Kingdom 6704
24 2 2 2 9 3 2 2 2 4 Sweden 07-MTB000246 1
25 2 2 2 9 3 2 2 3 4 Sweden 07-VIR052311 1
aGenotypes were based on similarity clustering of PCR product sizes obtained with the nine primer sets used. The genotypes are numbered 1 through 25. b
Isolates and the vaccine strain are underlined, and samples analyzed by MLST are in bold.
cPlacental swab (11E3-38). d
From cattery B.
eFrom cattery A. f
From cattery C.
on May 16, 2020 by guest
http://jcm.asm.org/
genotype 3 in its left eye and genotype 7 in its right eye. These two genotypes, which differ by only one tandem repeat in marker ChlaFe_500, were also detected in samples from other cats in this cattery: three cats harbored genotype 3, and four cats harbored genotype 7. In this cattery, the same genotype was detected in both eyes of one cat (number 4, genotype 3).
A different genotype was also detected in each eye of cat num-ber 9 (genotypes 3 and 12). Interestingly, whereas genotype 3 was identified in other cats in this cattery, genotype 12 was detected only in cat number 9. Genotypes 3 and 12 differ in two markers (ChlaFe_500 and ChlaFe_715), with two and one tandem-repeat differences, respectively.
Sequence analysis of PCR products.PCR products generated for each of the four polymorphic markers (Fig. 1) were sequenced. Sequence analysis confirmed the fragment size analysis of the PCR products. Locus ChlaFe_500 is localized in a noncoding sequence.
[image:4.585.40.546.77.199.2]As expected, DNA sequencing of representatives of the eight dif-ferent patterns identified difdif-ferent numbers of tandem repeats consisting of a 12-bp sequence (CCAACCTAGGAA) (frequencies of tandem repeats in ChlaFe_500: two tandem repeats, 28.1% of the samples analyzed; three tandem repeats, 15.8%; four tandem repeats, 18.0%; five tandem repeats, 19.4%; six tandem repeats, 10.1%; seven tandem repeats, 5.8%; eight tandem repeats, 2.2%; nine tandem repeats, 1.4%). All of the other markers are located in a coding region or in a hypothetical coding region. Locus ChlaFe_715 is located in a hypothetical-protein-encoding gene (CF0611) where the 9-mer GAAGCAGAA coding for the peptide EAE is repeated two to six times. In most (76.3%) of the samples analyzed, this 9-mer was repeated four times. Notably, CF0611 has no homology with other chlamydial proteins described to date. A particular pattern was observed for the locus ChlaFe_699, which was detected only in clinical samples collected from both TABLE 2Primers for PCR amplification and MLVAa
VNTR primer
Sequence (5=–3=)
Tm
(°C)
Repeat unit length (bp)
C.felis
Fe/C-56 theoretic repeat no.
Size ofC.felis
Fe/C-56 amplicon (bp)
Forward Reverse
ChlaFe_115 TCATCAGCGATGAATTATTCAGAATAG CAATGCCGCCGTAACATTT 58 23 1.9 220
ChlaFe_191 CCTCCCTGAAGAAGCACTGTTT ATTTTACGCGAACAACGTTATCC 58 21 2.0 215
ChlaFe_257 TTTTAGAGAAAGATGCGGTAATGAAAG TCCATGAGGTTGTGATTGCAA 59 21 1.9 143
ChlaFe_500 CGAGGTTCTCTCAATAATGCTCTAAACT TTCTTCTCATCGGCACCAAAC 59 12 4.0 213
ChlaFe_532 GGCCCCCTCGCAAGG CCCCACAACCCGACCA 58 17 2.6 160
ChlaFe_619 TTCTGAGTTCCCTTCAATAGCTTTTC TTTGCGCGTGTTGGCAT 59 18 2.2 167
ChlaFe_699 AAGTAGACAGTGCTGGTTTTGCTACT ACAAAAGAAACGTAGCAGCAAGA 57 30 2.3 470
ChlaFe_715 AAAATTGACACCAGCACCACC GGTAGAGTCGTTAGTAATCTCGTGGTAG 58 18/9 2.0/4.0 171
ChlaFe_721 AGAGATTGTAATGCCTGTGGGTTC TCCCCATTGGCTCCTGTG 59 111 4.0 572
a
The primers were used to characterize tandem repeats at nine genetic loci.
FIG 1C.felisMLVA patterns observed with different VNTR polymorphic markers. ChlaFe_500 showed eight patterns (2 [Cello strain], 3 [isolate A], 4 [Baker strain], 5 [isolate L], 6 [sample 07-MTB001055], 7 [sample 07-VIR022949], 8 [sample 07-MTB002098], and 9 [sample 07-MTB000246]), ChlaFe_699 showed two patterns (1.4 [sample 10_right] and 2 [sample 9607]), ChlaFe_715 showed five patterns (2 [sample 07-MTB000246], 3 [sample 07-1059], 4 [Baker strain], 5 [sample 07-VIRO43754], and 6 [sample 6910]), and ChlaFe_721 showed three patterns (2 [sample 11E3-02], 3 [sample 07-VIRO43754], and 4 [Cello strain]).
on May 16, 2020 by guest
http://jcm.asm.org/
[image:4.585.112.473.434.683.2]eyes of a single cat (the same pattern in samples from both eyes). DNA sequence analysis of PCR products revealed a 60-bp deletion outside the tandem-repeat sequence region. The deleted sequence which corresponds to the peptide VVFLTVLRAVVFLATVLRTV is part of the histone H1-like protein Hc2. Sequencing of the locus ChlaFe_721 revealed the presence of two different tandem-repeat sequences in this amplified fragment. These two tandem-repeat sequences are each repeated two times inC.felisreference strain Fe/C-56. Including the reference strain pattern, three new pat-terns were identified, one with one deletion in the first tandem-repeat sequence and another with one deletion in each of the tan-dem-repeat sequences. The deletions are contained in a hypothetical-protein-encoding gene (frequencies of tandem re-peats in ChlaFe_721: two tandem rere-peats, 0.7%; three tandem repeats, 2.2%; four tandem repeats, 97.1%).
MLST. The clinical samples selected from large (multitudi-nous) MLVAC.felispatterns 2, 3, 5, 6, 7, 11, 12, 14, 15, 18, 22, and 23 were successfully amplified by the MLST scheme. Identical se-quences (ST_21) were identified within this clinical sample panel for the seven targeted housekeeping genes, except for one point mutation in thehflXgene in sample 11E3-4. In addition, two nonsynonymous mutations were detected in theenoAgene of the
C.felisBaker vaccine strain.
DISCUSSION
Previous studies based on 16S rRNA,ompA, orgroELgene se-quences (8,9,29,33) have highlighted the remarkable homoge-neity ofC.felisstrains. Here we present an MLVA-based molecu-lar typing system for the discrimination ofC.felisisolates that does not require culture and can therefore be performed directly with DNA extracted from clinical specimens. MLVA typing is a repro-ducible method which can be standardized and performed at low cost, thus facilitating large-scale molecular epidemiological inves-tigations. Out of nine tandem-repeat loci identified in theC.felis
Fe/C-56 genome by the program Tandem Repeat Finder, four were polymorphic. The analysis of about 140C.felisclinical sam-ples and isolates resulted in a total of 25 distinct patterns desig-nated genotypes 1 through 25. High genotypic diversity was dem-onstrated in samples from each of the European countries of origin; however, four genotypes, 3, 7, 11, and 15, appear more widespread both in the samples tested and in the countries of origin. As transmission ofC.felisrelies on close contact between infected and susceptible cats, significant international spread ofC.
felisis unlikely. The discovery of broadly comparableC.felis
ge-notypic profiles in each country suggests thatC.felispopulations in different geographic locations undergo similar evolutionary di-vergence patterns in VNTR loci. Notably, variation arising in the most polymorphic VNTR marker, ChlaFe_500, is responsible for the majority of the genotypic differences, particularly between dominant genotypes. Thus, the potential for homoplasy may be
relatively high, which could have an impact on the discriminatory power ofC.felisgenotyping by MLVA in molecular epidemiology studies (32).
Interestingly, the Baker vaccine strain, which was first isolated from cats with pneumonia (2), was classified as genotype 11. Therefore, MLVA could provide the means to differentiate be-tween vaccinated and naturally infected animals if local strains were of a genotype different from that of the locally available vac-cine strains. Isolates and/or samples with genotype 11 were ob-tained from multiple countries; however, no information is avail-able concerning the vaccination history of the sampled animals included in this study. Thus, it remains undetermined if these field isolates represent the vaccine strain or circulating field strains with a similar MLVA genotype.
Two of the VNTR markers identified in the present study (ChlaFe_715 and ChlaFe_721) were located within open reading frames encoding putative proteins (CF0611 and CF0618, respec-tively). A further marker (ChlaFe_699) was located in hctB
(CF0595), which encodes a histone-like protein involved in the establishment of the nucleoid structure of chlamydial elementary bodies. Of the five polymorphic VNTR markers forC.abortus, one of them was located within this same family of genes (20). InC.
trachomatis, Hc2 (encoded byhctB) varies in size between strains
due to internal deletions encoding lysine- and alanine-rich penta-meric repeats (12). Different numbers of repetitive elements and point mutations were recently detected in this gene in genital clin-ical samples, leading to high-resolution intraserotype variation in
C.trachomatis(17). In the present study,hctBdiffered in only one
clinical sample with a 60-nucleotide deletion corresponding to a 20-mer peptide. Polymorphic sites, including those withinhctB, were shown to be responsible for the pathogenic diversity ofC.
trachomatistrachoma strains (16). The biological significance of
the coding tandem repeats identified in two putative proteins (CF0611 and CF0618) and the deletion in thehctBgene (CF0595) is unknown. Coding tandem repeats are believed to enhance spe-cific properties of bacterial virulence factors. Potential differences in pathogenicity between strains/isolates ofC.felisremain largely uncharacterized, and few studies have investigated virulence or genotypic differences (22,14). MLVA genotyping may, however, provide a useful molecular method for identifying strains from different lineages for investigation of phenotypic diversity and differences in clinical behavior. In the present study, it was not possible to determine ifC.felisisolates with different MLVA ge-notypes might be associated with variations in disease pathogen-esis or clinical outcomes, as detailed clinical information from individual cases was not available, except for cats observed within the Italian catteries. In the latter cases, cats infected with different
C. felisMLVA genotypes, including the cat bearing theC. felis
[image:5.585.41.541.78.142.2]isolate withhctB deleted, showed broadly similar clinical signs typical ofC.felisinfection.
TABLE 3MLVA patterns ofC.felisstrain samples from three Italian catteries where conjunctivitis was reporteda
Cattery Sample no.
Corresponding cat no.
Sample no. (corresponding cat no.) withC.felisMLVA pattern:
1 3 7 11 12 15
A 8 6 4 (3) 4 (4)
B 8 4 2 (1) 3 (2) 2 (1) 1 (1)
C 14 9 3 (2) 11 (7)
aBoth eyes of some cats were swabbed.
on May 16, 2020 by guest
http://jcm.asm.org/
Investigations conducted in catteries revealed the diversity of
C. felis strains that can be harbored by cats within a confined
environment. Interestingly, in samples from two cats in two dif-ferent catteries, where both eyes were sampled, two difdif-ferent ge-notypes were identified, which in one case differed by only one tandem repeat. These findings provide the first evidence that mixed infections with strains of differentC.felisgenotypes occur in cats, similar to those described forC.trachomatisin humans (39). The possibility that the different genotypes identified in these two cases may represent divergent populations resulting from instability in the MLVA loci cannot be excluded; however, analysis of twoC.felisisolates passaged 10 times through chicken eggs suggests that the MLVA genotype is highly stable. Neverthe-less suchin vitromethods may not adequately reflectin vivo selec-tive pressures from factors such as host immune responses. Future experiments analyzing the stability of MLVA genotypes during the course ofC.felisinfection and antibiotic treatment and studies of
C.felissuperinfection with two or more MLVA genotypes in
indi-vidual cats and in cat communities would be of considerable in-terest.
MLST based on the partial sequences of seven housekeeping genes was recently used to understand the population genetic structure ofC.trachomatis,C.pneumoniae(24), andC.psittaci
(25). Results of cluster analyses of concatenated sequences of the seven housekeeping genes validated the proposed typing system for all chlamydial species. In our study, 12 conjunctival samples with 12 differentC.felisMLVA patterns, as well as a unique sample collected from abortive feline placental tissue, and the Baker vac-cine strain were submitted to the MLST scheme. Sequences were identical, with only two mutations detected in theenoAgene of the Baker vaccine strain and one point mutation detected in thehflX
gene in a clinical sample. Almost all of the clinical samples were of the same sequence type (ST_21) as theC.felisFe/C-56 reference strain (http://pubmlst.org/chlamydiales/), highlighting the strong conservation of these genes inC.felis, except for one mutation identified only in the Fe/C-56 hemNgene. MLVA is based on differences in the numbers of tandem repeats at multiple loci on the chromosome of bacteria, whereas MLST is based on the se-quence analysis of housekeeping genes. In this case, the discrimi-natory power of MLVA was superior to that of MLST. No specific pattern was identified by either MLVA or MLST in the singleC.
felisabortion tissue sample examined or in the conjunctival
sam-ples collected. However, before any formal conclusion can be drawn, it is necessary to analyze a larger cohort ofC.felissamples recovered from cases of feline abortion. Interestingly, by MLST, the Baker strain could be differentiated from others strains by two nonsynonymous mutations in theenoAgene; however, the bio-logical significance of these mutations remains unknown.
In summary, our findings show that MLVA is a novel genotyp-ing method providgenotyp-ing enhanced discrimination ofC.felisstrains. Preliminary work conducted with clinical samples showed that MLVA is more discriminant than MLST forC.felisgenotyping.
ACKNOWLEDGMENT
We are grateful to Christine Pourcel for her invaluable help with the MLVA design.
REFERENCES
1.Azuma Y, et al.2006. Genome sequence of the cat pathogen, Chlamydo-phila felis. DNA Res.13(1):15–23.
2.Baker JA.1942. A virus obtained from a pneumonia of cats and its possi-ble relation to the cause of atypical pneumonia in man. Science96:475– 476.
3.Benson G.1999. Tandem repeats finder: a program to analyze DNA se-quences. Nucleic Acids Res.27:573–580.
4.Corsaro D, Venditti D, Valassina M.2002. New parachlamydial 16S rDNA phylotypes detected in human clinical samples. Res. Microbiol. 153(9):563–567.
5.Dean R, Harley R, Helps C, Caney S, Gruffydd-Jones T.2005. Use of quantitative real-time PCR to monitor the response ofChlamydophila felis
infection to doxycycline treatment. J. Clin. Microbiol.43:1858 –1864. 6.di Francesco A, Carelle MS, Baldelli R. 2003. Feline chlamydiosis in
Italian stray cat homes. Vet. Rec.153(8):244 –245.
7.di Francesco A, Donati M, Battelli G, Cevenini R, Baldelli R. 2004. Seroepidemiological survey forChlamydophila felisamong household and feral cats in northern Italy. Vet. Rec.155(13):399 – 400.
8.Everett KD, Bush RM, Andersen AA.1999. Emended description of the orderChlamydiales, proposal ofParachlamydiaceaefam. nov. and
Simka-niaceaefam. nov., each containing one monotypic genus, revised
taxon-omy of the familyChlamydiaceae, including a new genus and five new species, and standards for the identification of organisms. Int. J. Syst. Bacteriol.49:415– 440.
9.Fukushi H, Hirai K.1994. Heterogeneity and homogeneity ofompA(the outer membrane protein) andgroEL-homolog genes of avian and mam-malianChlamydia psittaciby PCR-based RFLP analysis, p 589 –592.In
Orifa J, et al. (ed), Eighth International Symposium on Human Chlamyd-ial Infection. Societa` Editrice Esculapio, Bologna, Italy.
10. Gruffydd-Jones T, et al. 2009. Chlamydophila felis infection. ABCD guidelines on prevention and management. J. Feline Med. Surg.11:605– 609.
11. Gunn-Moore DA, Werrett G, Harbour DA, Feilden H, Gruffydd-Jones TJ.1995. Prevalence ofChlamydia psittaciantibodies in healthy pet cats in Britain. Vet. Rec.136:366 –367.
12. Hackstadt T, Brickman TJ, Barry CE III, Sager J.1993. Diversity in the
Chlamydia trachomatishistone homologue Hc2. Gene132(1):137–141.
13. Harley R, et al.2007. Molecular characterisation of twelveChlamydophila
felispolymorphic membrane protein genes. Vet. Microbiol.124:230 –238.
14. Harley R, Day S, Di Rocco C, Helps C.2010. TheChlamydophila felis
plasmid is highly conserved, Vet. Microbiol.146:172–174.
15. Hartley JC, et al.2001. Conjunctivitis due toChlamydophila felisacquired from a cat: case report with molecular characterization of isolates from the patient and the cat. J. Infect.43:7–11.
16. Kari L, et al.2008. Pathogenic diversity amongChlamydia trachomatis
ocular strains in nonhuman primates is affected by subtle genomic varia-tions. J. Infect. Dis.197(3):449 – 456.
17. Klint M, et al.2010. Mosaic structure of intragenic repetitive elements in histone H1-like protein Hc2 varies within serovars ofChlamydia
tracho-matis. BMC Microbiol.10:81. doi:10.1186/1471-2180-10-81.
18. Kuo CC, Stephens RS, Bavoil PM, Kaltenboeck B. 2011. Genus I.
Chlamydia, p 846 – 865.InKrieg NR, et al. (ed), Bergey’s manual of
sys-tematic bacteriology, 2nd ed., vol. 4. Springer Verlag, New York, NY. 19. Laroucau K, et al.2008. High resolution ofChlamydophila psittaciby
multilocus VNTR analysis (MLVA). Infect. Genet. Evol.8(2):171–181. 20. Laroucau K, et al.2009. Genotyping ofChlamydophila abortusstrains by
multilocus VNTR analysis. Vet. Microbiol.137(3– 4):335–344. 21. Marrie TJ, Peeling RW, Reid T, De Carolis E, Canadian
Community-Acquired Pneumonia Investigators.2003.Chlamydiaspecies as a cause of community-acquired pneumonia in Canada. Eur. Respir. J.21(5):779 – 784.
22. May SW, Kelling CL, Sabara M, Sandbulte J.1996. Virulence of feline
Chlamydia psittaciin mice is not a function of the major outer membrane
protein (MOMP). J. Vet. Microbiol.53:355–368.
23. O’Dair HA, Hopper CD, Gruffydd-Jones TJ, Harbour DA, Waters L. 1994. Clinical aspects ofChlamydia psittaciinfection in cats infected with feline immunodeficiency virus. Vet. Rec.134:365–368.
24. Pannekoek Y, et al.2008. Multi locus sequence typing ofChlamydiales: clonal groupings within the obligate intracellular bacteriaChlamydia
tra-chomatis. BMC Microbiol.8:42. doi:10.1186/1471-2180-8-42.
25. Pannekoek Y, et al.2010. Multi locus sequence typing ofChlamydia
reveals an association betweenChlamydia psittacigenotypes and host spe-cies. PLoS One5:e14179. doi:10.1371/journal.pone.0014179.
26. Pantchev A, Sting R, Bauerfeind R, Tyczka J, Sachse K.2010. Detection of allChlamydophilaandChlamydiaspp. of veterinary interest using
on May 16, 2020 by guest
http://jcm.asm.org/
cies-specific real-time PCR assays. Comp. Immunol. Microbiol. Infect. Dis.33(6):473– 484.
27. Pedersen LN, Pødenphant L, Møller JK.2008. Highly discriminative genotyping ofChlamydia trachomatisusingomp1and a set of variable number tandem repeats. Clin. Microbiol. Infect.14(7):644 – 652. 28. Pudjiatmoko, Fukushi H, Ochiai Y, Yamaguchi T, Hirai K. 1996.
Seroepidemiology of feline chlamydiosis by microimmunofluorescence assay with multiple strains as antigens. Microbiol. Immunol.40:755–759. 29. Pudjiatmoko, Fukushi H, Ochiai Y, Yamaguchi T, Hirai K. 1997. Phylogenetic analysis of the genusChlamydiabased on 16S rRNA gene sequences. Int. J. Syst. Bacteriol.47(2):425– 431.
30. Pudjiatmoko, Fukushi H, Ochiai Y, Yamaguchi T, Hirai K. 1997. Diversity of felineChlamydia psittacirevealed by random amplification of polymorphic DNA. Vet. Microbiol.54:73– 83.
31. Regan RJ, Dathnan JRE, Treharne JD.1979. Infective endocarditis and glomerulonephritis associated with catChlamydia(C.psittaci) infection. Br. Heart J.42:349 –352.
32. Reyes JF, Chan CH, Tanaka M.2011. Impact of homoplasy on variable numbers of tandem repeats and spoligotypes inMycobacterium tubercu-losis. Infect. Genet. Evol. doi:10.1016./j.meegid.2011.05.018.
33. Sayada C, et al.1994. Homogeneity of the major outer membrane protein gene of felineChlamydia psittaci. Res. Vet. Sci.56:116 –118.
34. Sykes JE, Anderson GA, Studdert VP, Browning GF.1999. Prevalence of felineChlamydia psittaciand feline herpesvirus 1 in cats with upper respi-ratory tract disease. J. Vet. Intern. Med.13:153–162.
35. Sykes JE, Allen JL, Studdert VP, Browning GF.2001. Detection of feline calicivirus, feline herpesvirus 1 andChlamydia psittacimucosal swabs by multiplex RT-PCR/PCR. Vet. Microbiol.81:95–108.
36. Sykes JE.2005. Feline chlamydiosis. Clin. Tech. Small Anim. Pract.20(2): 129 –134.
37. TerWee J, et al.1998.Characterization of the systemic disease and ocular signs induced by experimental infection withChlamydia psittaciin cats. Vet. Microbiol.59:259 –281.
38. Vergnaud G, Pourcel C.2006. Multiple locus VNTR (variable number tandem repeat) analysis. Springer-Verlag, Berlin, Germany.
39. Wang Y, et al.2011. Evaluation of a high resolution genotyping method
forChlamydia trachomatisusing routine clinical samples. PLoS One
6:e16971. doi:10.1371/journal.pone.0016971.
40. Wills JM, Gruffydd-Jones TJ, Richmond SJ, Gaskell RM, Bourne FJ. 1987. Effect of vaccination on felineChlamydia psittaciinfection. Infect. Immun.55:2653–2657.
41. Wills JM, Howard PE, Gruffydd-Jones TJ, Wathes CM.1988. Prevalence ofChlamydia psittaciin different cat populations in Britain. J. Small Anim. Pract.29:327–339.