On
the
Evolution of Genetic Incompatibility Systems. VI.
A Three-Locus
Modifier Model for the Origin
of
Gametophytic Self-Incompatibility
Marcy
K.
Uyenoyama
Department of Zoology, Duke University, Durham, North Carolina 27706
Manuscript received November 18, 1990 Accepted for publication February 14, 199 1
ABSTRACT
Recent genetic analyses have demonstrated that self-incompatibility in flowering plants derives from the coordinated expression of a system of loci. T o address the selective mechanisms through which a genetic system of this kind evolves, I present a three-locus model for the origin of gametophytic self-incompatibility. Conventional models assume that a single locus encodes all physiological effects associated with self-incompatibility and that the viability of offspring depends only on whether they were derived by selfing or outcrossing. My model explicitly represents the genetic determination of offspring viability by a locus subject to symmetrically overdominant selection. Initially, the level of expression of the proto-S locus is insufficient to induce self-incompatibility. Weak gametophytic self- incompatibility arises upon the introduction of a rare allele at an unlinked modifier locus which enhances the expression of the pr0to-S locus. While conventional models predict that the origin of self-incompatibility requires at least two- to threefold levels of inbreeding depression, 1 find that the comparatively low levels of inbreeding depression generated by a single overdominant locus can ensure the invasion of an enhancer of self-incompatibility under sufficiently high rates of receipt of self-pollen. Associations among components of the incompatibility system promote the origin of self- incompatibility. Enhancement of heterozygosity at the initially neutral proto4 locus improves offspring viability through associative overdominance. Further, the modifier that enhances the expression of self-incompatibility develops a direct association with heterozygosity at the overdominant viability locus. These results suggest that the evolutionary processes by which incompatibility systems originate may differ significantly from those associated with their breakdown. The genetic mechanism explored here may apply to the evolution of other systems that restrict reproduction, including maternal-fetal incompatibility in mammals.
S
ELF-INCOMPATIBILITY in flowering plants and functionally similar forms of maternal-fetal incompatibility in mammals derive from genetically controlled interactions during reproduction which modify the distribution of genotypes among offspring. A growing body of evidence indicates that reproduc- tive incompatibilities of this kind derive from the coordinated expression of a system of genetic loci. This multilocus view differs from the conventional model for the origin of self-incompatibility, which entails the replacement of alleles permiting self-fertil- ity by fully functionalS
alleles encoding all physiolog- ical effects, including the determination of pollen specificities and the recognition and rejection of pol- len expressing specificities in common with the stigma or style (see, for example, CHARLESWORTH 1982). Because components of genetic systems undergo ex- tensive functional and physical integration during ev- olution, the selective pressures that maintain refined systems may differ fundamentally from those that favored their origin. The multilocus approach adopted in the quantitative model presented here permits the direct investigation of the evolution ofGenetics 128: 453-469 (June, 1991)
associations among components of incompatibility sys- tems which are fundamental to their origin and re- finement.
Genetic mechanisms of self-incompatibility in
flowering plants
M. K. Uyenoyama
tion of the specificities. HARINC et al. (1990) have summarized recent findings relating to the physiolog- ical process and molecular basis of self-incompatibility.
Glycoproteins associated with self-incompatibil-
ity: CORNISH, ANDERSON and CLARKE (1 988) reviewed
studies which have characterized to some extent the pistil proteins associated with self-incompatibility. In the best studied systems, including Nicotiana, Bras- sica, and recently Petunia (AI et al. 1990) and Solanum (XU et al. 1990a, b), S-allele-specific glycoproteins which inhibit germination of pollen grains or elonga- tion of pollen tubes have been isolated and their cDNA sequences determined. In other genera, includ- ing Lycopersicon, Prunus, Lilium and Trifolium, the presence of glycoproteins associated with incompati- bility has been established, and in some cases shown to segregate with S alleles in crosses or to parallel the onset of the expression of self-incompatibility during development.
In situ hybridization of a cDNA probe for the gly- coprotein in Nicotiana has indicated that transcripts are present throughout the tissues encountered by a germinating pollen tube, from the stigma, through the transmitting tissue of the style, to the epidermis of the placenta (CORNISH et al. 1987). Antibodies specific for the protein bind to the same regions (ANDERSON et al. 1989). In Brassica, transcripts for the S locus protein are present only in the papillar cells at the surface of the stigma (the site of inhibition of incompatible pollen grains in most sporophytic self- incompatibility systems), and not in other cells of the stigma, style, or ovary (NASRALLAH, YU and NASRAL-
LAH 1988). In both Nicotiana and Brassica, the ap- pearance of the transcripts during the course of de- velopment coincides with the expression of self-incom- patibility.
S locus proteins in gametophytic systems appear
to be ribonucleases: Although cDNA sequences cor-
responding to different S alleles in Nicotiana alata
show high heterogeneity, certain regions are com- pletely conserved (ANDERSON et al. 1989, KHEYR- POUR et al. 1990). These conserved regions show homology to fungal ribonucleases (MCCLURE et ul. 1989). In vitro studies have shown that S locus proteins exhibit ribonuclease (RNase) activity on yeast RNA (MCCLURE et al. 1989) and on ribosomal RNA in pollen tubes (MCCLURE et al. 1990).
, Specificity may derive from the ability of pollen tubes to inactivate or avoid the uptake of RNases encoded by different S alleles, while remaining sensi- tive to RNases encoded by the same S allele (MCCLURE
et al. 1989,1990; HARINC et al. 1990). S locus proteins exhibit less than complete specificity in their effects on pollen tube growth in vitro, causing stimulation as well as inhibition of incompatible pollen tubes (HAR- RIS, WEINHANDL and CLARKE 1989; JAHNEN, LUSH
and CLARKE 1989). In contrast, in vivo RNase activity exhibits high specificity, with ribosomal RNA in in- compatible pollen tubes growing in intact styles undergoing selective degradation (MCCLURE et al.
1990).
Inhibition in sporophytic systems: Glycoproteins
encoded
by
S alleles in sporophytic systems may not act as RNases, as cDNA sequences derived from Bras- sica bear little homology to those from Nicotiana (TAKAYAMA et al. 1987). In Brassica, incompatible pollen grains fail to extract water from the stigma, an event necessary for germination. SARKER, ELLEMAN and DICKINSON (1988) demonstrated that treatment of the stigma with drugs that inhibit either protein synthesis or glycosylation of proteins permits hydra- tion of incompatible pollen at rates equal to or greater than those of compatible pollen. Pollen grains exposed for 3 hr to an incompatible stigma hydrate more slowly after transfer to a treated stigma. This inhibi- tion can be reversed by holding the pollen grains for three hours on a neutral surface such as a glass slide. SARKER, ELLEMAN and DICKINSON (1988) character- ized sporophytic self-incompatibility as the expression of a balanced physiological system having the potential for a range of levels of compatibility through the dynamic adjustment of active components in both pollen and stigma.Modifiers of incompatibility: Modifier alleles seg-
regating at loci distinct from the S locus itself can confer pseudo-self-compatibility [see review in DE
NETTANCOURT (1 977) Chapter 41. In Brassica olerucea,
a suppressor allele at a locus that is not absolutely linked to the S locus permits self-fertility by reducing the concentration of the inhibiting glycoprotein in the stigma (NASRALLAH 1974). Because pollen grains pro- duced by a pseudo-self-compatible (PSC) plant are expected to retain their full function under this mech- anism, plants sharing S alleles would be expected to be compatible if the self-compatible plant is used as the female parent (PSC X SI), but not in the reciprocal cross (SI X PSC). This pattern was in fact observed in several studies in which pseudo-self-compatibility was attributed to non-allelic modifiers
(NASRALLAH
and WALLACE 1967; TAKAHASHI 1973; NASRALLAH 1974; HENNY and ASCHER 1976; FLASCHENRIEM and ASCHER 1979). Observations of the degree of self- fertility in offspring from crosses between PSC and SI plants indicate that modifier alleles that enhance self- incompatibility may express any level of dominance over suppressors (TSENG 1938; HENNY and ASCHER1976).
(SC X SI). This pattern agrees with that observed in crosses involving pseudo-self-compatible plants, sug- gesting that compatibility in both cases may arise through the suppression of inhibitory factors in the stigma or style. In agreement, MCCLURE et al. (1989) reported that Nicotiana tabacum, a self-compatible spe- cies which shows this form of unilateral incompatibil- ity in crosses with N. alata, does not produce the glycoprotein associated with self-incompatibility and shows little RNase activity.
MATHER (1 943) crossed Petunia axillaris (SC) and
P . uiolacea (SI), two species that show the common
pattern of unilateral incompatibility (SI x SC), and analyzed the segregation of self-compatibility in the progeny. Finding that loci distinct from the S locus itself influenced self- and cross-compatibility, MATHER suggested that self-incompatibility in flowering plants may have arisen not through the substitution of S- alleles of full function for S alleles with incomplete function, but rather through the evolution of nonal- lelic modifiers. Under this view, S alleles represent “switch” genes that induce a cascade of physiological responses, each of which is under polygenic control.
BATEMAN (1 952) proposed further that some number
of loci may have been recognized at one time as “switch genes,” with the apparent monogenic nature of S locus systems itself the product of evolution.
Genetic transformation: MOORE and NASRALLAH
(1 990) described the transformation of the self-com- patible N . tubacum with an S-locus gene from B. ole-
racea. Transcripts and the protein product of the
Brassica gene were detected in tissues and at devel- opmental stages more typical of the gametophytic system of Nicotiana than the sporophytic system of Brassica (MOORE and NASRALLAH 1990; KANDASAMY
et ul. 1990). While the introduced gene integrated stably and was expressed, transformation did not con- fer self-incompatibility. MOORE and NASRALLAH
(1 990) suggested that the self-compatibility of trans- formed plants may have reflected the absence or abnormality of expression of the S locus in another tissue. Whether the failure of the transformation to confer self-incompatibility reflects a deficiency of the component in pollen or pistil, this experiment dem- onstrates that expression of self-incompatibility re- quires factors in addition to the S locus itself.
The origin of incompatibility systems
This brief summary of recent physiological and genetic work supports the view that self-incompatibil- ity derives from the coordinated expression of a sys- tem of genetic loci, in agreement with the Mather- Bateman hypothesis for the origin of self-incompati- bility. T h e three-locus quantitative model analyzed here addresses the nature of associations among inter- acting genetic components, and their role in the origin of incompatibility.
Three components of incompatibility systems: I
have suggested that self-incompatibility systems in flowering plants and maternal-fetal incompatibility systems in mammals evolved to serve a common func- tion, the improvement of offspring quality (see UYE- NOYAMA 1988a, 1991). Because many characters that determine offspring quality, including viability and fertility, are not expressed sufficiently early in devel- opment to permit parents to redirect investment to-
ward the best offspring, parental discrimination may have evolved to respond not to offspring quality itself, but rather to distinct characters which show correla- tions with offspring quality. This hypothesis suggests that the recognition of the
S
locus as the switch for the expression of self-incompatibility reflects its suit- ability as a character which both can be assessed prezygotically and shows associations with offspring quality. A minimal incompatibility system comprises three kinds of loci: loci that affect offspring quality, antigen (switch) loci that are correlated with offspring quality, and modifiers that recognize the antigen loci as cues for the expression of incompatibility.Reproductive incompatibilities in mammals: Ma-
ternal-fetal incompatibility in the house mouse may have evolved under a similar selective process (UYE- NOYAMA 1989a, b). Within the context of the present discussion, the t-complex, at which strong segregation distortion maintains deleterious t-haplotypes, repre- sents the viability locus, and the major histocompati- bility complex (MHC) the antigen loci. Because re- combination in +/t individuals is suppressed within the entire t-complex, which encompasses the MHC,
absolute linkage between the viability and antigen loci was assumed. This recombination suppression has ev- idently served to maintain the strong gametic-level disequilibria that have been observed between the
MHC and the t-complex (HAMMERBERC and KLEIN
1975; LEVINSON and MCDEVITT 1976; STURM, FI-
GUEROA and KLEIN 1982). I obtained the conditions under which a modifier locus evolves to recognize a single MHC locus as an incompatibility locus. Enhanc- ing heterozygosity at the MHC locus reduces the rate of conception of offspring carrying the inviable or sterile t / t genotype if the number of +-specific MHC
antigens exceeds the number of t-specific MHC anti- gens. I found that a modifier locus situated anywhere in the genome would evolve to enhance expression of maternal-fetal incompatibility in response to matching of MHC antigens, provided that the ratio of the number of +-specific to t-specific antigens exceeds twofold, a condition that is satisfied in natural popu- lations.
A three-locus model for the origin of self -1ncom- *
patibility: Whether parental discrimination on the
tween the two characters. In the house mouse system, the strong gametic-level associations between the t-
complex and the MHC appear to have arisen at the origin of the t-complex and have persisted as a result of recombination suppression. An important question is whether the origin of incompatibility systems re- quire unusual genetic associations of historical origin, or whether more general conditions permit the evo- lution of associations of the appropriate kind. In order to address this question, I consider a quantitative model which permits the simultaneous evolution of an overdominant viability locus, an initially neutral antigen locus, and an unlinked modifier locus that causes the expression of gametophytic self-incompat- ibility in response to the specificities determined by the antigen locus. In functional genetic terms, the antigen locus may encode glycoproteins which inhibit fertilization by pollen expressing certain specificities, while the modifier locus influences the level of incom- patibility by controlling the expression of the proto-.3 locus. The structure of the model permits the direct evaluation of genetic associations among the proto-S locus, a locus controlling viability, and a modifier of the expression of incompatibility.
CHARACTERIZATION OF T H E SELF-
COMPATIBLE RESIDENT POPULATION
Structure of the model
Three loci undergo evolution in the model: locus B
affects offspring viability, locus A encodes antigens or specificities, and locus M determines whether the spec- ificities encoded by A are recognized as a switch for incompatibility. Loci A and B may reside on the same chromosome, with recombination between these loci occurring at rate r (0 d r
<
Y2). The modifier locus M shows linkage with neither the viability nor the antigen locus. Two alleles segregate at each of the loci A (Ao and A I ) and B ( B o and B 1 ) . Homozygotes at the viability locus ( B d o and BIBI) survive to reproduce at rate u (0 d u S 1 ) relative to heterozygotes. Initiallylocus M is monomorphic for allele M o , which does not recognize the antigen alleles as switch genes. The resident population is fully self-compatible, with selec- tively neutral variation at locus A. A fraction s (0 s
1 ) of the pollen received on any given stigma comes from the seed parent itself.
Let uy denote the frequency of resident (MdMO)
individuals that carry genotype i at the antigen locus and genotype j at the viability locus (i 0,1,2; j = O , l ) , with genotypes 0, 1 , and
2
corresponding re- spectively to At&, A d l , and AlAl at the A locus, and genotypes 0 and 1 at the B locus to homozygotes (BoBoand BIBI) and heterozygotes ( B a 1 ) . Of the two con-
figurations of doubly heterozygous genotypes, the coupling form (AoBo/AIB1) occurs with frequency ullC
and the repulsion form (AaI/A1B0) with frequency
u11
.
Equilibrium distribution of genotypes
Viability locus: While asymmetric overdominance (under which homozygotes have unequal viabilities) results in the fixation of the more viable homozygote for sufficiently high rates of self-fertilization (HAYMAN
1953; KIMURA and OHTA 197 1 Appendix A4), sym- metric overdominance ensures convergence to an equilibrium state in which the two homozygotes occur in equal frequencies. Let u1 denote the frequency of
heterozygotes at the B locus ( B d , ) , and uo the fre- quency of homozygotes ( B d o and B I B I ) . The popu- lation converges to a polymorphic state characterized
R
by
u1 =
-
1 - s2 m - s (la>
uo = ( 1
-
UI), (lb)in which
w,
the mean viability in the population, corresponds to the larger root of2 m 2
-
Iql+
u(l+
s)]+
su = 0. (2)Joint distribution of the antigen and viability loci:
Appendix 1 records recursions in the ten two-locus genotypic frequencies. The frequencies of the two homozygotes at the B locus converge to equality within each A locus genotype: for example, genotypes Ado/A,,Bo and AoBl/AoB1 occur in equal frequencies at equilibrium (see Appendix 1). These symmetries further imply equality between the frequencies of the two double heterozygotes ( u l = u1 lR).
These simplifications permit a reduction in the number of variables representing genotypic frequen- cies. Because locus A is selectively neutral, one expects no gametic level disequilibrium. This expectation is easily verified by demonstrating that the A locus alleles occur in the same relative frequencies among homo- zygotes and heterozygotes at the B locus:
uoo
+
u10/2 = pouou o 1
+
u11/2 = Paulu20
+
%0/2 = Flu0UP1
+
u11/2 = plul(3)
in which
po
andp l
denote the frequencies of alleles AO and A I( P o
+
p l
= 1). Under (3), the frequencies of heterozygotes at the A locus (AOAI) are determined frommu10 = u[popl(l
-
s)+
s(u10+
Cu11/2)/2] ( 4 4W U l l = pop1(1
-
s)+
SUll(1-
C/2)/2, (4b) in whichModification of Incompatibility
For values of the recombination fraction ( r ) in the range [0, l/z],
C
is a monotone function of r and varies over [0, 13. Solution of (4) yields expressions for the frequencies of individuals that are heterozygous at theA locus:
Using (3) and (6), one can obtain expressions for the remaining genotypic frequencies.
Effects of viability selection in inbred populations
Inbreeding generates associations among loci throughout the genome; consequently, selection im- posed on any particular region transforms the genome as a whole. In agreement with theoretical expectation, inbred plant populations exhibit extensive associations among loci (see summary by HASTINGS 1990). BROWN (1979) has documented the consistency with which excess heterozygosity relative to expectations based on estimates of selfing rates is observed at allozyme loci in inbred populations of plants. This aspect of the “heterozygosity paradox” appears to reflect the re- sponse of the entire genome to selection on any region within it.
Identity disequilibrium: For populations undergo- ing partial selfing, characterize the lineage of any individual by the number of consecutive generations of selfing in its immediate ancestry. T h e expected homozygosity at any neutral locus depends solely on this aspect of the genealogy of the individual. Because every locus within a given individual has the same history, the homozygosities at neutral loci situated anywhere in the genome become correlated. In a more general context, COCKERHAM and WEIR (1968) introduced the term identity disequilibrium to de- scribe departures between the probability of two-locus identity and the product of the probabilities of one- locus identity.
In the model under study here, let x, y and z denote the genotypic frequencies at the A locus (A,&, A d l and AIAI):
x = 2400
+
Uo1y = UIO
+
UI1(7)
z = 2420
+
uz1.T h e genotypic analogue of identity disequilibrium ( q ) is given by:
u11
-
yul = H q , ( 8 4in which y represents the frequency of heterozygotes
Horn Het
“0 “t
FIGURE 1.-Frequencies of genotypes at the antigen locus and the viability locus. Homozygotes at the viability locus occur with frequency uo and heterozygotes with frequency u,. The marginal frequencies of the antigen locus genotypes are x, y and z.
at the A locus
(7),
and H the product of the hetero- zygosities at the two loci,in which q o and q l , the frequencies of alleles Bo and B I , are both equal to I/z. Substitution of (6) into (8) yields an expression for q :
4 ~ ( l
-
S ) U ~ [ U ~ U+
~ l ( l-
C)]= q 4 w
-
4 2
-
C ) ] 9 (9)in which U O and u1 are given in (1). Using (3), (7), and
(8), the two-locus frequencies can be expressed in terms of the genotypic frequencies at each of the loci and (Figure 1). Those expressions indicate that the joint genotypic distribution between a neutral locus
and an overdominant locus has the same form as for two neutral loci (see WEIR and COCKERHAM 1973), with q now dependent on the intensity of viability selection as well as linkage and the rate of selfing [see (9)]. Positive identity disequilibrium implies an excess of double heterozygotes ( ~ ~ 1 ) and double homozy-
gotes ( ~ 0 0 and u z o ) , and a deficiency of single hetero- zygotes ( u ~ o , uo1, u ~ l ) relative to expectations based on the genotypic frequencies at each locus separately.
Effect of viability selection on identity disequi- librium: In the absence of viability selection, identity disequilibrium depends solely on the recombination fraction between the two loci and on the selfing rate:
4 ~ ( l
-
~ ) [ l+
( 1-
C)(1-
s)]TIN =
(2
-
s)2[4-
s(2-
C)] (10) (WEIR and COCKERHAM 1973). Figure 2 indicates that identity disequilibrium between two selectively neu- tral loci (qN) increases with linkage (lower C ) . For comparison, Figure 3 displays identity disequilibrium under overdominant viability selection [ q in (9)] as- suming complete linkage (C = 0) for three values of the selection parameter u. Viability selection can sub- stantially increase identity disequilibrium under high rates of selfing.0.5
-
r = 0.00o,4. .
-"r = 0.25
-"r = 0.50 0.3-
0.2
0.1
0.0 0.2 0.4 0.6 0.8 1 .o Fraction Self Pollen
FIGURE 2.--ldentity disequilibrium between two selectively neu- tral loci. Recombination between the loci occurs at rates r = 0.00, 0.25 and 0.50.
0.5
-
sigma = 0.500.0 0.2 0.4 0.6 0.8 1 .o
Fraction Self Pollen
FIGURE 3,"Identity disequilibrium between a selectively neutral locus and an absolutely linked locus subject to symmetric overdom- inance. Homozygotes at the locus under selection survive at rates u = 0.00, 0.25, and 0.50.
terious mutation in a population undergoing partial selfing. For completely recessive deleterious muta- tions, identity disequilibrium converges to
4s(l
-
s)[l+
(1-
C)(1-
ST)](2
-
s)[2-
s+
2 4 1-
~ ) ] [ 4-
4 2-
C)]'qM = ( 1 1)
in which T represents the relative viability of individ-
uals carrying the deleterious mutation in homozygous form (OHTA and COCKERHAM 1974). Figure 4 plots
q M under absolute linkage (C = 0) for three values of the viability parameter T. In contrast to overdominant selection, selection opposing a deleterious recessive uniformly decreases identity disequilibrium.
Effect of viability selection on homozygosity at the neutral antigen locus: COCKERHAM and RAWL- INGS ( 1 967) showed that inbreeding depression con- fers an apparent overdominance at neutral loci. T h e
0.5
-"tau = 0.0
0.4 -
0.3-
0.2-
0.1 ~
-O- tau = 0.5
tau = 1 .O
-
n n Y
I
Fraction Self Pollen
FIGURE 4,"Identity disequilibrium between a selectively neutral locus and an absolutely linked locus subject to mutation to delete- rious, recessive alleles. Individuals carrying mutations in homozy- gous form survive at rates T = 0 . 0 0 , 0 . 2 5 and 0.50.
expected frequency of heterozygotes at the neutral locus is
(WRIGHT 1921). Under the assumption that offspring derived by selfing survive to reproduce at a rate
4
relative to offspring derived by outcrossing, the fre- quency of heterozygotes converges to
4p0pl(l
-
s)sf$
+
2(1-
s) ( 1 2b)(COCKERHAM and RAWLINCS 1967). Inbreeding de- pression generates an excess of heterozygotes at the neutral locus, as (1 2b) exceeds ( 1 2a) for all 4 less than unity.
This excess heterozygosity derives entirely from the reduction in the net selfing rate due to inbreeding depression. Because inbred offspring are less viable, the fraction of inbred offspring among the survivors of viability selection
(i)
is less than the fraction of inbred offspring at conception (s). T h e net selfing rate, defined as the frequency of inbred offspring at reproduction, is given bys 4
S =s 4
+
( 1-
s)' (1 3)Substitution of
s^
for s in (1 2a) yields (1 2b), indicating that the apparent heterozygote excess derives wholly from the reduction in the net rate of selfing caused by inbreeding depression.Symmetric overdominance also generates an appar- ent heterozygote excess at the neutral antigen locus. Equation 6 indicates that the frequency of heterozy- gotes at the A locus converges to
4popl(l
-
S)(l+
U0[2q-
S(l-
c ) ] ]1.5
"-1- r = 0.00
Homozygote Viability
FIGURE 5.-Ratio of heterozygotes at a neutral locus linked to a locus subject to symmetric overdominance relative to the frequency of heterozygotes expected under complete neutrality for a rate of receipt of self-pollen (s) set equal to 1/2. The heterozygous excess is plotted for rates of recombination between the neutral locus and the selected locus of r = 0.00, 0.25, and 0.50.
Figure
5,
which plots the ratio of (14) to (1 2a) for sset equal to l/2, indicates that tight linkage (small C ) can markedly increase the frequency of heterozygotes at the neutral locus. T h e net selfing rate induced by symmetric overdominance is
~ S [ U l ( l
-
u)+
2a]s =
l + u
Replacement of s in (12a) by
i
in (15) yields an in- creased expected heterozygote fraction. Figure 6 plots the ratio of the actual frequency of heterozygotes at the neutral locus (1 4) to this revised expected value, setting s equal to '/z as before. While the reduction in the net rate of selfing by viability selection accounts for most of the heterozygote excess at neutral loci that are not linked to the viability locus (C = l), excess heterozygosity exists for all levels of linkage, and attains high values under tight linkage (low C).INTRODUCTION OF WEAK GAMETOPHYTIC
INCOMPATIBILITY
Weak gametophytic incompatibility arises upon the introduction of a rare allele (MI) at the modifier locus. Individuals that carry allele M1 weakly inhibit ferti- lization by pollen grains that share an antigen allele. Of the 36 possible three-locus genotypes, 10 (carrying genotype M&fo) are common and the remaining 26 ( M a I or M I M ~ ) are rare. Symmetric overdominance permits reduction of the number of common geno- types to the 6 shown in Figure 1, and the additional assumption of free recombination between the modi- fier locus and both the antigen and viability loci re- duces the number of rare genotypes to 12. Let wlj denote the frequency of M1Ml individuals that carry antigen locus genotype i (i = 0,1,2, corresponding to
v) 1.4 v)
'
1.3a 0
N 1.2
-
r = 0.258
-
r = 0.50c
R
0, $2
I 1.1
a¶
1 .o
0.0 0.2 0.4 0.8 0.6 1 .o
Homozygote Viability
FIGURE 6.-Ratio of heterozygotes at a neutral locus linked to a locus subject to symmetric overdominance relative to the frequency of heterozygotes expected under complete neutrality for a net rate of self-fertilization of j [see (15)]. Recombination occurs between the neutral locus and the selected locus at rates r = 0.00, 0.25 and 0.50.
A d o , A d , , and AIAI) and viability locus genotype j ( j = 0,1, corresponding to homozygotes and hetero- zygotes at locus B ) . Similarly, ug denotes the frequency of M a , individuals with genotype
i
at the antigen locus and genotypej at the viability locus.Linearized recursions in modifier frequencies T h e expression of incompatibility alters the geno- typic distribution among offspring without affecting seed set. All genotypes produce the same number of offspring through seeds, with the relative genotypic frequencies among those seeds determined by the frequencies of compatible and incompatible pollen received through selfing and random outcrossing, and the relative fertilization success of incompatible pol- len. For example, an individual with genotype M1Aof MIAo reduces the rate of fertilization by pollen carry- ing allele A. to g relative to A I pollen. T h e average fertilization rate among pollen received by this geno- type from self (fraction s) and through random out- crossing (fraction 1 - s) is
No = sg
+
(1-
s)(POg+
PI),
( 1 63)in which P o and
pl
denote the frequencies of alleles Ao and A l in the population. Similarly, the average ferti- lization rate among pollen received by MIAl/MIAI individuals isN1 = sg
+ (1
+
s)(Po+
P1g). (16b) Individuals with genotype MIAo/MIAl recognize and equally inhibit both antigen alleles, generating no net distortion in genotypic frequencies among their off- spring. In heterozygous form ( M a , ) , the active mod- ifier allele reduces the rate of fertilization by incom- patible pollen toin which E (0 d E
<
1) denotes the degree of domi- nance ofM I
in the stylar response. T h e average fer- tilization rates inM a l
individuals that are homozy- gous at the antigen locus are similar to No and N1 in (1 6), with g replaced by (1 7).For analytical tractability, I further assume com- plete dominance of the active modifier allele MI ( E = 1). Analyses of models in which a single locus was assumed to encode both antigen expression and stylar response indicate that the conditions for the origin of both gametophytic (UYENOYAMA 1988b, c) and spo- rophytic (UYENOYAMA 1989c) self-incompatibility are most stringent under full dominance in stylar re- sponse. I expect that the initial increase conditions obtained for the model presently under consideration are conservative in the sense that reductions in dom- inance may permit the origin of incompatibility under less stringent conditions.
The assumption of complete dominance of stylar response permits a change of variables that reduces the number of variables in the initial increase analysis from 12 to 6. Let zy (i = 0,1,2; j = 0 , l ) denote the frequency of allele
MI
among individuals that carry antigen genotype i and viability genotype j :zy = wy
+
4 2 . (1 8)Up to terms linear in the frequencies of the rare genotypes, recursions in those frequencies are given by
w Z 6 0 U ( s [ ( z o o
+
Zo1/2)4g/No+
IO+
Z I I ( ~-
c/2)]+ po(
1-
S)[(ZOO+
% O l ) ( l+
g/No)+
z10+
z11])/4Wz;,
= (s(zOl2g/No+
ZllC/2)+
pO(1-
s)w z i o = u(s(z1o
+
z1,/2)C/2)+
(1-
s)-
[(zoo+
Z O l ) ( l+
g/No)+
z10+
z11])/4* [(zoo
+
zol)pl(l+
l/NO)+
z1o+
Z l l+
(z20+
~ ~ ~ ) p ~ ( 1+
w 1 ) ~ ) (19)W Z i I = (sz112(1
-
C/2)+
(1-
s)' [(zoo
+
zol)pl(l+
1/No)+
z10+
Z l l+
(zzo+
z21)po(l+
l/Nl)]V4W Z ~ O
= ~ ( s [ ( Z n o+
Z21/2)4g/N1+
% I O+
z11(1-
C/2)]+
p,(l
-
s)[(z2o+
z21)(1+
g/N1)IO
+
z11])/4-
W%61 = (s(zn12g/N1
+
ZllC/2)+
pl(1-
s)* [(QO
+
z21)(1+
g/Nd+
%IO+
zlllv4.Initial increase condition
Stability criterion: Represent the transformation in (19) by
z' =
Mz,
(20)in which z = (ZOO, z01, %IO, z l l , z20, zzl) denotes the
vector of genotypic frequencies and
M
a six-dimen- sional matrix. BecauseM
is non-negative, a necessary and sufficient condition for local stability is that the determinants of all principal minors of the matrix (I-
M) be positive:Deti[I
-
MI>
0, (21)in which Deti denotes the determinant of an i-dimen- sional principal minor [see GANTMACHER (1959) p. 711. Further, the irreducibility of
M
under partial selfing (0<
s<
1) ensures that its leading eigenvalue is real and non-negative and corresponds to a simple root of the characteristic equation [Frobenius Theo- rem, see GANTMACHER (1959) Chapter XIII]. If MIwere to generate no incompatibility response ( g = l), the determinant of (I
-
M)
would be zero and the lower order principal minors (21) positive. Under sufficiently weak inhibition generated by the active modifier allele MI (g sufficiently close to unity), the lower order principal minors remain positive, with the negativity of the characteristic equation evaluated at unity necessary and sufficient for the invasion ofM I :
Det[I
-
MI<
0. (22)T h e remaining analysis is restricted to modifiers with sufficiently weak effects.
Using the equilibrium Equations 1 and 2, Equation 22 expressed in terms of the genotypic frequencies in the resident population corresponds to
(1
-
g)s(l-
s)(-U(4popl-
U I O-
U11)(2W-
s)+
2W(l-
a)[p,(Uoo-
UOI)+
p0(U20-
uel)ll>
0. (23)Under the assumptions of partial selfing (0
<
s<
1) and active modifier alleles ( g<
l), the expression in curly brackets in (23) determines local stability. Using the genotypic frequency distribution given in Figure 1 (together with (l), (3), and (9)) and using thatw
is the larger root of (2), (23) can be expressed solely in terms of the basic parameters of the system.Complete linkage: In the absence of recombination between the antigen and the viability loci (C = r = O ) ,
the sign of (23) depends on a quadratic in the fraction of self-pollen received (s):
s23a(2
-
a)-
~ ( l-
2 ~ ) ( 1-
3 ~ )For intensities of viability selection in the range
’/4
<
Qc
2/3, (25) the modifier allele that generates the expression of weak gametophytic incompatibility increases when rare for rates of receipt of self-pollen (s) greater than the single root of (24) lying in the interval (0, 1).Incomplete linkage: In the presence of recombi-
nation (C
>
0 ) , satisfaction of (23) depends on a cubic in s [see (A2.1) in APPENDIX21.
Analysis of this cubic indicates that modifier alleles causing the expression of incompatibility can increase when rare for inter- mediate intensities of viability selection (a).For loose linkage, satisfying
C > 4a2(2
-
3a)(1
-
u)‘+
2a2’invasion of the active modifier allele requires
‘/3
<
Q<
2/3. (2’7) Under (26) and (27), a single root (s*) of the cubic ins lies in the interval (sl, I), for s1 a root of a quadratic (see (A2.2) in APPENDIX 2). The modifier causing incompatibility invades for values of s greater than s*.
For tighter linkage (C violating (26)), the invasion of incompatibility again occurs under (27) for suffi- ciently high rates of receipt of self-pollen (s
>
s*). In this case, s* corresponds to the root of the cubic that lies in the interval (sl, sp), for s2 a root of a quadratic[see (A2.3) in APPENDIX 21. For tight linkage, satisfying
c
c
‘/9 (28)(which corresponds to recombination fractions below approximately 0.059 [see (5)], two roots of the cubic,
s* and s**, come into existence in the range ( $ 2 , 1)
for values of the viability parameter lying in
‘/4
<
a<
‘/3, (29) with Q sufficiently close to ‘/3. The modifier that induces the expression of incompatibility increases when rare for selfing rates lying between s* and s**.Figure
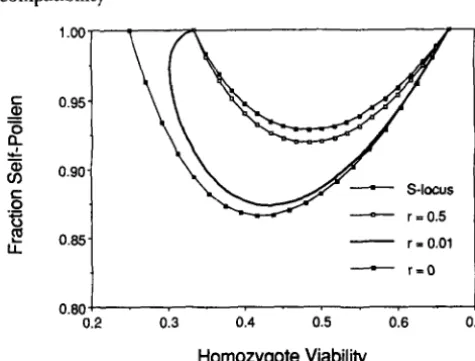
7
displays the threshold rates of receipt of self-pollen that favors the origin of gametophytic self- incompatibility. Under absolute linkage (C = 0) or moderate linkage (C>
2/9), incompatibility is favoredfor rates of receipt of self-pollen (s) exceeding a single threshold. For tight linkage (0
<
C
<
2/9), two thresh-olds arise for Q sufficiently close to %, with incompat- ibility favored for self-pollen fractions lying between the thresholds.
For comparison, J have plotted the threshold self- pollen fraction above which an active S allele encoding weak sporophytic self-incompatibility enters a self- compatible population (from UYENOYAMA 1989~). T h e value displayed (labeled “ S locus”) corresponds
0.95-
0.90-
o.=a
r = 0.010.80
0.2 0.3 0.4 0.5 0.6 0.7
Homozygote Viability
FIGURE 7,”Threshold rates of receipt of self-pollen permitting invasion of the modifier that induces self-incompatibility for rates of recombination between the viability locus and the antigen locus
of r = 0.5, 0.01 and 0. The curve labeled “ S locus” corresponds to results obtained from the model analyzed by UYENOYAMA (1 989c), which assumes that sporophytic self-incompatibility arises with the introduction of a dominant allele encoding both pollen specificity and the stylar rejection response. The condition obtained under absolute linkage between a locus under symmetric overdominance and the S locus is plotted.
to a dominant allele encoding both antigen expression and the stylar rejection response introduced at a locus that shows absolute linkage to a biallelic viability locus subject to symmetric overdominance. Looser linkage requires even more stringent conditions (higher self- ing rates) for the invasion of the active S allele. Figure
7
indicates that even under absolute linkage (the link- age relationship most conducive to the invasion of the active S allele), the condition for initial increase is still more stringent than the most stringent condition ob- tained for the three-locus model presented here.COMPONENTS OF SELECTION
of systems of transmission and inbreeding (UYENO- YAMA and BENGTSSON 1989; UYENOYAMA and
WALLER 1991a, b, c), and of sexual selection (OTTO 199 1).
Change of basis
Definition of variables: Define a new set of vari- ables, representing the frequency of the introduced modifier allele ( M I ) and measures of association in- volving the modifier locus:
z = zoo
+
201+
z10+
z l l+
z20+
z21D
= z01+
2 1 1+
z21-
z(uo1+
u11+
UP])60 = zoo
+
zo1-
z(uo0+
uo1)(30)
6 2 = 220
+
z21-
%(UP0+
u21) 601 = zo1-
zu016 2 1 = z21
-
z u p 1 .Variable z corresponds to the frequency of the intro- duced modifier allele M1, with the remaining variables representing associations between Ml and alleles or genotypes at the antigen or viability loci:
D = f(M1 with B O P ] )
-
f(M1)f(Bo/BJ60 = f(M1 with AoIAo)
-
f(M,)f(A,/Ao)62 = f ( M ] with &/AI) -f(Ml)f(A1/A1)
601 =f(MI with AoBo/AoB1) (31)
-f(Ml)f(AoBo/AoB~)
6 2 1 = f ( M 1 with A I B o / A ~ B 1 )
-f(Ml)f(AIBo/AIBl),
in which
f
denotes the frequency of the designated alleles or genotypes. D represents an association be- tween MI and the favored heterozygous genotype at the viability locus, 6o and associations between MIand homozygotes at the antigen locus, and 601 and 621
associations between MI and genotypes that are homo- zygous at the antigen locus and heterozygous at the viability locus. Positive D implies a positive association between the enhancer of outcrossing (MI) and the genotype with the highest viability (BoB1).
Change in gene frequency: Using the new basis described in (30), the change in frequency of the introduced modifier allele ( A t ) can be expressed in terms of the measures of association among loci. This expression for the change in frequency of M1 over a single generation is directly related to the condition for the asymptotic increase of MI (see also UYENO-
YAMA and BENGTSSON 1989; UYENOYAMA and
WALLER 1991 b). Let N represent the stability matrix associated with the new basis. This matrix is related
to the matrix associated with the original basis
[M
in (20)l byN
=AMB,
(32)in which A translates from the old basis to the new basis [from (30)], and B translates from the new basis back to the old basis (B = A-'). Because any change of basis preserves eigenvalues and eigenvectors, the positivity of the determinant of (I
-
N)
[see (22)] ensures the invasion of allele M I .Using the basis defined in (30), the condition for the invasion of MI (23) is
AZ
cc Ds(1-
~ ) 2+
(1-
g)s( 1-
s)z[(~P~P~
-
y)( W O
-
2Ws)+
(1-
a)pop,v]>
0 , (33)in which Wo and Ws represent the average viabilities of offspring derived by random outcrossing and by selfing:
w,
= (1+
u)/2 (344w,
= uou+
Ul(1+
u)/2, (34b)and q represents the equilibrium association in hetero- zygosity between the antigen and viability loci (9).
Condition (33) indicates that the twofold inbreeding depression condition ( Wo
>
2Ws) would apply only in the absence of associations between the viability locus and both the antigen and the modifier loci. Positive associations ( q , D>
0) permit the invasion of enhancers of incompatibility under levels of inbreeding depres- sion that are less than twofold.Analysis of associations
One-generational and asymptotic change: While the expression for the change in frequency of the introduced modifier allele (33) provides an appealing representation of the role of genetic associations (D and q ) in promoting incompatibility, it describes ge- netic change over only a single generation. In con- trast, local stability analysis (22) resolves the asymp- totic behavior of the system initiated at any arbitrary state in a sufficiently small neighborhood of the initial equilibrium state. In order to integrate these two descriptions of evolutionary change, I describe a gen- otypic distribution which has the property that the change in gene frequency over a single generation in a population initiated at this particular distribution reflects the asymptotic behavior of the system initiated at any arbitrary position close to the equilibrium. This property requires that the change in gene frequency in a population initiated at this particular genotypic distribution be proportional to the characteristic equa- tion evaluated at unity:
Describe the population in the current generation by:
v = (z, D, 80, 82, 801, 6 2 1 ) ~ " ~ (36) in which tr denotes the transpose. For any v, (N
-
I)v represents the vector of changes in the variables over one generation:(N
-
I)v = (h, AD, A&, A&, A601, Ad21)~'. (37)Define
i
such that all elements of (N-
1)i but the first are zero. The first element of (N-
1 ) i is equal to the change in the frequency of MI(Az) over one generation in a population initiated ati
(37).Let
P
represent the matrix obtained by replacing the first column of an n-dimensional identity matrix by f, for n the dimension ofM
(n equals 6 in this case). Multiplication ofP
on the left by (I-
N) produces a matrix of the form(I
-
N)P =in which R1 is an (n
-
1)-dimensional column vector of zeros,Rn
is an ( n-
1)-dimensional row vector with elements equal to the corresponding elements of (I-
N),
and (I-
N),,-J is the matrix obtained by removing the first row and column from (I-
N). Taking the determinant of both sides of (38) producesDet[I
-
N]Det[P] = Det[I-
N]z(39)
= -AzDet[(I
-
N)n-l][I am indebted to SARAH PERIN OTTO for this method of obtaining (39)]. The desired result (35) follows from (39) provided that
Det[(I
-
N)n-l]>
0 . (40) Appendix 3 presents the argument that (40) holds under any choice of basis, such as (30), for which all variables other than z represent measures of associa- tion that reduce to zero for the zg proportional to the uy.Relative effects of associations: At the distribution
i ,
for which the condition (33) for an increase in thefrequency of the introduced modifier allele over a single generation corresponds to the asymptotic con- dition (23) for invasion, the association between the modifier and viability loci is
D =
r(1-
g)s(l-
s)V4[2@
-
S(UO+
UU~)]'(4
1 a) in which V i s given byV = (IlPopl
-
y)[u+
(1-
G)UO(UO-
ul)]+
27lPop1(uo+
nu,)>
0. (4 1 b) Expression (4 1) indicates that the enhancers of incom- patibility (1>
g) develop positive associations with the favored heterozygous genotype at the viability locus.-
Residual Cost-
DMB-
DAB2.0
1.5
1 .o
0.5
0.0
0.0 0.2 0.4 0.6 0.8 1 .o
Fraction Self Pollen
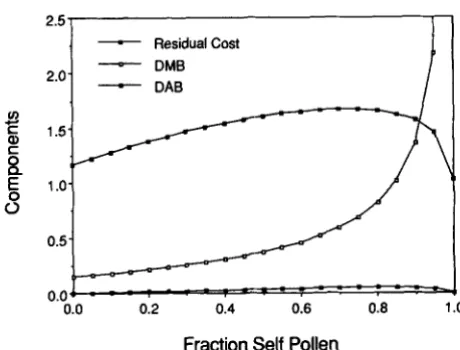
FIGURE 8.-Relative magnitudes of terms contributing to the change in gene frequency at the modifier locus (33) which involve
11 ( D A B ) , D (DMB), and the remainder (residual cost of outcrossing) for the case Y = u = 0.5. Modifier alleles that confer weak incom-
patibility increase when rare only if the sum of DAB and DMB exceeds the residual cost.
Positive identity disequilibrium between the antigen and viability loci ( 0 ) promotes the origin of incompat- ibility both through direct effects on the frequency of the modifier [see (33)], and by strengthening the association between the enhancer of incompatibility and high viability [see (41)].
Because the average viability of outbred offspring never exceeds twice that of inbred offspring (WO C
2Ws), incompatibility evolves only if the genetic asso- ciations
(D
and 7) compensate for the residual cost of outcrossing. Represent the effect of associations be- tween the viability and antigen loci (DAB) by the term involving q, and the effect of associations between the viability and modifier loci (DMB) by the term involv-ing D , for 9 given by (9) and D by (41). The negative
of the remaining term corresponds to the residual cost of outcrossing. Figure 8 compares the magnitudes of the three components of selection for the case T- = u
= 0.5. Incompatibility evolves only if the sum of the curves labeled DAB and DMB exceeds the residual cost curve. Figure 8 indicates that it is primarily the asso- ciation between the modifier and the viability locus (DMB) that compensates for the residual cost of out- crossing, with identity disequilibrium between the an- tigen and viability loci (DAB) having relatively little effect. For high rates of selfing, the modifier allele that confers incompatibility becomes strongly associ- ated with the heterozygotes at locus B , which implies that its carriers have higher expected viability. While linkage between the antigen and viability loci ( r
c
0.5) increases the effect of identity disequilibrium, this effect remains small relative to the residual cost of outcrossing (see Figure 2).DISCUSSION
determining in large part the nature of interactions among its components. While the origins of self-in- compatibility are obscured by its antiquity, recent studies of the genetic basis of S locus systems provide clues as to the structural framework within which they evolved.
Origin of the
S
locusIn Brassica, the S locus appears to be a member of a multigene family, with sequences showing some
homology present at perhaps eleven locations (NAS- RALLAH,
Yu
and NASRALLAH 1988). LALONDE et al.(1989) isolated an S locus-related sequence, designat- ing it SLRl
.
Transcripts of SLRl are abundantly produced in parallel with those from the S locus, appearing in stigmatic tissue at the same of develop- mental stages. Unlike the gene that encodes the Slocus glycoprotein, the SLRl gene is conserved among strains carrying different S alleles.
Sequences homologous to the SLRl gene have been detected in both self-compatible and self-incompatible species of Brassica, and in other crucifers, including
Arabidopsis thaliana (SC) and Raphanus sativus (SI)
(NASRALLAH, Yu and NASRALLAH 1988; LALONDE et al. 1989). LALONDE et al. (1989) suggested that the SLRl gene may perform an important function dis- tinct from self-incompatibility. These findings are con- sistent with the view that the S-locus may have been derived from an existing gene rather than arising de
novo, with the expression of self-incompatibility aris- ing through modification of a different function.
A second incompatibility locus (G), controlling the gametophytic determination of pollen specificities, has been reported in two crucifers, Brassica campestris and
R. sativus, the primary self-incompatibility system of
which is sporophytic (LEWIS, VERMA and ZUBERI 1988; ZUBERI and LEWIS 1988). In both species, the
S and G loci interact, with matching at the G locus causing incompatibility in the presence of certain S
alleles. The two loci appear to show partial linkage. LEWIS, VERMA and ZUBERI (1 988) suggested that the
G locus represents a relic of an ancestral self-incom- patibility system.
The finding of an additional locus involved with self-incompatibility and of a family of related genes in the crucifers lends credence to BATEMAN’S (1952) hypothesis that the origins of the mechanism of self- incompatibility and the regulation of its expression may represent separate evolutionary events. Regula- tion of self-incompatibility in primitive systems may have originated with the recognition of one or more existing loci as “switch genes.”
Self-incompatibility evolves under less than twofold levels of inbreeding depression
Predictions of conventional models: In conven- tional quantitative models, the origin of self-incom-
0.5
1
r
0
’
4
i
1
-
-
E = 0.10 E=0.33-
E11.000.04 . ‘ .
Y
0.0 0.2 0.4 0.6 0.8 1 .o
Fraction Self Pollen
FIGURE 9.-Maximal viabilities of offspring derived by selfing relative to that of outbred offspring that permit invasion of an active S allele causing gametophytic self-incompatibility for rates of expression of stylar rejection of E = 0.01, 0.10, 0.33 and 1.00
(UYENOYAMA 1988~). Higher rates of expression of the active S
allele discourage the origin of self-incompatibility.
patibility entails the invasion of a fully functional S allele encoding all aspects of incompatibility, including the determination and expression of pollen specifi- cities and their recognition and rejection by com- ponents in the stigma or style (see, for example, CHARLESWORTH and CHARLESWORTH 1979; UYE- NOYAMA 198813, c). In these models, the viability of offspring is assumed to depend only on whether they were derived by selfing or by outcrossing. An
S
allele encoding gametophytic self-incompatibility invades a fully self-compatible population only ifs4(24
-
1)[1-
E(1-
g)]+
E [ 4 ( 3-
2s)(421 \ I
-
(1-
s)][1-
s(l-
@(1-
g)]<
0(from Equation 14a in UYENOYAMA 1988c), in which
4 denotes the viability of inbred offspring relative to
outbred offspring, g the rate of fertilization by incom- patible pollen relative to compatible pollen in styles that are homozygous for the active S allele, E a meas- ure of dominance of the expression of the active S allele in the style [see (17)],
and s the fraction of pollen each plant receives that is derived from itself. Under weak inhibition of incompatible pollen [( 1-
g)
in (42) considered negligible], the active S allele increases when rare only if the relative viability of inbred off- spring (4) lies below the values plotted in Figure 9. Figure 9 indicates that lower levels of expression of self-incompatibility by individuals carrying only one active S allele (small E ) promote the invasion of the Sallele. Under higher rates of expression of stylar re- jection by heterozygotes ( E 3 %), invasion of the active S allele requires that the viability of outbred offspring exceed that of inbred offspring by more than threefold (4
<
Y3).structure from conventional models, the quantitative model presented here explicitly incorporates genetic loci which control different components of the system. The proto-S locus is represented as the antigen locus, which initially expresses no incompatibility activity. A separate modifier locus influences the expression of incompatibility by the proto-S locus, perhaps by reg- ulating the level at which it produces an inhibitory factor. Inbreeding depression derives from segrega- tion of alleles at a locus subject to overdominant viability selection, rather than appearing as a constant parameter (4) of the model.
While conventional models require very high levels of inbreeding depression (see Figure 9), inbreeding depression in a population subject to symmetric overdominance in viability never approaches twofold
(4
b 9'3, ZIEHE and ROBERDS 1989). In contrast, thethree-locus model analyzed here indicates that for intermediate intensities of viability selection spanning one-third of the parameter range (Y3
<
<
*/3), there exists some rate of receipt of self-pollen that is suffi- ciently high to permit the origin of self-incompatibility (see Figure7).
Evidently, genetic associations among components of the incompatibility system that evolve immediately upon the introduction of the enhancer of self-incompatibility promote its increase.A growing body of work indicates that the coevo- lution of modifiers of the breeding system and deter- minants of viability generate significant selective pres- sures (CAMPBELL 1986; HOLSINCER 1988). Analytical results describing the nature, magnitude, and effects of genetic associations between a locus controlling viability and a modifier of meiotic reproduction (UYENOYAMA and BENGTSSON 1989) and the rate of self-fertilization (UYENOYAMA and WALLER 199 la, b, c) indicate that such associations give rise to the major differences observed between models that permit co- evolution and those that do not. Assuming that asso- ciations between genetic determinants of viability and modifiers of the breeding system are absent leads to the conclusion that the origin of sporophytic self- incompatibility requires levels of inbreeding depres- sion of at least twofold (CHARLESWORTH 1988). In fact, associations between an overdominant viability locus and the S locus that arise immediately upon the appearance of an active S allele can permit the origin of sporophytic self-incompatibility under substantially lower levels of inbreeding depression (UYENOYAMA
1989~). The assumption that associations among com- ponents of breeding systems have negligible effects fails to hold even immediately following the introduc- tion of rare modifiers. While conventional models make an important contribution in exploring the framework within which the genetic associations among interacting components evolve, predictions de- rived from such constructions often show significant
qualitative and quantitative departures from those obtained from more derailed analyses.
Genetic associations direct the evolution of self-incompatibility
Symmetric overdominance in viability: I analyzed the effects of genetic associations among components of incompatibility systems on the evolution of a rare modifier that recognizes a neutral antigen iocus as a switch for self-incompatibility. The condition which permits the origin of gametophytic self-incompatibil- ity can be represented as a single inequality:
Ds(1
-
0)2 (1-
g)S(l-
S ) Z [ ( 4 P o f i l-
J')(Wo-
2ws)
+
(1-
a)popl~]>
0[(33) reproduced here for convenience], in which z
denotes the frequency of the modifier allele that causes the expression of self-incompatibility, and D and q denote measures of association among loci. This condition indicates that in the absence of genetic associations among the components of the system ( D ,
q = 0 ) , the origin of self-incompatibility requires at
least twofold levels of inbreeding depression (WSIWO
=
4
<
%), in agreement with predictions from simpler analyses (KIMURA 1959). In contrast, the analysis of the full three-locus model presented here indicates that a single overdominant viability locus situated anywhere in the genome can support the invasion of a modifier that causes weak expression of incompati- bility under the specified conditions (see Figure7).
Enhancers of self-incompatibility develop direct as- sociations with heterozygotes at the overdominant viability locus [D
>
0, see (41a)l. In addition, positive associations in heterozygosity between the proto-S locus and the viability locus [q, see (9)] ensure that the increase in heterozygosity at the S locus among off- spring induced by the expression of incompatibility implies an increase in heterozygosity at the overdom- inant viability locus. Both kinds of associations pro- mote the origin of self-incompatibility by improving the viability of carriers of enhancers of self-incompat- ibility.General forms of viability selection: While the genetic model studied here incorporates a particular form of viability selection, the analysis of genetic associations and the manner in which they influence the fate of the rare enhancer of incompatibility pro- vides a framework for conjecture concerning the ev- olution of self-incompatibility under general selective regimes. Associative overdominance refers to the ap- parent overdominance conferred on a selectively neu- tral locus by selection elsewhere in the genome. As- sociative overdominance at the proto-.S locus (through