M E T H O D O L O G Y A R T I C L E
Open Access
Establishing quantitative real-time
quaking-induced conversion (qRT-QuIC) for highly
sensitive detection and quantification of PrP
Sc
in
prion-infected tissues
Song Shi, Gerda Mitteregger-Kretzschmar, Armin Giese and Hans A Kretzschmar
*Abstract
Background:PrPSc, the only known constituent of prions, the infectious agents causing prion diseases, can be detected by real-time quaking-induced conversion (RT-QuIC). However, there is no efficient method to quantify the amount of PrPScby RT-QuIC.
Results:Here we introduce quantitative RT-QuIC (qRT-QuIC) to quantify with high accuracy minute amounts of PrPScin the brain and various peripheral tissues at levels far below detection byin vivotransmission. PrPScis relatively resistant to treatment with proteinase K (PK). However, as there can also be a fraction of pathological PrP that is digested by PK, we use the term PrP27-30to denote to the amount of PrPScthat can be detected by immunoblot after PK treatment. qRT-QuIC is based upon the quantitative correlation between the seeded amount of PrP27-30and the lag time to the start of the conversion reaction detected by RT-QuIC. By seeding known amounts of PrP27-30quantified by immunoblot into qRT-QuIC a standard calibration curve can be obtained. Based on this calibration curve, seeded undetermined amounts of PrP27-30can be directly calculated. qRT-QuIC allowed to quantify PrP27-30concentrations at extremely low levels as low as 10-15.5g PrP27-30, which corresponds to 0.001 LD50 units obtained byin vivoi.c. transmission studies. We find that PrP27-30concentration increases steadily in the brain after inoculation and can be detected at various time points during the incubation period in peripheral organs (spleen, heart, muscle, liver, kidney) in two experimental scrapie strains (RML, ME7) in the mouse.
Conclusions:We suggest that an automatic quantitative system to measure disease progression as well as prion contamination of organs, blood and food product is feasible. Moreover, the concept of qRT-QuIC should be applicable to measure other disease-associated proteins rich inβ-pleated structures (amyloid) that bind ThT and that show seeded aggregation.
Keywords:Prion, PrPSc, PrP27-30, Quantitative RT-QuIC
Background
Prion diseases are a group of transmissible neurodege-nerative lethal disorders in humans, cattle, sheep, elk, mink and experimental animals. These diseases are cha-racterized by neuronal death and the accumulation of pathological disease-associated prion protein (PrPSc) in the central nervous system [1]. PrPSc is thought to be the essential, if not the exclusive, component of the
transmissible agent, or prion. Prion propagation seems to rely on autocatalytic amplification of PrPScby converting the host-encoded cellular prion protein (PrPC) to the pathogenic PrPScform without the participation of nucleic acids [2,3]. The conversion of PrPC to PrPSc is a post-translational event and involves a conformational change of the protein [4,5]. To distinguish between PrPScwhich is isolated from infectious tissue and isper definitionem as-sociated with the TSE agent on the one hand and structu-rally altered PrP, which has been converted into a ProteinaseK-resistant formin vitro, on the other, we refer * Correspondence:[email protected]
Center for Neuropathology and Prion research, Ludwig-Maximilians-University, Munich, Germany
to the latter as ‘PrPres’. The most commonly used approach to distinguish PrPScfrom PrPC when analyzing infected tissues is based on pre-treatment with proteinase K as PrPScis relatively PK-resistant. However, as there can also be a fraction of pathological PrP that is digested by PK treatment [6], we use the term PrP27-30 [2] in this manuscript whenever we refer to the amount of PK-resistant PrPScthat can be detected by immunoblotting.
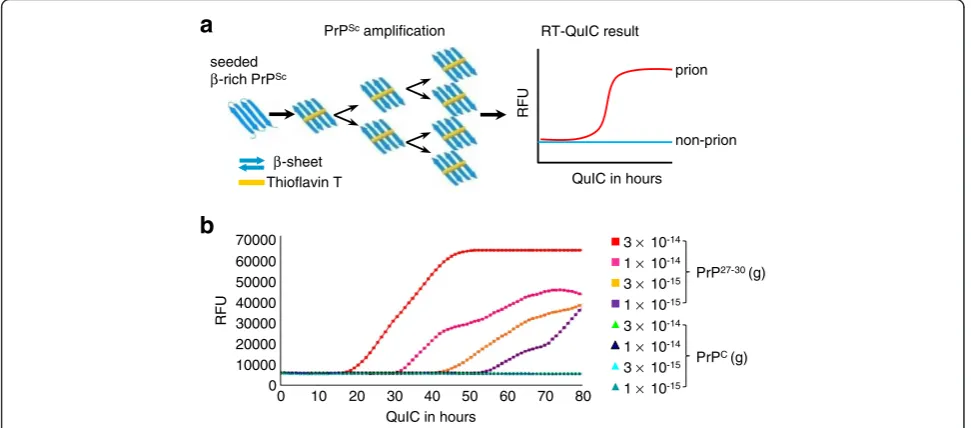
The infectivity of prions presents a serious risk to human health. One important issue in prion research is the sensitive detection and quantification of prions at low levels during the incubation period and in peripheral tissues to provide suitable detection assays for medicine and bio safety. Real-time quaking-induced conversion (RT-QuIC) was established to detect prion infectivity with very high sensitivity from diluted brain [7], from cerebrospinal fluid (CSF) of end-stage disease containing only very low levels of infectivity [8-10], and from CSF during the incubation period [11]. The mechanism of RT-QuIC is based upon the conversion of PK-sensitive recombinant prion protein (rPrPsen) into a PK-resistant rPrP (rPrPres) conformer, which is rich inβ-sheet struc-tures, by seeding the reaction mixture with PrPSc and periodic shaking (Figure 1a). The correlation between Thioflavin T (ThT) fluorescence and rPrPres has been shown previously and is used to monitor the conversion in a real-time curve [8].
Although protease-resistant PrP27-30is often used as a definitive biological marker for TSE infection, the titer
of infectivity measured by inoculation in experimental animals does not always fully correlate with the amount of PrP27-30 detected by immunoblot. The correlation of the seeding activity on rPrPsen conversion into rPrPres measured by RT-QuIC with the amount of PrP27-30 is also a complex issue. For example, it has been shown that vCJD prions have less seeding activity than sCJD prions despite the relatively high PrP27-30 concentration [10], and that prions from brains of 263 K-affected mice with little immunoblot-detectable PrP27-30have a seeding activity comparable to that associated with the high-PrPSc strain, 139A [12]. In previous studies, we showed that the complex correlation between amounts of PrP27-30 and infectivity in vivo and seeding activity in vitro can be explained at least in part by differences in size distribution of PrP aggregates [13]. Consequently, RT-QuIC seeding activity may correlate more closely with prion infectivity than with PrP27-30 levels, which can be considered an advantage in regard to the development of assays for prion detection.
We observed that in the RT-QuIC reaction, adding small amounts of PrPScresulted in a delayed initiation of conversion detected by ThT fluorescence (Figure 1b). This delay and the shape of the resulting ThT fluores-cence curve appeared similar to the kinetics of amplifi-cation of DNA that is used for quantitative RT-PCR analysis [14,15]. Thus we investigated whether real-time protein amplification was quantitative and established a highly sensitive assay for the quantification of prion
b
3 ×10-14
1 ×10-14
3 ×10-15
1 ×10-15
3 ×10-14
1 ×10-14
3 ×10-15
1 ×10-15
PrP27-30 (g)
PrPC (g)
70000 60000 50000 40000 30000 20000 10000 0
0 10 20 30 40 50 60 70 80 QuIC in hours
RF
U
a
PrPScamplificationThioflavin T
β-sheet seeded
β-rich PrPSc prion
non-prion
RF
U
QuIC in hours RT-QuIC result
Figure 1The basis of amplifying PrPScwith RT-QuIC. (a)Schematic illustration of RT-QuIC. PrPScconverts rPrPsen to rPrPres thereby
increasing the total amount ofβ-sheeted PrP. This increase can be demonstrated by the increased value of ThT-fluorescence. Therefore, the sample containing PrPSc(prion) is distinguished from that without PrPSc(non-prion). RFU, relative fluorescence units.(b)However, it was not the
amount of newly formed rPrPres but the time to reach the steep increase of amplification (ascendant curves) that was related to the seeded quantity of PK-treated PrPSc(PrP27-30). Different amounts of purified mouse RML scrapie-prion PrP27-30and normal mouse PrPCwere seeded into
infectivity in a high-throughput system based on mea-suring lag time of detectable conversion. We termed this approach qRT-QuIC and show that it allows quantita-tion of prions in various tissues with a detecquantita-tion limit corresponding to 0.001 LD50units.
Results and discussion
Establishing quantitative RT-QuIC
To establish a high-throughput quantification system, PrP27-30derived from two mouse-adapted scrapie prion strains, RML and ME7, was purified from infected C57BL/6 mouse brain by repeated NaCl precipitation allowing recovery of 97% of the total PrPSc [16]. The concentration of protease-resistant PrPScwas estimated by semi-quantitative immunoblotting and comparing band intensities to reference samples containing known quantities of rPrPsen [17] (data not shown). To estimate the minimum amount of PrP27-30 equivalent that can start the seeding reaction, we serially diluted PrP27-30 (from 10-8to 10-11.5g) (Figure 2a). 10-9.5g of both RML and ME7 PrP27-30were visible as a faint band on the im-munoblot while 10-10g was not detectable. As the con-trol, PrPC from healthy C57BL/6 mouse brain was purified [18] and quantified as above. For controlling the quality of the real-time curve, the ThT-binding fluorescence of both PrP27-30 and PrPC was measured (Figure 2b); the results showed that the fluorescence starting from 10-9.5g of PrP27-30was identical to that of
PrPC and the blank (no PrP). Since 10-10 g of PrP27-30 from both prion strains was undetectable in either immunoblot or ThT-fluorescence, we chose it as the initial seed.
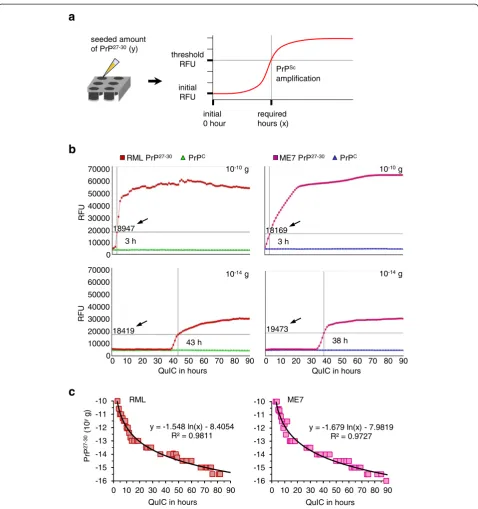
To establish the quantitative RT-QuIC (qRT-QuIC), the seeds containing 10-10 to 10-16 g of purified RML and ME7 PrP27-30 and PrPC per 10μl were prepared by serial half-log (100.5-fold) dilution (a total of 39 samples). Full-length mouse rPrPsen (amino acids 23–230) was utilized as the substrate. To estimate the time-span of the reaction, 130 h of RT-QuIC was tested for determin-ing the appropriate duration. Spontaneous conversion seemed to occur after 90 h (Additional file 1: Figure S1). For standardizing the results, a‘positive reaction’of RT-QuIC was recorded when the detected intensity of fluo-rescence was equal or higher than a threshold. The threshold was 3 times of the fluorescence of the initial phase (0 h). The corresponding time (hours) required to reach the threshold was recorded as the independent vari-able (x), and the equivalent amount of seeded PrP27-30was the dependent variable (y), as shown in Figure 3a. Next, RT-QuIC reactions seeded with 10-10to 10-16g of PrP27-30 and PrPCwere performed up to 90 h (shown in Figure 3b and Additional file 1: Figures S2 and S3). Five repeats of RT-QuIC for each seeded amount of PrP27-30 and PrPC equivalents (a total of 260 reactions) were tested. Reac-tions seeded with 10-10to 10-15.5 g of PrP27-30were posi-tive within 90 h whereas most of those seeded with 10-16g
PrPC
no PK
RML PrP27-30(g)
kDa
45 35
26
17 55
ME7 PrP27-30(g)
PrPC
no PK kDa
45 35
26
17 55
RF
U
quantity of protein (g) 10-8 10-9 10-9.5 10-10 10-10.5
b
a
0 10000 20000 30000 40000 50000
RML PrP27-30 ME7 PrP27-30 PrPC no PrP
Figure 2Directly detecting purified PrP27-30with immunoblotting and ThT-binding fluorescence. (a)Purified mouse RML and ME7
scrapie-prion PrP27-30was serially diluted and detected by immunoblotting. Aliquots were digested with 100μg/ml Proteinase K (PK) before
loading on the gel. Undigested PrPCis shown as a migration control. Signals were detected by 4H11 monoclonal antibody. M
ris shown on the
right.(b)Fluorescence of purified RML and ME7 PrP27-30and normal PrPCwas measured after ThT-binding. The mean and s.e.m. are
RML PrP27-30 PrPC
b
3 h
ME7 PrP27-30 PrPC
3 h
Pr
P
27-30
(10
yg)
c
RML
QuIC in hours
y = -1.548 ln(x) - 8.4054 R² = 0.9811
-16 -15 -14 -13 -12 -11 -10
0 10 20 30 40 50 60 70 80 90
ME7
QuIC in hours
y = -1.679 ln(x) - 7.9819 R² = 0.9727
-16 -15 -14 -13 -12 -11 -10
0 10 20 30 40 50 60 70 80 90
43 h 38 h
10-14g
18419
18947 18169
19473 70000
60000
50000 40000 30000
20000 10000 0
0 10 20 30 40 50 60 70 80 90 70000
60000
50000
40000 30000
20000 10000 0
10-10g
RF
U
RF
U
10-10g
10-14g
QuIC in hours
0 10 20 30 40 50 60 70 80 90 QuIC in hours
initial 0 hour
required hours (x)
initial RFU
threshold RFU
PrPSc
amplification
a
seeded amount of PrP27-30(y)
Figure 3Establishing the quantitative RT-QuIC (qRT-QuIC). (a)Schematic illustration of qRT-QuIC. The PrP27-30propagating duration (hour)
required to reach the threshold which was at least 3 times the starting fluorescence was set as the independent variable (x), the correlated seeded amount of PrP27-30was the dependent variable (y).(b)Different amounts of purified PrP27-30(10-10to 10-16g with serial 100.5-fold
dilution) were seeded into RT-QuIC reactions using mouse rPrPsen as the substrate. The RT-QuIC process was followed from 0 to 90 h by showing the number of hours required to reach the threshold (indicated by black arrows and intersecting lines). Purified PrPCwith identical
amounts was seeded in independent RT-QuIC reactions as control. Seeded amounts of both PrP27-30and PrPCare indicated on the top right; see
also Additional file 1: Figures S2 and S3.(c)The results from five repeats of RT-QuIC seeded by PrP27-30were provided to yield standard
calibration curves and formulas for quantification. This relates QuIC time necessary to reach the threshold to the amount of seeded PrP27-30. The
PrP27-30were negative (Additional file 1: Figure S4). Reac-tions seeded with different amounts of PrPCdid not show spontaneous conversion up to 90 h. Therefore, the RT-QuIC allowed detecting 10-15.5 g (≈ 0.32 fg) of PrP27-30, which was 1 million times more sensitive than the immu-noblot shown in Figure 1a. By analyzing the distribution of the required hours mathematically using a standard tool (Microsoft Excel), we obtained calibration curves and derived two formulas for calculating the quantities of seeded RML and ME7 PrP27-30in the qRT-QuIC system (Figure 3c).
Measuring PrP27-30concentrations in brains and
peripheral organs at different dpi of prion infection
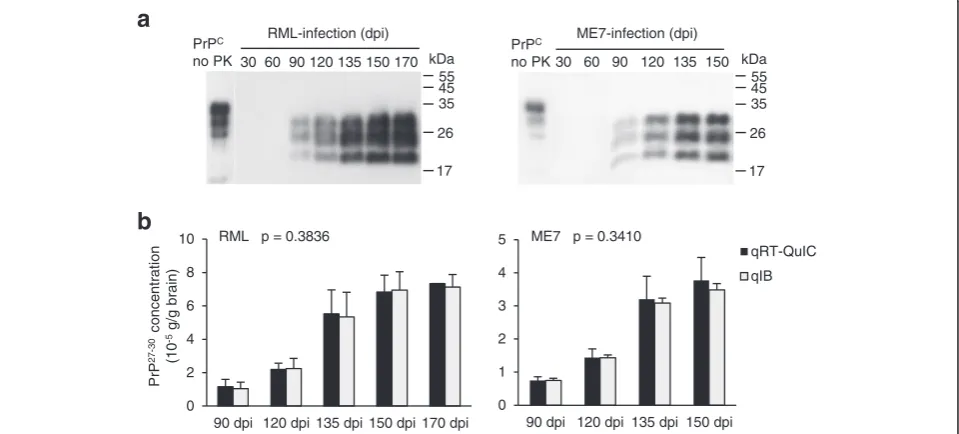
To see the feasibility of qRT-QuIC to determine the pro-gression of prion disease, qRT-QuIC was used to meas-ure PrP27-30 at different days post inoculation (dpi). 30, 60, 90, 120, 135, 150 and 170 dpi were chosen for RML, 30 to 150 dpi were chosen for ME7, since 170 dpi de-notes the terminal stage of the RML strain while 150 dpi is the terminal stage of the ME7 strain. Groups of each 5 C57BL/6 mice were inoculated intracerebrally and the brains and peripheral tissues (heart, liver, spleen, lung, kidney and hind-limb muscle) were harvested at the dpi indicated above. The tissues of age-related healthy C57BL/6 mice were chosen as controls (n = 5 for each dpi). 10 mg of each tissue were treated with the method of purifying PrP27-30 for obtaining tissue extracts as the seeds of RT-QuIC reactions. This purification step was
important to remove potential components affecting RT-QuIC efficiency. Since the PrP27-30 level is much higher in the brain than in other organs, 1 mg of brain from infected mice of different time-points was first analyzed by immunoblotting for obtaining an overview of the presence of PrP27-30 (Figure 4a). By immunoblotting, neither in RML nor in ME7 was PrP27-30 detectable at 30 and 60 dpi, in both strains PrP27-30 first appeared at 90 dpi and showed a steady increase to the terminal stage. For the preparation of RT-QuIC seeds, the brain extracts of 90 to 170 dpi of the brain were diluted 104 -fold, and those of the spleen and muscle were diluted 10-fold. Before measuring PrP27-30 concentrations in peripheral organs, a comparison was done to estimate the reliability of qRT-QuIC (Figure 4b). The results showed that PrP27-30concentrations in the brains obtained from qRT-QuIC were comparable to those from quantita-tive immunoblottings at different time points after inocu-lation (see also in Additional file 1: Table S1), indicating that qRT-QuIC is suited to measure the PrP27-30 concentrations.
After 90 h of amplification we calculated the concentra-tion of PrP27-30equivalents based on the detected seeding activity by using the formulas in Figure 3c. The levels of PrP27-30 equivalents (i.e. seeding activity) in the brain showed the expected increasing tendency (Figure 5). In particular, seeding activity reached the highest levels at 170 dpi in the RML strain and 150 dpi in the ME7 strain. The detection of seeding activity from 30 to 90 dpi of both
Pr
P
27
-30
co
n
c
e
n
tr
a
tio
n
(10
-5
g/
g brain)
RML p = 0.3836
0 2 4 6 8 10
90 dpi 120 dpi 135 dpi 150 dpi 170 dpi 0 1 2 3 4 5
90 dpi 120 dpi 135 dpi 150 dpi
qRT-QuIC qIB ME7 p = 0.3410
PrPC
no PK 30 60 90 120 135 150 170 RML-infection (dpi)
a
45 35
26
17 kDa 55
PrPC
no PK 30 60 90 120 135 150 ME7-infection (dpi)
45 35
26
17 kDa 55
b
Figure 4Quantification of RML and ME7 PrP27-30in brain at various days post infection (dpi). (a)1 mg of brain from scrapie-infected mice
at was analyzed by immunoblotting after digestion with 100μg/ml PK. PrPCis shown as a migration control. Signals were detected by 4H11
monoclonal antibody. Mris shown on the right.(b)Comparing PrP27-30concentrations measured by quantitative RT-QuIC and quantitative
immunoblot. The brains of RML-inoculated mice 90 dpi to 170 dpi and ME7-inoculated mice 90 to 150 dpi were chosen regarding the capacity of immunobloting to detect PrP27-30from the time-point not earlier than 90 dpi. The significance (p value) was calculated with two-way ANOVA
prion strains was negative in the heart and hind-limb, but the signals from the heart started to show at 135 dpi in the RML strain and 120 dpi in the ME7 strain, while those from muscle were positive starting at 120 dpi of the RML strain and 90 dpi of ME7. Interestingly, seeding activity in the liver and kidney in the RML strain was detectable in early stages (30 and 60 dpi), and disappeared during the intermediate stages of infection (90, 120 and 135 dpi), whereas those of ME7-infection were continuously negative till 135 dpi. Seeding activity in the spleen of both prion strains was decreasing at early stages followed by an increase in the intermediate stages. The concentration of PrP27-30equivalents in all three tissues was increased at late to final stages. No seeding activity was detected in the lungs at any time. Since qRT-QuIC was 1 million times more sensitive than normal immunoblotting, these results suggest a possibility of using qRT-QuIC to track disease progres-sion or analyze prion propagation in various tissues.
To demonstrate the feasibility of qRT-QuIC in the assessment of bio-hazard risks, the concentration of PrP27-30/g tissue (g/g) in various tissues measured by qRT-QuIC was compared to the reported half-lethal doses (LD50) of both prion strains. The known LD50 of
RML and ME7 are 10-8.8and 10-8.3g of terminal brains, respectively [19]. Using the PrP27-30 concentration measured by qRT-QuIC, we can roughly extrapolate that 1 LD50 unit contained 10-12.93 g of PrP27-30 for
RML and 10-12.72g for ME7. One LD50unit is shown as
the gray dotted lines in Figure 5 to indicate the extent of prion concentration in each gram of tissue (see also Additional file 1: Tables S2 and S3), suggesting that qRT-QuIC can be used for estimating prion contami-nation in biological materials. As our protocol uses an extraction method for PrP27-30 prior to qRT-QuIC that removes potentially interfering factors present in peri-pheral tissues, the calibration curve obtained for brain-derived PrP27-30 should also provide meaningful results for other tissues from the same species. Moreover, our
findings for RML and ME7 indicate that similar assay conditions can be efficient for different strains. How-ever, to ensure optimum sensitivity, the exact conditions of the qRT-QuIC assay need to be established for differ-ent species and strains.
In a recent study, Wilham and colleagues [7] used SD50 (50% seeding dose) to quantify seeding activity in the RT-QuIC. SD50 was defined as minimum seeded amount causing 50% of RT-QuIC reactions to be posi-tive. Using end-point titration in a dilution series for quantification, SD50 was shown to correlate with the infectivity of 263 K strains in hamster. To obtain SD50 values for one prion strain in RT-QuIC, serially diluted standard sample are required, e.g., serially diluted prion-infected reference brain homogenate and serially diluted undetermined materials. Thus, this approach requires multiple repeats to yield the percentage of positive reac-tions for every dilution. In contrast, qRT-QuIC is a lag time-based assay and the amount of PrP27-30equivalents in suspected materials can be directly calculated and quantified with much fewer repeats based on the calibra-tion curve. The respective advantages and disadvantages of end-point titration and lag time assays are well known from assays of prion infection in vivo. End-point titra-tion provides an accurate measure of infectious units. However, end-point in vivo assay also requires multiple repeats for each dilution of seeds to obtain the titration of one prion strain, whereas the incubation timein vivo assay is more commonly used for prion quantification as much less animals are required. Hence, we conclude that the qRT-QuIC assay which uses lag time is more suitable than SD50 measurements for quantitative detection and high-throughput assay for prion diagnosis.
Conclusion
In conclusion, qRT-QuIC is a new advancement of the RT-QuIC system, which enables us to quantify PrP27-30 concentrations at extremely low levels as low as 10-15.5g PrP27-30, which corresponds to 0.001 LD50 units. qRT--15
-13 -11 -9 -7 -5 -3
30 dpi
60 dpi
90 dpi
120 dpi
135 dpi
150 dpi
170 dpi
Pr
P
27
-3
0e
q
u
iv
a
len
t
(10
yg/
g t
is
s
ue)
-15 -13 -11 -9 -7 -5 -3
30 dpi
60 dpi
90 dpi
120 dpi
135 dpi
150 dpi
brain heart
liver spleen kidney
muscle
RML-infection ME7-infection
1 LD50unit/g
Figure 5The seeding activity (expressed as PrP27-30equvalents in g/g tissue) in various tissues from different time-points after
inioculation were measured by qRT-QuIC by using the formulas shown in Figure 3c.The PrP27-30concentration corresponding to 1 LD 50
QuIC is based upon the quantitative correlation between the seeded amount of PrP27-30 and the lag time to the start of the conversion reaction detected by RT-QuIC. The principle of generating standard curves by monitoring the fluorescence and setting threshold levels makes qRT-QuIC applicable to the development of an automatic system similar to quantitative RT-PCR. qRT-QuIC can be used as a rapid and reliable novel research tool, increase the precision of diagnosis, and help to follow the effects of therapy of prion diseases. Moreover, this concept should be applicable to measure other proteins rich inβ-pleated structures (amyloids) that bind ThT and that show seeded aggregation.
Methods
rPrPsen expression and purification
BL21 (DE3) E.coli and pET41a system (Merk, Germany) were used to express mouse rPrPsen. Bacteria were cul-tured in LB medium at 37°C with 220 rpm shaking and were added 1 mM of IPTG (final concentration) when OD reached 0.7 to 0.9. The cell pellet was harvested by 10,000 g centrifugation at 4°C for 10 min after 4 hours in-duction. Thereafter, cells were resuspended by BugBuster Master Mix (Novagen, Germany) containing rLysozyme and Benzonase for extracting inclusion bodies (manual for BugBuster, Novagen, Germany). The extracted inclusion bodies were denatured with 8 M Guanidine hydrochloride at 25°C for 1 hour. The denatured protein solution was centrifuged at 16,000 g at 4°C for 20 min to remove the debris.
We chose Ni2+-NTA superflow (Qiagen, Germany) resin for protein purification. The NTA resin was pre-pared by following the manufacturer’s manual. The de-natured protein was loaded onto prepared resin followed by binding with inversion on the rotor at 25°C for 1 hour. After that, the resin was loaded into the column (GE healthcare, USA) followed by connecting to the AKTA prime (GE healthcare, USA). The followed purification procedures including refolding and elution were performed as described [7].
After elution, the rPrPsen solution was loaded into 6 KDa Cellu Sep dialysis tubing (Interchim, France) followed by immersion in pre-chilled dialysis buffer (9 mM NaH2PO4, 1 mM Na2HPO4, pH 5.9) at 4°C.
The dialysis was done in 2 successive steps, which were 2 hours and 18 hours, respectively. 100 volumes of dialysis buffer were used for each step. The dia-lyzed solutions were sterilized with a 0.22 μm filter (Millipore, USA) and the absorption was measured at 280 nm for calculating the concentration of rPrPsen. The concentration of each fraction was adjusted to 0.5 mg/ml by adding sterilized and chilled dialysis buffer. The rPrPsen solution was aliquoted and frozen
in liquid nitrogen, followed by transfer to a freezer (−80°C) for long-term storage.
Tissue preparation
6-week old C57BL/6 mice were inoculated with mouse-adapted RML and ME7 scrapie (i.c). 10 μl of 10% brain homogenate in 1 × PBS (pH 7.2) was used for each inoculum. 7 time-points (30, 60, 90, 120, 135, 150 and 170 dpi, days post inoculation) were set for RML-infection and 6 time-points (30, 60, 90, 120, 135 and 150 dpi) were for ME7-infection. At each dpi, five mice were sacrificed with CO2. To harvest the tissues (brain, heart,
liver, spleen, lung, kidney and hindlimb muscle) they were washed with chilled 1 × PBS containing 5% sodium citrate to remove the blood. Then tissues were weighed and stored in liquid nitrogen. The tissues of age-related control C57BL/6 mice were prepared following the same procedure.
PrP27-30purification
We followed a published protocol [16] to purify PrP27-30. The RML- and ME7-infected mouse tissue was prepared to 10% homogenate (w/v) with lysis buffer (pH 7.2) containing 130 mM NaCl, 10 mM NaH2PO4, 10 mM
Na2HPO4, 0.5% Triton X-100, 0.5% sodium deoxycholate,
2 mM MgCl2, 2.5 U/ml of Benzonase (Merck, Germany)
and EDTA-free protease inhibitor cocktail (Roche, Switzerland). Homogenate was incubated at 25°C for 30 min for digesting nucleic acids followed by 1,000 g centrifugation at 4°C for 5 min for removing debris. Thereafter, 100μl of supernatant were doubly diluted with lysis buffer to reach 200 μl of total volume followed by 20μg/ml of PK-digestion at 37°C for 1 h. The digestion was stopped by adding 5 mM PMSF (Sigma-Aldrich, Switzerland) and then the PK-treated supernatants were transferred into 300μl of 1 × QuIC buffer (130 mM NaCl, 5 mM NaH2PO4, 5 mM Na2HPO4 and 1 mM EDTA,
pH 7.4). The total volume reached 500μl.
The preparations were then brought to equal volume of buffer containing 20% NaCl and 0.1% sarkosyl. These solutions were vortexed vigorously followed by incuba-ting on ice with gentle shaking for 10 min. After centri-fugation at 16,000 g at 4°C for 10 min, the pellets were washed by 500μl of 20 mM Tris–HCl containing 0.05% sarkosyl followed by 16,000 g of centrifugation at 4°C for 10 min. This washing step was repeated twice. The pellet was stored at−80°C till being used as the seed.
For preparing the prion seeds, the frozen pellets were thawed at 4°C followed by washing with 500 μl of ddH2O. The resuspended solutions were precipitated by
centrifugation at16,000 g for 10 min at 4°C. This step was repeated twice. The last pellets were resxuspended thoroughly by 50 μl of ddH2O followed by 1,000 g of
from peripheral tissues (tissue extract) was brought to qRT-QuIC to be both the seed and required water. 10μl of PrP27-30 with known concentration quantified by semi-quantitative immunoblotting [17] was used as the seed for obtaining standard a calibration curve.
PrPCpurification
We referred to a published protocol [18] to purify mouse PrPC. The brain from a healthy 20-week old C57BL/6 mouse was homogenized in chilled 1 × PBS (pH 7.2) containing EDTA-free protease inhibitor cocktail (Roche, Switzerland) to make a 10% homogenate (w/v). After cen-trifugation at 3,000 g at 4°C for 30 min, the pellet was resuspended with an equal volume of chilled buffer (pH 7.2) containing 130 mM NaCl, 10 mM NaH2PO4,
10 mM Na2HPO4, 2% NP-40, 1% sodium deoxycholate
and EDTA-free protease inhibitor cocktail (Roche, Switzerland). After incubation on ice for 30 min, the ho-mogenate was subjected to 100,000 g centrifugation at 4°C for 30 min to remove the debris. The supernatant was filtrated with a 0.22μm filter (Millipore, USA) followed by pouring over the ImmunoPure Immobilized Protein A column (Pierce, USA) to remove the endogenous immu-noglobulins. The filtrated solution was incubated with 1 ml of Protein A resin cross-link to mouse-PrP specific monoclonal 4H11 antibody at 4°C over night. Then the resin was washed with 10 volumes of washing buffer (20 mM Tris–HCl, 500 mM NaCl and 5 mM EDTA, pH 8.0) followed by washing with 10 volumes of 1 × PBS containing 0.5% NP-40. PrPC was eluted with 5 ml of 200 mM glycine. The solution was loaded into 10 kDa fil-ter centrifugal tubes (Millipore, USA) to centrifuge at 3,000 g at 4°C for 1 h for desalting and buffer exchanging. Then the protein was resuspended with 5 ml of 1 × QuIC buffer and subjected to gel filtration by passing over the Superdex 75 column (GE lifescience, USA). The harvested protein in the peak was concentrated with a 10 kDa filter centrifugal tube (Millipore, USA) followed by estimating protein concentration with quantitative immunoblot. The yield of mouse PrPCwas approx. 3μg/brain.
Quantitative RT-QuIC
Preparations for quantitative RT-QuIC were loaded into a Nunc 96-well plate (Thermo Scientific, USA) in a bio-safety cabinet. The plate was sealed with transparent tape for avoiding cross-contamination and aerosol. Reac-tions were performed on a FLUOstar Optima (BMG Labtech, Germany) at 37°C for 90 hours with 1 min shaking at 600 rpm followed by 1 min stationary incuba-tion. The fluorescence was measured with bottom optic every hour. Excitation was 440 nm, emission was 480 nm and the gain setting was 2000. Curves and intersecting lines (for indicating both the threshold and required hour) were directly shown by the provided
functions of BMG Optima Data Analysis software. Other details are shown in the following Method Table 1.
PK-digestion and immunoblot
The 10% brain homogenates of RML- and ME7-infected mice and purified RML- and ME7-PrP27-30were digested with 100 μg/ml of proteinase-K (Roche, Switzerland) at 37°C for 1 h. The digestions were stopped by heating at 100°C for 10 min with 2 × loading buffer. The proteins were separated in a 15% SDS-PAGE followed by transfer to a PVDF membrane (Millipore, USA). The membrane was blocked with 5% non-fat milk for 1 h at room temperature. The proteins were detected with mouse-PrP specific 4H11 monoclonal antibody (a gift from E. Kremmer, National Research Center for Environment and Health, Munich, Germany). Signals were measured on a Diana III luminescence imaging system (Raytest, Germany).
Additional file
Additional file 1: Figure S1.Determining the time-span of qRT-QuIC. RT-QuIC reactions were seeded with PrP27-30or PrPCwith the indicated amounts. A non-seeded reaction was performed as the control. After 130 h at 37°C, both PrPC-seeded and non-seeded reactions showed rising
curves, indicating that the spontaneous conversion started at approx. 100 h. Therefore, we chose 90 hours as the maximum time-span of monitoring prion conversion in the qRT-QuIC system.Figures S2 and S3: Detecting seeded PrP27-30with RT-QuIC. Purified RML and ME7 PrP27-30
and control PrPCwith the quantities from 10-10to 10-16g were seeded
into reactions independently to perform 90 h of RT-QuIC at 37°C.Figure S4: The positive RT-QuIC reactions seeded with PrP27-30(10-10to 10-15.5g
for RML and 10-10to 10-16g for ME7) within 90 h are shown. Each scale
on the Y-axis represents one effective reaction, the X-axis indicates the required hours corresponding to the reaction. The reactions seeded with 10-16g of RML PrP27-30were negative up to 90 h and thus are not shown
in the figure.Table S1: Comparing PrP27-30concentrations measured by
quantitative RT-QuIC and quantitative immunoblot.Table S2: The concentration of PrP27-30in 7 tissues from RML scrapie-infected mice of 7 time-points.Table S3: The concentration of PrP27-30in 7 tissues from
ME7 scrapie-infected mice of 6 time-points.
Competing interests
The authors declare that they have no competing interests. Table 1 The ingredients of qRT-QuIC preparation
Component volume
10 x QuIC buffer
10μl (100 mM NaH2PO4, 100 mM Na2HPO4,
10 mM EDTA, 1.3 M NaCl, pH 6.9)
dd H2O 35μl for standard curve, not
for testing tissue extract
3 M NaCl 10μl
rPrPsen (0.3 mg/ml) 34μl (final 0.1 mg/ml)
1 mM thioflavin T 1μl
seed 10μl for standard curve, 45μl
for testing tissue extract
Authors’contributions
HK conceived the research. SS and GM performed the experiments and analyzed data. HK, SS and AG wrote the manuscript. All authors read and approved the final manuscript.
Acknowledgment
We thank Dr. B. Caughey, Dr. A.G. Hughson, G.J. Raymond (These three researchers are from NIAID, NIH, USA), Dr. L.I. McGuire and Dr. A.J. Green (both two researchers are from NCJDRSU, University of Edinburgh, UK) for nicely sharing their experiences on RT-QuIC technique. We thank Dr. E. Kremmer (National Research Center for Environment and Health, Munich, Germany) for providing 4H11 monoclonal antibody. We thank Janina Mielke for technique supports. This study was supported by the Deutsche Forschungsgemeinschaft (SFB 596).
Received: 20 July 2013 Accepted: 20 July 2013 Published: 2 August 2013
References
1. Prusiner SB:Novel proteinaceous infectious particles cause scrapie. Science1982,216:136–144.
2. Prusiner SB:Prions.Proc Natl Acad Sci USA1998,95:13363–13383. 3. Bieschke J, Weber P, Sarafoff N, Beekes M, Giese A, Kretzschmar H:
Autocatalytic self-propagation of misfolded prion protein.Proc Natl Acad Sci USA2004,101:12207–12211.
4. Caughey B, Raymond GJ, Ernst D, Race RE:N-terminal truncation of the scrapie-associated form of PrP by lysosomal protease(s): implications regarding the site of conversion of PrP to the protease-resistant state. J Virol1997,65:6597–6603.
5. Pan KM, Baldwin M, Nguyen J, Gasset M, Serban A, Groth D, Mehlhorn I, Huang Z, Fletterick RJ, Cohen FE:Conversion of alpha-helices into beta-sheets features in the formation of the scrapie prion proteins. Proc Natl Acad Sci USA1993,90:10962–10966.
6. Safar J, Wille H, Itri V, Groth D, Serban H, Torchia M, Cohen FE, Prusiner SB: Eight prion strains have PrP(Sc) molecules with different conformations. Nat Med1998,4:1157–1165.
7. Wilham JM, Orrú CD, Bessen RA, Atarashi R, Sano K, Race B, Meade-White KD, Taubner LM, Timmes A, Caughey B:Rapid end-point quantitation of prion seeding activity with sensitivity comparable to bioassays. PLoS Pathog2010,6:e1001217.
8. Atarashi R, Satoh K, Sano K, Fuse T, Yamaguchi N, Ishibashi D, Matsubara T, Nakagaki T, Yamanaka H, Shirabe S, Yamada M, Mizusawa H, Kitamoto T, Klug G, McGlade A, Collins SJ, Nishida N:Ultrasensitive human prion detection in cerebrospinal fluid by real-time quaking-induced conversion.Nat Med2011,17:175–178.
9. McGuire LI, Peden AH, Orrú CD, Wilham JM, Appleford NE, Mallinson G, Andrews M, Head MW, Caughey B, Will RG, Knight RS, Green AJ:Real time quaking-induced conversion analysis of cerebrospinal fluid in sporadic Creutzfeldt-Jakob disease.Ann Neurol2012,72:278–285.
10. Peden AH, McGuire LI, Appleford NE, Mallinson G, Wilham JM, Orrú CD, Caughey B, Ironside JW, Knight RS, Will RG, Green AJ, Head MW:Sensitive and specific detection of sporadic Creutzfeldt-Jakob disease brain prion protein using real-time quaking-induced conversion.J Gen Virol2012, 93:438–449.
11. Orrù CD, Hughson AG, Race B, Raymond GJ, Caughey B:Time course of prion seeding activity in cerebrospinal fluid of scrapie-infected hamsters after intratongue and intracerebral inoculations.J Clin Microbiol2012, 50:1464–1466.
12. Vascellari S, Orrù CD, Hughson AG, King D, Barron R, Wilham JM, Baron GS, Race B, Pani A, Caughey B:Prion seeding activities of mouse scrapie strains with divergent PrPSc protease sensitivities and amyloid plaque content using RT-QuIC and eQuIC.PLoS One2012,7:e48969.
13. Weber P, Reznicek L, Mitteregger G, Kretzschmar H, Giese A:Differential effects of prion particle size on infectivityin vivoand in vitro. Biochem Biophys Res Commun2008,369:924–928.
14. Schmittgen TD, Zakrajsek BA, Mills AG, Gorn V, Singer MJ, Reed MW: Quantitative reverse transcription-polymerase chain reaction to study mRNA decay: comparison of endpoint and real-time methods. Anal Biochem2000,285:194–204.
15. RadonićA, Thulke S, Mackay IM, Landt O, Siegert W, Nitsche A:Guideline to reference gene selection for quantitative real-time PCR.Biochem Biophys Res Commun2004,313:856–862.
16. Polymenidou M, Verghese-Nikolakaki S, Groschup M, Chaplin MJ, Stack MJ, Plaitakis A, Sklaviadis T:A short purification process for quantitative isolation of PrPScfrom naturally occurring and experimental transmissible spongiform encephalopathies.BMC Infect Dis2002,2:23. 17. Saá P, Castilla J, Soto C:Ultra-efficient replication of infectious prions by
automated protein misfolding cyclic amplification.J Biol Chem2006, 281:35245–35252.
18. Deleault NR, Harris BT, Rees JR, Supattapone S:Formation of native prions from minimal components in vitro.Proc Natl Acad Sci USA2007, 104:9741–9746.
19. Browning S, Baker CA, Smith E, Mahal SP, Herva ME, Demczyk CA, Li J, Weissmann C:Abrogation of complex glycosylation by swainsonine results in strain- and cell-specific inhibition of prion replication. J Biol Chem2011,286:40962–40973.
doi:10.1186/2051-5960-1-44
Cite this article as:Shiet al.:Establishing quantitative real-time quaking-induced conversion (qRT-QuIC) for highly sensitive detection and quantification of PrPScin prion-infected tissues.Acta
Neuropathologica Communications20131:44.
Submit your next manuscript to BioMed Central and take full advantage of:
• Convenient online submission
• Thorough peer review
• No space constraints or color figure charges
• Immediate publication on acceptance
• Inclusion in PubMed, CAS, Scopus and Google Scholar
• Research which is freely available for redistribution