Copyright 0 1990 by the Genetics Society o f America
Characterization and Nucleotide Sequence
of
the Cryptic Operon
of
Escherichia coli
K12
Laura L. Parker' and Barry G . Hall*
Department of Molecular and Cell Biology, University of Connecticut, Storrs, Connecticut 06269
Manuscript received April 6, 1989 Accepted for publication November 27, 1989
ABSTRACT
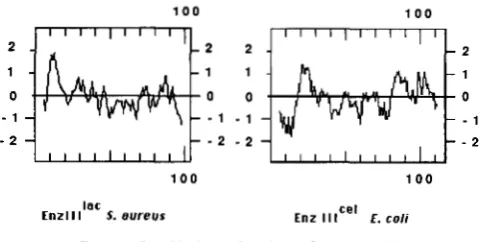
Wild-type Escherichia coli are not able to utilize @-glucoside sugars because the genes for utilization of these sugars are cryptic. Spontaneous mutations in the cel operon allow its expression and enable the organism to ferment cellobiose, arbutin and salicin. In this report we describe the structure and nucleotide sequence of the cel operon. The cel operon consists of five genes: celA, whose function is unknown; celB and celC which encode phosphoenolpyruvate-dependent phosphotransferase system enzyme 11"' and enzyme I W ' , respectively, for the transport and phosphorylation of @-glucoside sugars; celD, which encodes a negative regulatory protein; and celF, which encodes a phospho-@- glucosidase that acts on phosphorylated cellobiose, arbutin and salicin. The mutationally activated cel operon is induced in the presence of its substrates, and is repressed in their absence. A comparison of proteins encoded by the cel operon with functionally equivalent proteins of the bgl operon, another cryptic E. coli gene system responsible for the catabolism of @-glucoside sugars, revealed no significant homology between these two systems despite common functional characteristics. The celD and celF encoded repressor and phospho-@-glucosidase proteins are homologous to the melibiose regulatory protein and to the melA encoded a-galactosidase of E. coli, respectively. Furthermore, the celC encoded PEP-dependent phosphotransferase system enzyme 111"' is strikingly homologous to an enzyme 111'"'
of the Gram-positive organism Staphylococcus aureus. We conclude that the genes for these two enzyme
111s diverged much more recently than did their hosts, indicating that E. coli and S. aureus have
undergone relatively recent exchange of chromosomal genes.
W
ILD-type Escherichia coli cannot utilize P-glu- coside sugars because the gene systems respon- sible for their catabolism are not expressed. Sponta- neous mutants capable of utilizing these sugars are easily isolated and analysis of the mutants reveals the presence of several different systems. T h e bgl operon was the first characterized. It is located at 8 3 minutes on the E. coli linkage map, and specifies a phosphoen- olpyruvate-dependent transport protein encoded bybglF, a phospho-P-glucosidase encoded by bglB, and a
positive activator of the operon encoded by bglG
(SCHNETZ, TOLOCZYKI, and RAK 1987; MAHADEVAN,
REYNOLDS and WRIGHT 1987). T h e bgl operon can
be activated by the insertion of IS1 or IS5 upstream of the transcriptional start site, or by mutations in the cyclic-AMP receptor protein binding site (REYNOLDS,
FELTON and WRIGHT 1981 ; REYNOLDS et al. 1985,
1986). In the active state the operon is inducible by arbutin and salicin (SCHAEFLER and MASS 1967; PRA-
SAD and SCHAEFLER 1974).
bglA, a phospho-P-glucosidase specific for arbutin,
School, Boston, Massachusetts 021 ' 1 1 .
Present address: Department o f Physiology, Tufts University Medical
Rochester, Rochester, New York 14627.
Present address: Department of Biology, Hutchison Hall, University of
of pdge charges. This article must therefore be hereby marked "advertisement"
T h e publication costs of this article were partly defrayed by the payment
in accordance with 18 U.S.C. 51734 solely to indicate this fact.
Genetics 124: 455-471 (March, 1990)
lies outside of the bgl operon (PRASAD, YOUNG and
SCHAEFLER 1973). bglA is expressed constitutively in
wild type E. coli, but does not, by itself, permit growth on arbutin because the genes for transport of arbutin are cryptic.
arbT, encodes a phosphotransferase protein, spe-
cific for arbutin. This gene is cryptic in the wild type organism, but it can be activated by spontaneous mutations. In the active state, arbT is inducible by arbutin (KRICKER and HALL 1987).
Sac is a fourth cryptic gene system that can be activated by spontaneous mutations. T h e chromo- somal location of sac is not known, but it is not linked to either the bgl or cel operons. In the active state sac
is inducible and allows utilization of arbutin, salicin and cellobiose (PARKER and HALL 1988).
T h e cel operon is located at 37.8 min on the E . coli
linkage map, almost directly opposite the bgl operon
(KRICKER and HALL 1984). In addition to arbutin and
salicin, the cel operon allows utilization of cellobiose, a disaccharide hydrolysis product of cellulose. Bio- chemical assays have shown that the cel operon speci- fies a phosphoenolpyruvate dependent transport sys- tem and a phospho-P-glucosidase, both of which are active toward all three sugars (KRICKER and HALL
456 L. L. Parker and B. G . Hall
T h e transport and &glucosidase functions have been localized on the cel restriction map by transposon mutagenesis of a cloned Cel+ allele (HALL, BETTS and
KRICKER 1986). Based on the phenotypes of the inser- tionally inactivated alleles, it was determined that the transport gene is transcribed before the B-glucosidase gene and that the genes constituted an operon (HALL, BETTS and KRICKER 1986). T h e cel operon was mapped by PI transduction to 37.8 min on the E . coli
genetic map (KRICKER and HALL 1984). In good agreement with the earlier mapping data, we have located the cel operon on the physical map of E . coli
(KOHARA, AKIYAMA, and ISONO 1987) at 38 min by
comparing the KOHARA et al. restriction map with the restriction map of cel. The order of the restriction enzyme sites on this map, and our knowledge of the direction of transcription of the cel operon, indicates that the cel operon is transcribed counter-clockwise on the E . coli genetic map.
For purposes of discussion the genes for transport and hydrolysis of cellobiose had previously been des- ignated celT and celH, respectively (KRICKER and
HALL 1987). In this paper we change those designa- tions to more conventional notation. Because the Cel+ mutation in strain MK2 was cis-dominant the un- known site of the activating mutation was designated
celR. In the accompanying paper we show that celR is
a region just upstream of the coding region where insertion sequences can insert and thereby activate the operon (PARKER and HALL 1990).
In this report we describe the structure and nucleo- tide sequence of the cel operon. In addition to the permease and /3-glucosidase functions, the cel operon encodes a negative regulatory protein that functions at the level of transcription. T h e entire cel operon consists of five open reading frames, designated celA,
celB, celC, celD and celF. T h e function of celA remains
unknown, but in this paper we assign functional roles to each of the other four genes.
MATERIALS AND METHODS
Culture media and conditions: Minimal media consisted of a phosphate buffered mineral salts solution (HALL and BETTS 1987) with 1.5% agar as a solidifying agent. Sugars were used at a concentration of 0.1% (w/v) except for dextrose which was used at 0.2% (w/v). When required, amino acids were added at a concentration of 100 pg/ml, and ampicillin was used at 200 pg/ml. MacConkey indicator plates contained 1 % (w/v) of the appropriate sugar and were prepared according to the manufacturer (Difco). If the added sugar was utilized, colonies were pink or red, if not, colonies were white.
Strains and plasmids: E. coli strains and plasmids are listed in Table 1.
Plasmid constructions: Several plasmids were con- structed for the functional analysis of the cel operon. Plasmid pUF671 contains an EcoRI/BamHI fragment carrying the
cel operon from the Gel+ strain BGH2008 cloned into the EcoRI/BamHI sites of plasmid pBlu+ (PARKER and HALL
1990). pUF680 was created by deleting pUF671 from the 5' end of the cel operon to base pair 3260 by digestion with Exonuclease I and S1 nuclease. pUF681 was created by cloning the HindIII/HpaZ fragment containing celR and celA
from pUF671, into the HindIII/EcoRV sites of plasmid pBlu'. Plasmid pUF682 was created by deleting pUF671 from the 3' end of cel operon beginning at the BamHI site in the vector and ending at base pair 2240, leaving celR celA
and celB intact. pUF683 was created in the same manner and carries bp 1-3294, i.e., celR, celA, celB and celC. Plasmid pUF684 was created by digesting pUF683 with Sty1 and religating, resulting in the inframe deletion of bp 1591- 1925, entirely within celB. pUF685 was created by digesting pUF67 1 with AvaI and subsequently religating, resulting in the deletion of 700 bp of celD. pUF686 was created by digesting pUF673 with AvaI and subsequently religating, resulting in the deletion of bp 2488-3202, entirely within
celD. Plasmid pUF688 carries bp 2361-3362 of the wild type operon, includes all of celD gene, and was created by cloning the 1324 bp ClaIIEcoRV fragment of pUF673 into the ClaI/EcoRV sites of the plasmid pBlu+, and deleting the resulting plasmid using the enzymes Exonuclease 111 and S1 nuclease from the ClaI site to almost the end of celC at base pair 2361.
Molecular techniques: Methods for the preparation of plasmid DNA, isolation and labeling of probe DNA, DNA/ DNA hybridizations and DNA/RNA hybridizations were previously described (HALL and BETTS 1987). The probe specific for the entire cel operon was prepared from the 4.0- kilobase (kb) HpaI fragment of plasmid pUF564 (HALL, BETTS and KRICKER 1986). T h e celA specific probe was prepared from the 276 bp TaqI fragment of plasmid pUF681 (this study). T h e celB specific probe was prepared from the 1 164 bp EcoRVIHindIII fragment of plasmid pUF682 (this study). The celC specific probe (bp 2049- 2287) was prepared from the 239-bp Rsal fragment of plasmid pUF683 (this study). Preparation of competent cells and transformations with plasmid DNA were performed according to the method of MANIATIS, FRITSCH and SAM-
BROOK (1 982).
Growth rates: Growth rates were determined as previ- ously described (PARKER and HALL 1988).
Isolation of RNA: T h e 40-ml cultures were grown over- night at 37' with constant aeration in 0.0 1 % dextrose. After the cultures reached a limiting density of about 10' cells/ml dextrose was added to a concentration of 0.2% and growth was continued until a density of approximately 5 X 1 O8 cells/ ml was reached (this assured that the cells were growing rapidly, thereby optimizing the quantity of mRNA present). T h e cells were harvested by centrifugation, resuspended in 750 pl of lysis solution [50 mM dextrose, 10 mM EDTA, 25 mM Tris (pH 8)] and extracted with 750 pl of phenol at 65". The preparations were centrifuged to separate the phases and the aqueous phases were sequentially extracted with 750 pl of a 1 : 1 mixture of phenol and chloroform, with 750 pl of chloroform, and were ethanol precipitated over- night at -20". The nucleic acids were resuspended in 500 pl of 100 mM sodium acetate (NaAc) (pH 5.0), 5 mM MgCb and incubated with 100 units (5 pl) of RNase free DNase I
(Boehringer Mannheim), for 2 hr at 37". The RNA was ethanol precipitated and resuspended in 100 pl of 10 mM Tris, 1 mM EDTA containing 24 units (1 PI) RNase Inhibitor (Boehringer Mannheim). This procedure yields 60-100 pg of RNA.
Sequence of the cel Operon 457
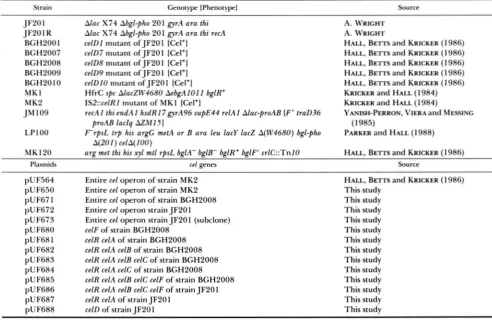
TABLE 1
E. coli strains and plasmids
Strain Genotype IPhenotypel Source
JF20 1 Alac X74 Abgl-pho 20 1 gyrA ara thi A. WRIGHT
JF20 1 R Alac X74 Abgl-pho 20 1 gyrA ara thi recA A. WRIGHT
BGH2001 celDI mutant ofJF2Ol ICel+] HALL, BE- and KRICKER ( 1 986)
BGH2007 cefD7 mutant of JF201 {Cel+l HALL, BETTS and KRICKER (1 986)
BGH2008 celDR mutant ofJF2Ol {Cel+l HALL, BEITS and KRICKER (1 986)
BGH'LOO9 celDY mutant of JF201 ICel+l HALL, BEITS and KRICKER (1 986)
BGH'LOIO celDlO mutant ofJF2Ol {Cel+l HALL, BEITS and KRICKER (1 986)
M K l HfrC spc AlacZW4680 AebgAIO11 bglR+ KRICKER and HALL (1 984)
M K2 IS2::cefRI mutant of M K I ICel+) KRICKER and HALL (1 984)
JM109 recAl thi endAl hsdR17gyrAY6 supE44 relAI Alac-proAB IF' traD36 YANISH-PERRON, VIERA and MESSING
proAB laclq AZM I5 I (1 985)
A(201) celA(100)
LPlOO F r p s L tr p his argG metA or B ara leu lacy lac2 A(W4680) bgl-pho PARKER and HALL (1988)
MK120 arg met thi his xy1 mtl rpsL bglA- bglB- bglR+ bglF' sr1C::TnlO HALL, BE- and KRICKER (1 986)
Plasmids ccl genes Source
pUF564 Entire cel operon of strain MK2 HALL, BETI-S and KRICKER (1986)
pUF650 Entire cel operon of strain MK2 This study
pUF67 1 Entire cel operon of strain BGH2008 This study
pUF672 Entire cel operon strain JF201 This study
pUF673 Entire cel operon strain JF201 (subclone) This study
pUF680 celF of strain BGH2008 This study
pU F68 1 celR celA of strain BGH2008 This study
pUF682 celR celA celB of strain BGH2008 This study
pUF683 celR celA celB celC of strain BGH2008 This study
pUF684 celR celA celC of strain BGH2008 This study
pUF685 celR celA celB celC celF of strain BGH2008 This study
pUF686 celR celA celB celC celF of strain JF20 1 This study
pUF687 celR celA of strain JF201 This study
pUF688 celD of strain JF201 This study
Other cloning vectors: The plasmid pBlu+ is a 3.0-kb high copy number vector that carries a multiple cloning site cartridge and a gene for ampicillin resistance (Stratagene). The plasmid pHSG4 15 (HASHIMOTO-GOTOH et al. 198 1) is a low copy number plasmid which confers resistance to ampicillin, kanamycin and chloramphenicol.
&Glucosidase assays: @-glucosidase assays were per- formed as previously described (PARKER and HALL 1988).
Colony hybridizations: Colonies were lifted onto Gene Screen Plus nylon membranes (New England Nuclear), lysed, and fixed as described by the manufacturer. The filters were allowed to air dry, and then hybridized as described in HALL and BETTS (1 987).
Polymerase chain reaction (PCR) amplification of DNA fragments for sequencing: Amplification of fragments for sequencing was accomplished in two phases. The first phase generated double stranded DNA and the second phase generated primarily single stranded DNA. First phase am- plification reactions contained 100 ng of genomic DNA, 20 nmol of each dNTP, 100 ng of oligonucleotide primer #1,
100 ng of oligonucleotide primer #2, 2.5 units AmpliTaq (Perkin Elmer Cetus) in 50 mM KC], 1.5 mM MgC12, 10.0 mM Tris-CI (pH 8.3), 0.01% (w/v) gelatin in a total volume of 100 pl. The mixture was overlayed with 100 pl of mineral oil and amplified for 15 cycles in a Perkin-Elmer Cetus DNA Thermocycler under the following conditions: denatura- tion: 94" for 45 sec, annealing: 55" for 10 sec, polymeriza- tion: 72" for 2 min.
The second phase generated single stranded DNA and was identical to the first except that 2 pl of the first phase
celR celA cell3 celC celD celF
0 1 2 3 4 5
Kilobase Pairs
FIGURE 1.-Restriction map of the cel operon and flanking re- gion. Sequences outside the cel operon are designated with a thick black line. Abbreviations: A = &aI, C = e l , E = &RI, EV =
-
EcoRV, H = Hindlit, Hp = Qal, S = g l .amplification was used instead of E. coli genomic DNA, and oligonucleotide primer #2 was omitted. Amplification was for 25 cycles under the following conditions: denaturation: 94" for 30 sec, annealing: 50" for 5 sec, polymerization: 72" for 2 min. Samples were purified using a Qiagen t i p 5 according to directions from the manufacturer (Qiagen) and sequenced using oligonucleotide primer #2.
SI transcript mapping: Aliquots of 30 pg of total RNA were combined with 1 X lo6 cpm of ["SIdATP (an analog of dATP that has the a-phosphate replaced with radioactive sulfur)-labeled single stranded probe fragment generated by PCR and dried in a vacuum oven. The mixture was resus- pended in 20 pl Berk-Sharp buffer [0.4 M NaCI, 0.04 M
458 L. L. Parker and B. G. Hall
0 0 0 0 0 0 0 0 0 0 0 0 0 0 0 0 0
0 0 0 0 0 0 0 0 0 0 0 0 0 0 0 0
m
i 3 N 0 N 3 N N m N c N m N P N m N m N 0 m 0 m N m 0 0
0
' 8 8
1 5
,
pj
8 3
E!
8.g
Ii
3 5
. U
E 4
u.a;:
.i4$ 8 %a;:
E d 8 8, e P,
$ 2
8 g , q
$ 3
83
E 3
3:
9;
F:
3;
3 2
'8: 8 '. * P
.
p
W * @ c 3w c a n
' 8 %
8:
i4E
. 8 $
3 3
. E ;
8;
; 2
8 2
8 4
. &
P
:>
$ 2
3 3
w 3
?g
q
*!E
E x
. 3 $
s e
' 2 +2
$ 5
' 8 E: eE %
a;:
8 ;
6 5 8 9 $ 1. * - * o
Y k
' 5 :
e *!j;
8 2' 3 . 8 j u ' s a
3
j p
ig
3
2 2 ; , &'1
, g a rE j
H
$i;
!4
:
2
i
. *
3
#
. q1
' $s
3 2
2;
2
jj
1
I
a 2
9 q
e r;il
$ 3
.;r
.t:
.!;
3 2 :
s
c j
. 8 3
* v i urn. $
g.!;
8;
2
* a$ 2
. E $
, 3
, i ;
u a
3
5 :
F G
H
3 2
8 ;
8:
g
8 %
.I
.I.
. E 2 - 2ox
8
K
a;:
E E *
. E
. 3
, 8 6z
w 8 2. * ' W . w ,
g ;
F
2 -E 4
3 5
$ J $ ' E
.
-
y .1 ru'
t l c
[
@$E
. * *vi Y1 4 1
E.*
' $
z
A
w u
9 E
$ 5
' 9 2 * m
:;
. g :
3':
8 3
2 t8 I
a 1
3 8
U d *yI< .+
!{
' E 3 1 4 m
J j 3
6 "
. 8 5
2;
.4;
!$
u - .;;
* SE % .
5 :
8 2
. 8 8
2 2
. 2 2
E 4
3 3
3;
8 " W' "3. * c 3 a u n
g j
3 2
' 8 3
:':
. S f
:;
6!
3 5
E %3 3
3;
* a 14 .iU T
,141 u w g 4
;i
' 8 %u -
C r n
* m u r l
C I O w -
" >
. C 0 - 1 u g 2
2:
' E t
e *
3;
$ 2
3;
W Y
. w w 8 3
8 %
$ 2
U Y I
w 1 4
'i;
3w w
e . 0
* -
u w
0 3
. 2 5 ' 3 2
3 2
$ 2
' E 2
g:
ii!
z;
2;
. Y Z
. z ;
0 %
3 %
jp
. 3 z
2;
a :
3:
E;
' E ? u o' 5 3 "
. $ $ $ . ? !
e m
[i
,0g3j!ji
3 ;
u * Y .:
u 2 u w
&
$iJ
4j;:
$
1:
g;
[ $
11
" 5 a z
; a $
3 ;
E . u s * Y2!8 * d
. 3 $
. s . u2
3:
8 z ;
u w $ 2E 2
e S
. # . a 8 %
. r e
i?:
C -
8 $
' $ i
3 5
$ 2
E $
, w
-0 2 6 E A
,143 w w
3 ;
* u C S *ai. #
3 E %9 .:
4 .3
UT
, $ ; c v )
* x
8 2
. "
"04U P * w C S
3;
n r e a *A. w > ern
C t l
*.c E % 5 : 8 2
3 ;
;:
w 22:
' E 3
8 % - 2 8 2 0 - * * . # $
2;
. g z
8 8
c -
U P I
* s * a
. E ;
I 2
n m
8 2
Uk U S
, 1 4 4
E : 2.:
Y e r : 8 5
. U " C 9
E {
-0.C S u r n C "
* o . g u "
5 2
E S
8 5
2 2
. w a
u u
U P
w 3
e.*
Ud
.I: :
E::
2 5
u .+
UT
. w w
C U c s C P e u CS C P u o
3 2
. l i
*LI 2: r ' 4 w.I$
* - E :$ 2
. * a * u
4 - U D
5'
8 3
% E
.!4;
c xw - w w c s e.= * a a 2
. C P
e.*
2 6
E:
' 2 2
g %
. 2 3
e."
$ 2
w w * -
e m
c s C c u
2 2
g
i
&
$ 5
8 2
8 % 8 2 - 2. U 0
* s * u * a *vi * k u u -01 * * * - 4 s w 3 . 3 ;
. s <
$ 5
g
8 2E 9 5 2 . C u
. * r
;;
3 :
E % 8 2 e 2 8 % ,pJ W Y2 5
C C ,1433;
p
,i;
[ $
* a $ {$ 2
, z ;
2 E
.4 U.4 I
6
8 %
e.*
' 8 8 2 5
9 2
5 :
"LI
i ;
. * a 1 4 u l * m w e
* -
4 - 0 - E : . 2 5
3 3
.
$ $2;
, $ $
3;
C Y )
u .4
E %
E S
* m
ig
*D
. e 4
U " m u COI CS a - E : $ 2
. 4 0 W Y u * U P
!$
. Y 2g e
E 6 , w 2
P B " 4 C C a x C Y 4 %
8 5
g
$ 2
C L 1
, 3 ' &
E 3
2 2 , E 4
2,.
'Ei
8:
E ?
4 - c u. 4 x
re
u 2
$ 3
*VI
w -
' E e!
g g
E <
. *
3 2
Y. u r l
8 4 8 5 E:
3 :
E ?
*E Y Y . & V I* u
3 :
$ $ ' 8 2 8;
2 8
C *
c s CP
, u - w m w 4
e o
-3
$ 2
5 :
. P B o u w -
ern
8 ;
' 3 2
C v i C Y $e?
U L I
u s 5 : :
. I 2
Y- 3
. r l
I!
urn
g ;
2;
e S
* P .Cc 2 z 8 2 W Z C o
$
c c2;
5 5
e - . u ne o
* "
5 ' ;
2 3
' E
,ii!
e $ 0
I
* ;1;
. E
. z
*2 L
8 5 2
, 2 2 5 . *
8.2
8
3:
E
$ $
3
2:
4
. *
3:
0, H
c r
w w C C k .C Y . 5
w y l
. 8 2 . U P
3;
8
2 2
4
CU
g
' E X ' $
3 2 f
- 3
22;
6
v u
"LI u a
u * 8
P E E t E
* s u
I 1 Y
&
. $
. g
,i
C U
&
E
E % ::
C O
a
. g z
z-2
. P E C " 2C a
S d
s
15
$
3 3 3 3 3 4 + i
-
-
-
i-
3 3 3 30 0 0 0 0 0 0 0 0 0 0 0 0 0 0 0 0
P m 0 3 N m c m m P m m 0 i N 0
3 3 3 N N N N N N N N N m 0 rn m
0 0 0 0 0 0 0
0 0 0 0 0 0
- w m
0
0 0 0 0 0 0 0
0 0 0 0 4
3
-
3 3 30 0 0 0 0 0 0
0
N m c m m P
3 d 3
0 0
w rn m P m m
' E ' E .Y.: ' 2 %
' $ 5
' 4 3 -3 3
, 8 > 23 p
.;; . ) j 2 ' 6 % ' $ 2 .3 2
'2;
. g <
' $ 4
' $ 2 ' 3 ;F
8 5
is
4:
E 3$ 3
$ 5 ;
2 3
E~3 $
2;:g a
8 4 ':;i
C a $ 28 %
g ;
8 3
E $
u 48;
8;
ii
g $
E 2. E
. z :
. g :
, 4 . p. E $
. $ $.El
. 8 2. E ?
g
:
u r l E d
h
8 2 E S 37.3
L$
6 Z 4;!$
E
3;
5 :
g t
E <
5 :
3;
. E 2E!
,];
,:z. 8 2
U L I
2;
a;:
e*I:
* P ; 2 " 0 1 2: E d w 2
E3
8 ;
2;
B E
'$5
$ 3
2: E tE;
. E 7 4
.sLa
.I!
.!.
.:Z . E 9 . 8 J J . 2 %. s t
' U @ * *, E ;
, 5 ;
, E ;* u 8: vi
r@
[ i
$!
U8 t Y E 22;
*vig g
g $
*VI , $ !1%
!$
F S
F:
:: u d $ 2 C " k@ w y *ir C r lg
i
8':Hi
&j
:: * " E $ 8:i $
g.;x
E ;
82
g;g
. D
. Y . E . p. z ;
u 3. E
. 3 1
, E $
.!j$
, 3 $
, $ ;. i ;
. 5 g
' 3 ;
$ 2 . j $. 8 z
E
[
E 2;@
!g
1
&
8;
$ g
;#
E74
8 d$ 9 9
i;
8 ;
Ei
!$
c m * mE E: 8 2 2 s
5 ;
g
* v ) up1g g
@
8 5
. S
. *a
2
' g g
' 8 8 ' E :' 5 :
2 : . 5 s 8': . * 3. $ 3
. E $. g j
.ig
. 2 2
.I%
.;s
g
[
.E
p
gg
I-:
f g
g j
a >
E $E-"
* * 2:5
E , $b
, f d
145 * I * X'[
8 2
E!$ 1 E $
*f a
3Bj
2; * t$ 2
* *. 8
. g . E-
2 . $ $ .&j. z ;
. a $
. 2 z 3 ;. E ;
. $ % . p . g. E 3
Y
[
8
"021;
.;!
2
$ 5
u 4 - E B E 3 * w;;
g j$ 3
$!
y12 2
8 : 8 2E $ B 8:: E 3 :
1;
g
i;
, @3:
8 2
8 s 8 23;
K z
$ $
Bj
. E
' 2
' 4 7 E 48:
1%
E
[ j
ij
E j
8 3
5 :
y ;
i;
j j
ij
L&.;: . $ e . 2 .6 . * a ' 8 s w $
' 5 ; . 5 :
: [ E
E $
U U 8 2 2 E ; 5$ 5
, E $
5
. g e
2;
. f5 :
* a U r n 2:. 3 $
. g 5. 8
E
' 2
*
-I.
g ;
8 4j
$ 2
3. g z
. @. g j
. E $
4;
. I 4
. $ ; 8 :8
z
4 '
g 3
3%
3 $
8 : c1 8 3E {
E S$ 4
z gi$
2 3
3 5
[ j
8 j
8 3
* @
E
, E $.r
'1
. $. j ;
. g .*;: . C . Q . $ $E ?
2 2
. E ! . # . Q5 %
. E $
. E a3s
.k$
,3
!j
8
E !
kj
8 2
g
Q
j 2
E d
8 1
8 i
:E
i2
E)&
g:
E $
g $
. f
. 8
. E % . b. @ .!j
. E $
u 4, g 3
,i$
. I s
. g z
, # $
, J , : g e d4 %
E $
8 ;
E.2 2;: 8 2g z
E 's z
' 3 ;
E
8
'I?
E;
i$
8 8
E $
$;
is
$ 4
o 2
E 3 -10 8 % -6 2 4g g
l $
9:
KS
! $ E
s a
E
3 8
3 2 * a E;: E $ * * Y 2 , 5::2 8
6 %
I
.I
.!&
c 3g g u o 4 E % E :
8
Y
E
. E
. p
, 8
?
E
8 . 8 3
w o
ucu u x C P
1 4 x
Y w 5 ::2
U O 3 2
* a Yd 2.2 e a
c
E S
8 0 E s S IF
3 ;
8
w - , 8 2 * m . 2 ; .*4 . 8 s . g 2
* a I
* o * c *vi
* P
$ 2
* s " Y% a * 8 % c S
e
2
F E
- 4 .f E 8 %8 ;
g 2
g $ E ;E!
- 3 4 3 3 4 3 3 3 3
-
n-
d 3 3 30 0 0
3 i
-
3 3 3 30 0 0 0 0 0 0 0 0 0 0 0
Sequence of the cel Operon 459
0 0 0 0 0 0 0 D 0 0 0 0 0 0 0
0 0 0 0 0 0 0 0 0 0 0 0 0 0 0
m Lo r m 0 d N m
-
m w r m m0 0 0 0 *
-
w * *-
F * - r e- 3 "1: G 9 8: 2 2
$ 2
8 2
3:
C - lE : 2 : 5 2
c m
[ $
* w e
'
dW
e s e o * P 4 2
$ 2
E ;
2 2 4 ;e . 0
e - 1
LY,
"0
u u u c r
U Y
gi
* E $ 8 8 2 9 8 8$!!E
U Y 8; 6:: 2 6 2: 8 3 e * e '
!$
* i
I$-
i. e
!.
e 3
5,"
1 2
rlL l u
' 3 6
.;; "Y "0. "3 * m w >
: j
h
. e e - v 143 w u 4 2 2 8 8 t
3 ;
8 5
5i
!;
. 5 4 e a
* u
e 3
c u e 4
. * & de dm a e
E 2
3 2
33
3 ;
V ";!
0 0. U u
* x
w -
, 8 ;
, E '
E :
E?. e m w > 2 % 2.;. u a
C O e 4 €.m a n
@
8:
. 8 E
$!
) $
E5
* '8 2
e m . " u! I
&5
. 9
G 9
. U m
2 2
M e
2 6
. U T :
< e
C Y
c -
C r n
" >
u o
-1:
. e o * E.&
3 3
8 2 e &.!$
3 2
1%
?g
0 3 E 9 5g:
8.2,' 8 :
8 4
rl . e " > 4
e a
!I
. E 9
E ? !
4 2
4 2
c u
, e @
c s *cr - 4 < n " 4 e 9 " > " 0 u u
. e 3
u u
U T 4 *
ern . U r l
" 3 *(u e x :% a H e 2
43
c u -1:. G b
8 % 8; $; .;:
3 2
" 48 5
.ig
" 0 $ 2 :E 8 d E 3. C 3
:E
.
4:i
.+i
s
E 3
4 0
u u
. u @ * w C T * a U D 4;1
.ig
€ . u c s ccr
$ 3
B S
g <
U Y. 2 6
4 .2 2 %
5,"
, e 3
$ 5
. U T : - 4
w >
"Y
8 1 : 5 2
i g
, " "rng g
u a3 2
3 :
. e il
u o )
c s
* a
2 6
, C - c w
s z
8 %
* *g:
3 2
. a w o v u "0. *ul w x * u. * 1
- 3 2 2
$ 2
0 "
U '
. " I
3 2
" 2 e a.I$
@
" 2 c d 4 2 2; " a . e -" @
C U C V I s t 3 e r l
e a
, g z
4 3
9 8
2 $ , i j ;
1 ;
. e we -
u - l
$ 8
E 9
s t
. 8 2 0 4
u r n
e u e - * u c s ccr W Y a e 2 8 $ 2
2:
' 8 2
2 8
.e.- u u
u a
e . 0
* -
4 u
. g j
W '
w x
* u
8 3
E 9' $ $
" > c u u u *VI v u. e m u u o m
i j
-0
e - 1
. " c
9;
3 ?
.;: "U e m 0 - e m 2 % 8i; 2 2 c 3' w a *3
* a
" 3
c w
3 5
' i 2$ ?
e o e s
e a
u t "
. U Iu
g
, $ j
A2$
$ 2
u x w - 0 " e - -0 4'; g 2 $ 5.!;
-a r 4" 4
" 0
GJ
2 2 2 :.
' 8 V - l 2
W Y
3 8
, $ -l
8 2
U "
$ 5
3:
. e a
u e e s cL. " 3 e @ " 2 2 2
, E 3
5 2
E ; c u
u c
*VI
. " X e m
3:
E:
$ 4
8 %. E 3
E 3 e 2 * c "4 u u *VI
. e 0.
e o e -
2 2
.;;
3:
e - l
2.: z z c s .*cr 6:
E
* a. u @I
4 m w 4 e s ecr e o e 3
$ 2
a -' s ;
c 3 2 2 " 0
c o
U Y
u n .
'1;
-lc i 5 2
4.:
" 2
. * w
u - l
c u
c s
" "
e a
. E 2
3.2
2;
3 5
" 2
. u -
e m
4 d
"C
6 3
. 5 ;
2 5 8 2
3'
' $ $ " > U "8;
2;
UX "i. u 0 " x
"3
v u 5 2
. 2 2
8 2
8 9
3 2
0 "
. $ j
o w
e o C 3
a -
2:
. a ..
" I
2:
a 4
;;
. U "
$ 2
3 3
u a
3 ;
. " u
4 x c e u o c - 4 e e4 ::a
. 5 ?
U Q
e -
cio
" >
. o w
" x
c u V Y I
5 2
2 '
4 . 2
$ 2
5 1
9 2
. e a
c i u u cVI U L < > e *
e i i
' d 2
8 2
*Io
8;
e x
0 - ' 8 B
e a 4.5
3 2
3 23 2
$ 5
" 0. " 3
" 3
3 3
. U il w o
v c r
e 4 * m
' 4 2
U L
40
i i
, v - o m
" -
2 E
8 %
E X , $ a
c 3
a m
" 4
u t "
" X
"4
" 0 " 2
8 5
. e YI
" X c u u u 2 6 .
!$
3 ;
" > 4 m8 %
. * e
!.!
i i g
( 1 4
v u
"0 u a
. c e - m
5 :
8 ; ' Z Z
$ 2 8 %
s i
e @ 0 - " 0 e10. U Z 4 .*
U L
"0
U "
U & 4 n
2 .:
:?! . $ $ e m " >
$ 2
e m. g . U
a !
"
j
, g
::3
;
. u
: : :
. C$
$
. U
3
, [
3
:
iz
$
3
" "
. " ' 8
:: "
2 E
4
c
e W
. e . "
U 2
2 s 2 y g
;
;
Y E c. " . U
$
g
3
;
. e . C; !
. 4 . *
2 : :
. $ S $ 8 0 .:: 2
- : 8 8
J
E
a s ,
3
g + @
0u c r
3
v u . e U W e c . 4
3
U . e c2
8 8 2 " . e U$
8 ' 2 4 4 U . ";
4 :: . UI
. 6
4 4 . 5 e U C 4 W . e c[
2 . E Y. 2 4
2
. s
8 u c c c W;
" ' 2 4 2 E 2. E 2
8 e " . w * a W
9
. " I " U 0 . " 24
8
Y
, 3
3
. e
51
~ oi
: g
m
--
._
y :
F
,%. E s
Y =
2 2
5 %
' 4 4
-
3E t
4 %
Y " ,
iL
' E 0
d
-2 %
.:: r
E *
c x
" -
E ;
" 3 ,
" 3 ,
$ 5 . U -3
$ 3
$ L
.I
F;0 . .
. 5 u .E r
0
-g >
e
-u 5
$ 1
, E
3 L i
5
G i :
S L 2
e
3 3 3 3 3 3 d d
0 0
-, r, w P m 0' 0 i N 0
m m m m rn
-
w-
*3
-
0 0 3
-
4 3 - 3 3", 0 0 0 0
m r m m
v
-
T " "460 L. L. Parker and B. C. Hall
containing 80% formamide. T h e mixture was overlayed with 20 pl of paraffin oil, heated to 90" for 3 min, cooled slowly to 50" and allowed to hybridize for 3 hr at 50". Aliquots of 200 pI of S1 buffer [0.25 M NaCl, 0.03 M NaAc (pH 5.5), 0.0005 M ZnSO,, 20 pg/ml sheared, denatured salmon sperm] were added directly to the hybridization mixture, and S1 nuclease digestion was carried o u t using 100 units of S I nuclease for 30 min at 37". The reaction was terminated by extraction with an equal volume of phenol followed by an equal volume of chloroform. T h e mixture was precipitated with 2 volumes of ethanol and resuspended in loading dye. T h e fragments were separated on a 6% acrylamide sequencing gel.
PCR amplification of the probe fragment was accom- plished using 30 ng of Hind111 digested plasmid pUF687 (this study) DNA, 20 nmoles of each dNTP, including 5000 Ci/mole ["SIdATP, and 100 ng of an oligonucleotide primer corresponding to the complement of bp 300-324 of the cet operon in a total volume of 100 pl under identical conditions to those for stage two of DNA amplification for sequencing. Hind111 digestion of the plasmid ensured that the PCR reaction terminated at the Hind111 site at bp 1 of the cel operon. T h e fragment was not purified before use.
DNA sequencing: Fragments for sequencing were sub- cloned from plasmid pUF650 into plasmid pBluC and se- quenced directly from the double stranded plasmid, using the dideoxy chain termination method according to SAN-
GER, NICKLEN and COULSON (1977), except that instead of alkaline denaturation of DNA fragments, samples were boiled for 5 min, frozen in a dry ice/ethanol bath, and allowed to thaw at room temperature.
Creation of unidirectional deletions: Deletions were cre- ated with the enzymes exonuclease 111 and S1 nuclease using the Erase a Base kit according to directions provided by the manufacturer (Promega).
Protein comparisons and analysis: Proteins encoded by
celF and bglB, and celB and bglF, were compared on the basis of amino acid similarity using the programs Gap (DEV-
EREAUX, HAEBERLI and SMITHIES 1984), Align, FastA and RDF2 (PEARSON and LIPMAN 1988). Structural analysis of
celD was determined using the program Pepplot (DEVER-
EAUX, HAEBERLI and SMITHIES 1984) and the parameters of
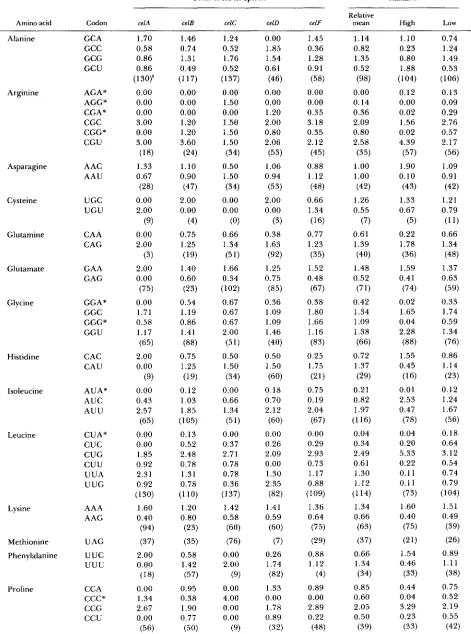
CHOU and FASMAN (1978). T h e hydrophobicity plots were constructed using the program DNA Strider (MARCK 1988) and the parameters of KYTE and DOOLITTLE 1982. Codon usage analysis was aided by the program DNA Strider (MARCK 1988).
RESULTS
Cloning and sequencing the cel operon: A partial
restriction map of the cel operon is shown in Figure
1 . Previous results (HALL, BETTS and KRICKER 1 Y86),
together with additional unpublished data, indicated that the entire cel operon of the Cel' strain MK2 was contained within a 7.7-kb EcoRIIBamHI fragment. T h e cel operon from the strain MK2 (KRICKER and HALL 1984) was therefore cloned in vitro as follows: genomic DNA of strain MK2 was digested simulta- neously with the enzymes EcoRI and BamHI and li-
gated into the EcoRIIBamHI sites of plasmid pHSG4 15 (HASHIMOTO-GOTOH et al. 198 1). This low copy number vector was used because it had been previously shown that the active cel operon is selec- tively disadvantageous when carried on a high copy
number plasmid (HALL, BETTS and KRICKER 1986). T h e ligation mixture was transformed into strain JF201 R and the transformation mixture was plated
onto arbutin minimal medium. One colony was iso- lated, shown to be cellobiose, arbutin, and salicin positive, and its plasmid, carrying the MK2 cel operon, was designated pUF650.
For purposes of comparison, the wild-type (cryptic) allele of strain JF201 was cloned in viuo using the Mini-mu system as described in GROISMAN and CASA-
DABAN (1 986). Strain JF20 1 was lysogenized with both Mud5005 and M u cts62 pAp5. A genomic library was produced by thermal induction of the dilysogen, in- fection of the Ace1 strain LPlOO (PARKER and HALL
1988) with the lysate, and selection on rich medium containing kanamycin. T h e colonies were lifted onto a nitrocellulose filter, lysed, fixed, and hybridized with the 4.0-kb HpaI fragment of pUF564 which had been labeled with [35S]dATP (HALL, BETTS and KRICKER
1986). A plasmid from one of those colonies included 12 kb of insert DNA, contained the entire cel operon, and was designated pUF672. The 5.5-kb EcoRIISalI fragment from pUF672 contained the entire cel ope- ron and was ligated into the EcoRI/SalI sites of the high copy number vector pBlu+ to generate plasmid pUF673. T h e 800-bp HindIII/HpaI fragment from pUF673 that contained celR and celA was subcloned into the HindIII/EcoRV sites of plasmid pBlu+ and designated pUF687.
T h e cel operon of strain MK2 was sequenced by subcloning fragments from pUF650 into the sequenc- ing vector pBlu+, and sequencing directly from the double stranded plasmid using chain terminating di- deoxynucleotides according to the method of SANGER,
NICKLEN and COULSON (1977). Both strands of the entire coding region were sequenced from the active allele carried on pUF650, with the exception of the HindIII/HpaI fragment (bp 1-801) that contained the insertion sequence IS2 inserted between bp 166 and
167 of the operon (PARKER and HALL 1990). T h e nucleotide sequence of both strands of this fragment was determined from the wild-type (cryptic) allele carried on pUF687. Only one strand of the HindIIIl
HpaI fragment was sequenced from strain MK2. Base
pairs 166-901 of the MK2 operon were amplified through polymerase chain reaction using oligonucle- otide primers corresponding to bases 70 to 51 of IS2
(GHOSAL, SOMMER and SAEDLER 1979) and to the complement of bases 902 to 921 of the cel operon.
N o sequence differences between the wild type and active alleles of the cel operon were found in this region.
The cel operon contains five open reading frames:
Sequence of the cel Operon 46 1
I S 2
S HD
.<....,....,.<.,.<.,.,
d
pUF671celR celA celB CelC celD celF
IS 2
, . . . . pUF683
celR celA celB CelC
E & -\
' pUF682
$
&A,
st st C A p U F 6 8 4celR celA celB
pUF689
celR celA
celR celA celB celC
FIGURE 3.-Restriction maps of insert D N A in plasmids carrying
various portions of the cel operon. Abbreviations: A = A w l , C =
C l a l . E = EcoRI, E\' = EcoR\', H = Hindlll, Hp = Hpal. S = Sall. restriction map (Fig. 1). Sequence analysis indicated that the cel operon consists of five open reading frames, designated celA, celB, celC, celD and celF. Up- stream of celA lies a 256-bp region of DNA that we have designated celR. This region contains the pro- moter, putative operator, and the 108 base pairs where the activating insertion sequences have inte- grated (PARKER and HALL 1990).
Localization of the transcriptional start site: We
have localized the transcriptional start site of the cel
operon of strain MK2 using the RNA protection method described by BERK and SHARP (1977). One major fragment was present and indicated a primary transcriptional start site at the guanosine at bp 238 of the cel operon (see Figure 4 of PARKER and HALL 1990). That guanosine is located 49 bp prior to the first nucleotide of celA. Several reproducible minor fragments were observed, indicating that additional transcriptional start sites may be present. Alterna- tively, these fragments may result from RNA degra- dation.
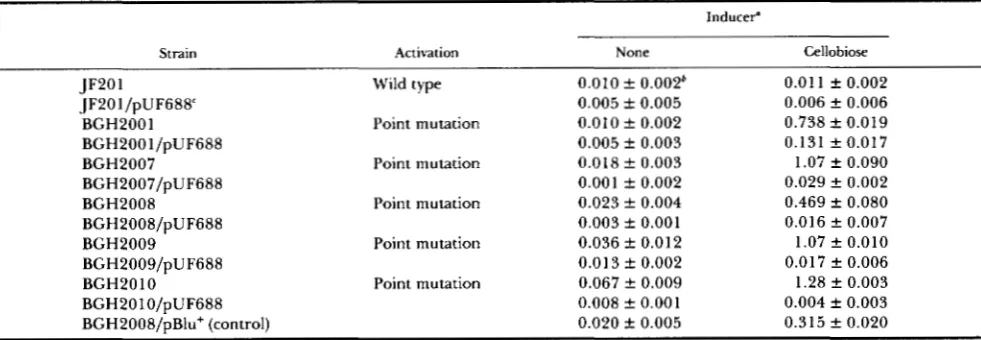
Functional analysis of the genes of the eel operon:
celA: The first open reading frame, designated celA,
begins at base pair 256 of the cel operon, ends at base pair 606, and encodes a 1 1.4-kD protein consisting of 106 amino acids. T h e function of this protein is not known. We have isolated RNA, spotted it onto a nylon filter and hybridized with ["SIdATP labeled celA spe- cific fragments. Autoradiography indicated that celA
is transcribed. Several lines of indirect evidence sug- gest that it is translated. First, celA has a Codon Adaptive Index (CAI) of 0.341, much higher than the
TABLE 2
&Glucoside transport activity conferred by plasmids carrying genes of the cel operon
Plasmid"
pUF67 1 celRAUCDE 0. I78 f 0.004
pUF689 celRA 0.016
*
0.008pUF682 celRAU 0.4 14 f 0.006
pUF683 celRAUC 5.17 f 0.240
pUF684 celRAC ( A U ) 0.01 2 & 0.004
pBlu+ Vector only 0.01 8 f 0.009
Functional cel
genes Transport activityb
a Host strain was LP 100 (Abgl Acel).
*
Transport activity is expressed as nmol of PNPG hydrolyzedmin" A,& f 95% confidence limits in dextrose grown intact cells (see text).
value of 0.17 that is expected for a random collection of nucleotides (SHARP and LI 1986b). Second, the codon usage profile of celA is consistent with the other genes of the cel operon. Third, celA is preceded by the potential ribosome binding site (SHINE and DAL-
GARNO 1977; STORMO, SCHNEIDER and GOLD 1982)
GAGGG, 7 bp before the initiation codon.
celB and celC: celB begins at bp 69 1 of the cel operon,
ends at bp 1944, and encodes a 418 amino acid protein with a molecular mass of 44.95 kD. celB is separated from celA by 83 bp and is preceded by 9 bp with a translational initiation site sequence AAGGG.
celC begins at bp 2089, ends at bp 2439, and encodes
a 1 17 amino acid protein with a molecular mass of 12.75 kD. celC is separated from celB by 153 bp. A ribosome binding site, of the sequence GAGAGGAA, is present 7 bp prior to the start of celC.
It has been shown that integration of the transposon
7-6 into a region of the cel operon now identified as
celB resulted in the loss of the ability of a cloned Cel+
allele to ferment the 8-glucoside sugars cellobiose, arbutin and salicin (HALL, BETTS and KRICKER 1986). However, integration of 7-6 into the same Cel+ plas- mid in a region now identified as celD and celF resulted in the ability to transport, but not hydrolyze, the
8-
glucoside sugars (the transport activity was measured biochemically, independent of the ability to hydrolyze 8-glucoside sugars) (HALL, BETTS and KRICKER 1986). These data indicate that regions proximal to celD andcelF are responsible for the transport function. This
implicates either celA, celB, celC, or some combination of these genes, in the phosphotransferase system (PTS) transport of 8-glucoside sugars.
To identify which gene(s) specified the phospho- transferase protein, several plasmids were con- structed. Plasmid pUF681 contains celR and celA,