Original Article Expression of PD-L1 in mononuclear cells, multinucleated cells, and foam cells in tenosynovial giant cell tumors
Full text
Figure
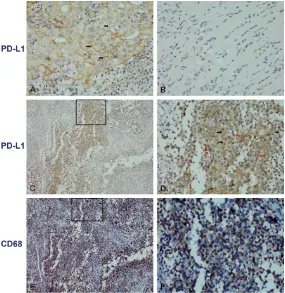
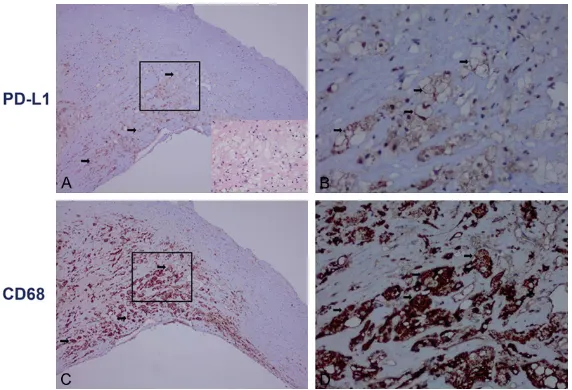
Related documents
Monitoring PD-L1 positive circulating tumor cells in non-small cell lung cancer patients treated with the PD-1 inhibitor Nivolumab. Association of a putative regulatory
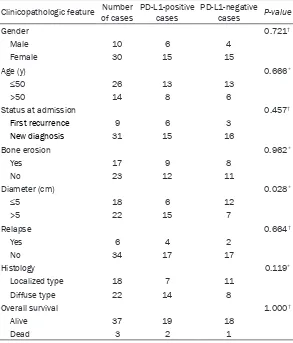
The positive expression rate of PD-L1 in colorectal carcinoma tissues was not related with age, gender, tumor size, differentiation, TNM clinical stage, or depth of invasion ( P
In patients with advanced NSCLC and PD-L1 expression on at least 50% of tumor cells, pembrolizumab has been associated with significantly longer progression-free and overall
Keywords: PD-L1, oral squamous cell carcinoma, gene expression, circulating tumor cells, survival Received: February 23, 2015 Accepted: May 05, 2015 Published: May 15,
Furthermore, higher Exo-PD-L1 content was associated with larger tumor size, positive lymph node status, distant metastasis and advanced TNM stage in NSCLC patients.. However,
of 111 patients had positive PD-L1 expression either on membrane or in the cytoplasm of tumor cells, and there was a positive correlation between the expression of PD-L1 and
High expression of PD-L1 in lung cancer may contribute to poor prognosis and tumor cells immune escape through suppressing tumor infiltrating dendritic cells maturation..
This study investigated the prognostic significance of PD-L1 expression on tumor cells and tumor-infiltrating immune cells (TILs) in the tumor microenvironment in Chinese
