CORRELATION OF PHENOTYPIC AND GENOTYPIC ADHESION
AND BIOFILM FORMATION FEATURES IN
CANDIDA ALBICANS
STRAINS ISOLATED FROM DIFFERENT CLINICAL SPECIMENS
1
*,2Omar Sadik, 1,2Othman Almahdawy, 1,2Dunya A. A. Gurgea, 1,2Lia-Mara Ditu,
1,2
Alina Maria Holban, 1,2Gratiela Gradisteanu, I. Gheorghe, Otilia Banu3,
1,2
Mariana Carmen Chifiriuc
1
University of Bucharest, Faculty of Biology, 2Research Institute of the University of Bucharest, 3Institute of Cardiovascular Diseases Prof. C.C. Iliescu.
ABSTRACT
In the recent years, there have been important changes in the epidemiology of fungal infections, Candida albicans having emerged
as an important cause of invasive infections mainly among immunocompromised patients. Considering the increasing prevalence rate of candidiasis worldwide, the aim of the present study was to investigate the possible correlations between the adherence and biofilm development capacity and the adherence genes profiles of 109 C. albicans strains recently isolated from different clinical specimens.
The highest adherence capacity was registered for strains isolated from urine and sputum samples, while the most intensive biofilm producers were those isolated from respiratory tract secretions. The most frequent genes encoding for C. albicans adhesins in the studied strains were ALS1 and ALS3 genes, demonstrating their important role in the pathogenicity of the respective clinical isolates. We have found a specific correlation between the ability to colonize the cellular substratum and the presence of ALS1 gene in case of urine isolates, of ALS3 in sputum and tracheal isolates, and of SAP 2 in bronchial and tracheal secretions. Also specific correlations between the biofilm development capacity and the presence of ALS1 in tracheal and of ALS8 in urinary isolates have been revealed. These results demonstrate that the detection of these genes in different clinical isolates could represent and easy and cost-effective tool for the prediction of highly virulent isolated
Volume 6, Issue 7, 1753-1770. Research Article ISSN 2277–7105
Article Received on 19 May 2017,
Revised on 09 June 2017, Accepted on 30 June 2017
DOI: 10.20959/wjpr20177-8918
*Corresponding Author'
Dr. Omar Sadik
University of Bucharest,
capable of colonizing either the cellular or the inert substratum and to generate clinical infections.
KEYWORDS: Candida albicans; Biofilm; adherence genes.
INTRODUCTION
Although Candida species are microorganisms that live commensally in the oral cavity of healthy individuals, their incidence in different pathological conditions associated with the expression of different virulence features seem to be increased in immunocompromised patients.[1, 2] These species can become pathogenic in response to physiological changes in the host, causing oral candidiasis or invasive systemic infections.[3] Emerging from the dental plaque reservoir, Candida spp. can also cause endodontic infections, peri-implantitis lesions and periodontitis, especially in HIV-positive and diabetic patients.[4–9] The higher prevalence of Candida spp. in diabetic patients has been associated with the type and duration of the disease and the degree of glycemic control.[10, 11]
Candida spp. may produce some virulence factors that facilitate their proliferation and,
consequently, adherence to the epithelium and invasion of the connective tissue.
Adherence to host tissue is the first step in the pathogenic process, allowing the microorganism to attain a foothold and to colonize a specific niche environment, from which, under suitable predisposing conditions when the host is immunocompromised, take place candidal proliferation, invasion, and, in some instances, dissemination. The mechanisms of adherence to different cellular types or surfaces by Candida ssp. are complex, multifactorial and still not completely understood. It would appear that this process is achieved by a combination of specific (candidal adhesins and receptors or via the production of proteinases) and non-specific (electrostatic forces, aggregation and cell surface hydrophobicity [CSH]) mechanisms.[17] These yeasts produce essential exoenzymes that improve their capacity to establish colonization and/or infections.[12,13] The exoenzymes, such as secreted aspartyl proteinase (SAPs) and phospholipases, degrade immunoglobulins and extracellular matrix proteins, inhibit phagocytosis and induce inflammatory reactions.[6, 14] One of the first early studies to link proteinase production to adherence in C. albicans showed that strongly proteolytic strains of C. albicans adhered significantly more strongly to human buccal epithelial cells in vitro than did strains producing less proteinase.[24] A more recent report
of mice; the higher-Sap-producing strains showed greater levels of tissue colonization in the liver, kidneys, and spleen.[25]
However, the majority of studies linking Sap production with C. albicans adherence have been performed using the proteinase inhibitor pepstatin, which inhibits Sap2 (and probably Sap1 and Sap3) very efficiently.[26] Borg and Ru¨chel[27] demonstrated a marked decrease in C. albicans adhesion and invasion of human mucosa by pepstatin, and a similar reduction of
C. albicans adherence using pepstatin was also shown with human epidermal cells.[28, 29] In anaerobic environments, such as that of the root canal systems and periodontal pockets, the SAP secretion is increased, suggesting that the oxygen concentration in the atmosphere surrounding these cells can exert an influence on the virulence attributes of C. albicans.[22]
The gene EAP1 was isolated as a putative cell wall adhesin. Sequence analysis of EAP1 shows that it contains a signal peptide, a glycosylphosphatidylinositol anchor site, and possesses homology to many other yeast genes encoding cell wall proteins. In addition to increasing adhesion to polystyrene.[35] CSH has a central role in the pathogenesis of C. albicans. Hydrophobic cells exhibit greater adherence to epithelial and endothelial cells and
extracellular matrix proteins and seem to be more resistant to phagocytosis.[18] C. albicans is unique among Candida spp., in that the CSH status varies in response to different environmental conditions and growth phases.[19] Considering the increasing prevalence rate of candidiasis worldwide, the aim of the present study was to investigate the possible correlations between the adherence and biofilm development capacity and the adherence
genes profiles of 109 C. albicans strains recently isolated from different clinical specimens.
MATERIAL AND METHODS Patients samples
A number of 109 C. albicans strains were isolated from patients admitted for surgery in the Institute of Cardiovascular Diseases Prof. C.C. Iliescu, Bucharest, Romania, aged from 20–85 years, from different clinical specimens, i.e.: upper respiratory tract (2 thrush, 10 bronchial,
required inoculum suspension. The YBC cards were afterwards incubated at 37°C for 24 h and submitted for reading. Some of the YBC cards were incubated for an additional period of 24 h, giving a total incubation period of 48 h. The attachment to the inert substratum and the subsequent biofilm production was assessed by using a micro-titer method. The C. albicans strains were grown overnight in Sabouraud broth solution at 370C and centrifuged at 150 rpm. The cells were harvested, washed with phosphate buffered saline (PBS) and standardized to 1 × 107 yeast cells/ml. Afterwards 100 µl from the yeast cell suspension were placed in the wells of a 96-well micro plate and incubated for 96 h at 370C. After incubation
the presence of the biofilm in each well was quantified by two distinct methods: a semi-quantitative method based on the microscopic examination of the biofilm with an inverted
microscope and violet crystal (CV) staining, and by reading the optic density of the biofilm cells suspension at a 490 nm with an ELISA reader. The experiments were performed in triplicate.
For the adherence to the cellular substratum represented by HeLa cells, the Cravioto adapted
method was used. In this purpose the yeast inoculum was prepared from the Sabouraud-glucose broth cultures supplemented with 3% Sabouraud-glucose, after 48h of incubation, at 37ºC. The cultures were centrifuged at 1500 rpm for 10 min and the sediment washed once in PBS, pH 7.2. The density of cells in the final suspension was 108 CFU/ml (corresponding to McFarland 1 standard density). Adhesion to cellular substratum was investigated using 24 h HeLa cells monolayers of 80% confluency grown in 96 multi-well plastic plates, in DMEM medium supplemented with antibiotics. The growth medium was removed by washing the wells three times with PBS. A 1 ml aliquot of the final fungal suspension was aseptically added to each well containing the cellular monolayer. The plates were incubated at 37°C for 2 h, allowing the yeast cells to adhere to the cellular surface. Then, the cells were washed three times with PBS and fixed with methanol for 5 min. Afterwards the wells were stained with Giemsa solution (1:10) (Merck, Darmstadt, Germany) for 20 min, washed with tap water and dried at room temperature, examined microscopically (magnification, ×2500) with I.O. and photographed with a Contax camera adapted for Zeiss microscope.
DNA extraction
agitation at 120 rev/min. After incubation, cells were harvested by centrifugation at 3000 g for 10 min at 4 C and washed twice in 10 ml of PBS (pH 7). The cellular density of each strain was adjusted to 1x107 cells/ml. Pooled biofilms from each site were transferred to separate Eppendorf microtubes containing 1 ml of reduced transport fluid (RTF). Immediately after collection, the samples from each site were diluted and plated onto a Sabouraud dextrose agar (SDA) with chloramphenicol and, Biomerieux, Paris, France) and incubated at 37°C for 48 h in a reduced oxygen atmosphere (10% CO2 and 90% air). DNA
was extracted according by Nascimento et al.[30], with slight modifications. Colonies of
yeasts with growth of 24 h/37 °C onto Sabouraud dextrose agar plates were suspended in 1 mL of 1 M sorbitol and 125 mM of EDTA; suspension was centrifuged (10 min, 13000 x g,
25 °C), the supernatant was discarded, and the pellet was resuspended in 0.5 mL of lysing solution (1 M TrisHCl, pH 8.0, with 250 mM of EDTA and 5% SDS) plus 10 L of proteinase K (Invitrogen) and incubated for 1 h at 65 °C. Next, 500 L 5 M potassium acetate was added, incubated on ice for 2 h and then centrifuged (10 min, 13000 x g, 25 °C). Supernatant was transferred to an Eppendorf tube containing 1 mL of absolute ethanol and mixed by inversion and centrifuged (10 min, 13000 x g, 4 °C). The supernatant was discarded; the pellet was washed with 500 L of cooled 70% ethanol and centrifuged (10 min, 13000 x g, 4 °C). Finally, supernatant was discarded and the pellet was resuspended in 0.5 mL of sterilized MilliQ water. The obtained DNA was quantified in a spectrophotometer at 260 nm (Genesys 10UV, Rochester, NY, USA) to obtain a standard concentration of 100 ng/ml and stored at −20°C for subsequent PCR reactions.
PCR (polymerase chain reaction) for detection of adhesion genes
Statistical analysis
The following statistical data analysis approaches were used in order to analyze and assess the results of the study under application of the statistical package (SPSS) ver. (16.0).
1. Descriptive data analysis
a- Contingency Coefficients for the association tables. b- Graphical presentation by using
- Cluster Bar Charts.
2. Inferential data analysis
These were used to accept or reject the statistical hypotheses, which included the following : a- Contingency Coefficients (C.C.) test for the cause's correlation ship of the association
tables.
Where is the Chi Square statistic and is the overall total of the contingency table.
b- Redistribution (Under/Upper) cutoff point(s) for creating an association tables of 2X2
rank throughout applying Stem-Leaf plot technique.
For the abbreviations of the comparison significant (C.S.), we used the followings. - NS : Non significant at P>0.05
- S : Significant at P<0.05
- HS : Highly significant at P<0.01
RESULTS
The 109 analyzed strains were isolated predominantly from patients aged from 60 to 69 years. There were no significant differences in isolation rates depending on gender (male: female ratio: 1.08, p>0.05).
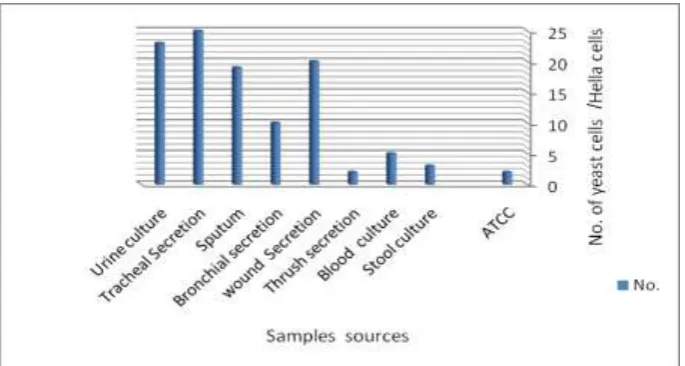
The majority of the selected C. albicans strains were isolated from upper respiratory tract
Fig 1: Distribution of C. albicans strains according to the isolation source.
All tested strains adhered to the cellular substratum, but with different intensities (AIn) (from 4.7 to 7.9 yeast cells/eukaryotic cells) (Fig. 2).
Fig. 2 Graphic representation of the Ain of C. albicans strains isolated from different clinical specimens to the HeLa cells.
[image:7.595.128.471.394.577.2]biofilm than those from non-invasive infections.[33] The highest biofilm producers were isolated from URTS and urine and correlated with high AIn.
Fig 3: Distribution of C. albicans strains according to origin source and capacity to develop biofilm to inert substratum (none/low/moderate/high).
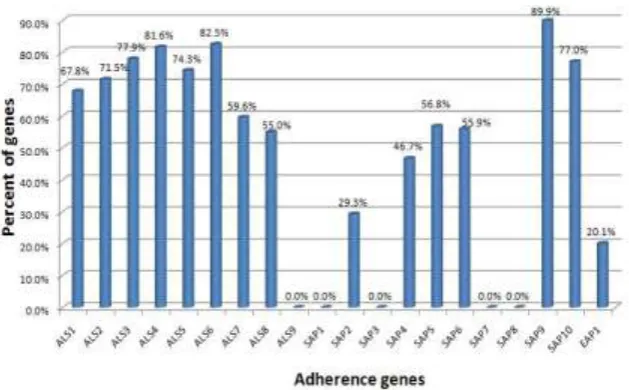
Fig (4) Distribution of C. albicans adhesions genes among different clinical isolates.
The most frequently encountered adherence genes were ALS3 and SAP9, suggesting that these genes significantly contribute to the overall virulence of C. albicans strains isolated from symptomatic infections, presumably by facilitating the adherence to cellular and inert substrata.
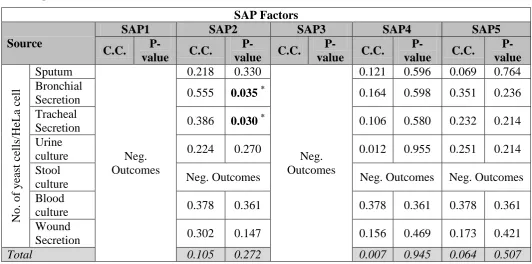
[image:8.595.144.479.132.284.2] [image:8.595.126.441.350.545.2]of urine isolates, ALS3 gene for sputum and tracheal isolates, and SAP 2 for bronchial and tracheal secretions, which correlated with a high value of the Ain (P<0.05).
Table (1): Association between No. of yeast cells/ HeLa cell and the presence of ALS and EAP genes in different clinical isolates.
ALS Factors Source
ALS1 ALS 2 ALS 3 ALS 4 ALS 5
C.C. P-value C.C.
P-value C.C. P-value C.C.
P-value C.C. P-value No. of ye ast c ell s/HeL a ce ll
Sputum 0.031 0.891 0.016 0.943 0.517 0.008* 0.016 0.943 0.184 0.414 Bronchial
Secretion 0.000 1.000 0.102 0.747 0.243 0.429 0.378 0.197 0.243 0.429 Tracheal
Secretion 0.302 0.100 0.155 0.414 0.386 0.030
*
0.060 0.756 0.064 0.738 Urine
culture 0.450 0.016
*
0.227 0.263 0.248 0.219 0.227 0.263 0.292 0.144
Stool
culture Neg. Outcomes Neg.
Outcomes Neg. Outcomes
Neg. Outcomes
Neg. Outcomes Blood
culture 0.243 0.329 Wound
Secretion 0.171 0.436 0.190 0.375 0.072 0.740 0.050 0.819 0.228 0.283 Total 0.074 0.442 0.049 0.607 0.108 0.259 0.133 0.163 0.024 0.806
Source
ALS 6 ALS 7 ALS 8 ALS 9 EAP1
C.C. P-value C.C.
P-value C.C.
P-value C.C.
P-value C.C. P-value No. of ye ast c ell s/HeL a ce ll
Sputum 0.184 0.414 0.218 0.330 0.077 0.737
Neg. Outcomes
0.251 0.259 Bronchial
Secretion 0.164 0.598 0.311 0.301 0.000 1.000 0.311 0.301 Tracheal
Secretion 0.205 0.277 0.103 0.590 0.025 0.895 0.009 0.963 Urine culture 0.227 0.263 0.337 0.086 0.178 0.385 0.175 0.394
Stool culture
Neg. Outcomes
Neg. Outcomes
Neg.
Outcomes Neg.
Outcomes Blood
culture 0.243 0.576 0.522 0.171
Wound
Table (2): Association between No. of yeast cells/HeLa cell and the presence of SAP genes in different clinical isolates.
SAP Factors
Source
SAP1 SAP2 SAP3 SAP4 SAP5
C.C.
P-value C.C.
P-value C.C.
P-value C.C.
P-value C.C.
P-value No. of ye ast c ell s/HeL a ce ll Sputum Neg. Outcomes
0.218 0.330
Neg. Outcomes
0.121 0.596 0.069 0.764 Bronchial
Secretion 0.555 0.035
*
0.164 0.598 0.351 0.236 Tracheal
Secretion 0.386 0.030
*
0.106 0.580 0.232 0.214
Urine
culture 0.224 0.270 0.012 0.955 0.251 0.214
Stool
culture Neg. Outcomes Neg. Outcomes Neg. Outcomes
Blood
culture 0.378 0.361 0.378 0.361 0.378 0.361
Wound
Secretion 0.302 0.147 0.156 0.469 0.173 0.421
Total 0.105 0.272 0.007 0.945 0.064 0.507
C. albicans adherence have been performed using Secreted Aspartyl Proteinases (Sap), Our
study is indicate that the C. albicans SAP gene family is differentially expressed in the yeast and hyphal, and may contribute to C. albicans adherence in different levels. Adhesion of Candida to host tissues allows the fungus to attain a foothold and to colonize a specific niche environment., this colonized site provides the base for candidal proliferation, invasion, and, in some instances, dissemination.[34]
Source
SAP6 SAP7 SAP8 ASP9 SAP10
C.C. P-value C.C.
P-value C.C.
P-value C.C.
P-value C.C. P-value No. of ye ast c ell s/HeL a ce
ll Sputum 0.346 0.107
Neg. Outcomes
Neg. Outcomes
Neg.
Outcomes 0.251 0.259 Bronchial
Secretion 0.311 0.301 0.164 0.598 0.000 1.000
Tracheal
Secretion 0.106 0.580 0.205 0.277 0.014 0.943
Urine
culture 0.322 0.103 0.082 0.692 0.148 0.472
Stool
culture Neg. Outcomes
Neg. Outcomes
[image:10.595.31.564.106.370.2]No significant associations were revealed among the capacity of biofilm forming on the inert substratum and different genes involved in adherence, excepting ALS1 for tracheal and ALS8 for urinary isolates (P<0.05).
Table (3): Association between the capacity to form biofilms on the inert substratum and the presence of ALS and EAP genes in different clinical isolates.
ALS Factors
Source
ALS1 ALS 2 ALS 3 ALS 4 ALS 5
C.C. P-value C.C.
P-value C.C.
P-value C.C.
P-value C.C. P-value B iofilm to i ne rt subst ra tu m
Sputum 0.031 0.891 0.016 0.943 0.184 0.414 0.282 0.200 0.204 0.364 Bronchial
Secretion Neg. Outcomes
Neg. Outcomes Neg. Outcomes Neg. Outcomes Neg. Outcomes Tracheal
Secretion 0.475 0.019
*
0.215 0.521 0.388 0.091 0.146 0.746 0.195 0.586 Urine
culture 0.145 0.781 0.214 0.575 0.086 0.918 0.272 0.399 0.310 0.293 Stool
culture 0.447 0.386 Neg. Outcomes
0.447 0.386
Neg. Outcomes
0.707 0.083
Blood
culture 0.243 0.576
Neg. Outcomes
Neg. Outcomes Wound
Secretion 0.234 0.543 0.405 0.127 0.210 0.643 0.371 0.186 0.344 0.243 Total 0.117 0.683 0.212 0.164 0.040 0.981 0.185 0.283 0.062 0.936
Source
ALS 6 ALS 7 ALS 8 ALS 9 EAP1
C.C. P-value C.C.
P-value C.C.
P-value C.C.
P-value C.C. P-value B iofilm to i ne rt subst ra tu m
Sputum 0.184 0.414 0.218 0.330 0.346 0.107
Neg. Outcomes
0.369 0.084 Bronchial
Secretion Neg. Outcomes
Neg. Outcomes Neg. Outcomes Neg. Outcomes Tracheal
Secretion 0.146 0.746 0.111 0.844 0.231 0.468 0.171 0.665 Urine
culture 0.272 0.399 0.271 0.402 0.539
0.009
* 0.211 0.584
Stool
culture 0.707 0.083 0.707 0.083 0.707 0.083 0.447 0.386 Blood
culture Neg. Outcomes 0.243 0.576 0.378 0.361
Neg. Outcomes Wound
Secretion 0.338 0.258 0.371 0.186 0.405 0.127 0.155 0.781
Total 0.106 0.746 0.224 0.128 0.294 0.017* 0.125 0.631
are implicated in attachment to endothelial cells and biofilm formation. The presence of the gene has been reported in previous study.[35]
Also our study show that C. albicans ALS8 gene play acritcal role n adherence and boflm production ,considered as low and high biofilm producers, this disagree with previous study that show ALS3 gene polymorphism was also encode among the different sources of isolation: blood, urine culture , vulvovaginal secretions wich determine the production of biofilm.[36]
Table (4): Association between the capacity to form biofilms on the inert substratum and the presence of SAP genes in different clinical isolates.
SAP Factors
Source SAP 1 SAP 2 SAP 3 SAP 4 SAP 5
C.C. P-value C.C.
P-value C.C.
P-value C.C.
P-value C.C. P-value B iofilm to i ne rt subst ra tu m Sputum Neg. Outcomes
0.129 0.570
Neg. Outcomes
0.165 0.466 0.251 0.259 Bronchial Secretion Neg. Outcomes Neg. Outcomes Neg. Outcomes Tracheal
Secretion 0.212 0.529 0.263 0.366 0.212 0.529
Urine culture 0.246 0.478 0.158 0.744 0.351 0.198
Stool culture 0.447 0.386 0.707 0.083 0.707 0.083
Blood culture 0.378 0.361 0.378 0.361 0.378 0.361
Wound
Secretion 0.325 0.308 0.170 0.742 0.307 0.353
Total 0.199 0.211 0.085 0.849 0.209 0.174
Source
SAP 6 SAP 7 SAP 8 SAP 9 SAP 10
C.C. P-value C.C.
P-value C.C.
P-value C.C.
P-value C.C. P-value B iofilm to i ne rt subst ra tu m
Sputum 0.077 0.737
Neg. Outcomes Neg. Outcomes Neg. Outcomes
0.069 0.764 Bronchial
Secretion Neg. Outcomes
Neg. Outcomes Tracheal
Secretion 0.015 0.997 0.146 0.746 0.183 0.627
Urine
culture 0.414 0.093 0.421 0.083 0.214 0.575
Stool
culture 0.447 0.386
Neg. Outcomes
Neg. Outcomes Blood
culture 0.522 0.171 0.243 0.576 0.243 0.576
Wound
Secretion 0.181 0.712 0.369 0.206 0.178 0.734
DISCUSSION
The increased incidence of systemic mycoses caused by Candida albicans in hospitalized patients is an important cause of morbidity and mortality worldwide, especially in critically ill patients. Peterson et al.[37] observed that the occurrence of oral yeasts from saliva of hospitalized patients was 55%. In patients with advanced cancer, the incidence ranged between 47% and 87% of the population. In diabetic patients, the presence of Candida spp. in the oral mucosa reached up to 80%.[38] In our study, C. albicans strains were isolated from hospitalized patients predominantly from tracheal secretions (22.9%), followed by urine
culture (21.1%) and wound secretions (18.3%).
One of the most important category of virulence factors in C. albicans strains is represented by the capacity to colonize the host tissues.[37] Biofilms also play an important role in the perpetuation of these infections primarily with respect to their ability to adhere to various medical devices.
Although C. albicans adhesion to host tissues contributes essentially to its virulence and pathogenicity, we know relatively little about the molecular mechanisms governing C. albicans adhesion to materials or mammalian cells, as well as about the correlation between
phenotypic and genotypic aspects of this phenomenon.
The study was performed on a significant number of recent C. albicans clinical isolates harvested from patients hospitalized for cardiovascular surgery. Our findings indicated that the C. albicans strains isolated from different clinical specimens exhibited specific adherence to cellular and inert substrata and biofilm formation properties, also revealing specific adherence gene profiles. . In this study, we identified C. albicans ALS1,ALS3 gene that enhances cell adhesion to polystyrene as well as to human epithelial cells respectively . other study done by[39] ALS genes was detected across the set of clinical specimens, So other studies ALS1, ALS3, and ALS9 were detected most frequently among these clincal samples.
From this study it has been found that C. albians SAP9 gene isolated from diffrents clinical
with strains harboring the SAP9 adherence gene (90 %), followed by SAP2, revealed by the tracheal and bronchial isolates.
CONCLUSION
Our study represents one of the first reports on the evaluation of the correlation between different phenotypic features involved in adherence and biofilm development and the presence of different adherence genes in C. albicans clinical strains. We have found a specific correlation between the ability to colonize the cellular substratum and the presence of ALS1 gene in case of urine isolates, of ALS3 in sputum and tracheal isolates, and of SAP 2 in bronchial and tracheal secretions. Also specific correlations between the biofilm development capacity and the presence of ALS1 in tracheal and of ALS8 in urinary isolates have been revealed. These results demonstrate that the detection of these genes in different clinical isolates could represent and easy and cost-effective tool for the prediction of highly virulent isolated capable of colonizing either the cellular or the inert substratum and to generate clinical infections.
ACKNOWLEDGMENTS
This work has been done at the University of Bucharest –ICUB-Faculty of Biology, We apologize to all colleagues in the Departement of Microbiology –Faculty of Biology whose work we were unable to cite due to space limitations.
REFERENCES
1. Arendorf TM, Walker DM. The prevalence and intra-oral distribution of Candida albicans in man. Arch Oral Biol, 1980; 25: 1–10.
2. Belazi M, Velegraki A, Fleva A, et al. Candidal overgrowth in diabetic patients: potential predisposing factors. Mycoses, 2005; 48: 192–196.
3. Jabra-Rizk MA, Falkler WA Jr, Merz WG, et al. Coaggregation of Candida dubliniensis with Fusobacterium nucleatum.J Clin Microbiol, 1999; 37: 1464–1468.
4. Velegraki A, Nicolatou O, Theodoridou M, Mostrou G, Legakis NJ. Paediatric
AIDS-related linear gingival erythema: a form of erythematous candidiasis? J Oral Pathol Med, 1999; 28: 178–182.
5. Baumgartner JC, Watts CM, Xia T. Occurrence of Candida albicans in infections of endodontic origin. J Endod, 2000; 26: 695-698.
7. Yuan K, Chang CJ, Hsu PC et al. Detection of putative periodontal pathogens in non-insulin-dependent diabetes mellitus and non-diabetes mellitus by polymerase chain reaction. J Periodontal Res, 2001; 36: 18–24.
8. Portela MB, Souza IP, Costa EM et al. Differential recovery of Candida species from subgingival sites in human immunodefi ciency virus-positive and healthy children from Rio de Janeiro, Brazil. J Clin Microbiol, 2004; 42: 5925–5927.
9. Song X, Eribe ER, Sun J, Hansen BF, Olsen I. Genetic relatedness of oral yeasts within and between patients with marginal periodontitis and subjects with oral health. J
Periodontal Res, 2005; 40: 446–452.
10.Darwazeh AM, MacFarlane TW, Mc Cuish A, Lamey PJ. Mixed salivary glucose levels
and candidal carriage in patients with diabetes mellitus. J Oral Pathol Med, 1991; 20: 280–283.
11.Willis AM, Coulter WA, Hayes JR, Bell P, Lamey PJ. Factors affecting the adhesion of Candida albicans to epithelial cells of insulin-using diabetes mellitus patients. J Med Microbiol, 2000; 49: 291–293.
12.Willis AM, Coulter WA, Fulton CR et al. The infl uence of antifungal drugs on virulence properties of Candida albicans in patients with diabetes mellitus. Oral Surg Oral Med Oral Pathol Oral Radiol Endod, 2001; 91: 317–321.
13.Sardi JC, Duque C, Mariano FS, et al. Candida spp. in periodontal disease: a brief review. J Oral Sci, 2010; 52: 177–185.
14.Hube B, Naglik J. Candida albicans proteinases: resolving the mystery of a gene family. Microbiology 2001; 147: 1997–2005.
15.Ramanan N, Wang Y. A high-affi nity iron permease essential for Candida albicans virulence. Science, 2000; 288: 1062–1064.
16.Tsang CS, Chu FC, Leung WK et al. Phospholipase, proteinase and haemolytic activities of Candida albicans isolated from oral cavities of patients with type 2 diabetes mellitus. J Med Microbiol, 2007; 56: 1393–1398.
17.Manfredi M, McCullough MJ, Al-Karaawi ZM, Vescovi P, Porter SR. In vitro evaluation
of virulence attributes of Candida spp. isolated from patients affected by diabetes mellitus. Oral Microbiol Immunol, 2006; 21: 183–189.
18.Hazen KC, Glee PM. Cell surface hydrophobicity and medically important fungi. Curr Top Med Mycol, 1995; 6: 1–31.
20.Dumitru R, Hornby JM, Nickerson KW. Defi ned anaerobic growth medium for studying Candida albicans basic biology and resistance to eight antifungal drugs. Antimicrob Agents Chemother, 2004; 48: 2350–2354.
21.Jamieson DJ, Stephen DW, Terri è re EC. Analysis of the adaptive oxidative stress response of Candida albicans. FEMS Microbiol Lett, 1996; 138: 83–88.
22.Rosa EA, Rached RN, Ign á cio SA, et al. Phenotypic evaluation of the effect of anaerobiosis on some virulence attributes of Candida albicans. J Med Microbiol, 2008; 57: 1277–1281.
23.Pizzo G, Barchiesi F, Falconi Di Francesco L, et al. Genotyping and antifungal susceptibility of human subgingival Candida albicans isolates. Arch Oral Biol, 2002; 47:
189–196.
24.Ghannoum, M., and K. Abu Elteen. 1986. Correlative relationship between proteinase production, adherence and pathogenicity of various strains of Candida albicans. J. Med. Vet. Mycol. 24:407–413
25.Abu-Elteen, K. H., A. Z. Elkarmi, and M. Hamad. 2001. Characterization of phenotype-based pathogenic determinants of various Candida albicans strains in Jordan. Jpn. J. Infect. Dis, 54: 229–236.
26.Pichova, I., L. Pavlickova, J. Dostal, E. Dolejsi, O. Hruskova-Heidingsfeldova, J. Weber, T. Ruml, and M. Soucek. 2001. Secreted aspartic proteases of Candida albicans, Candida tropicalis, Candida parapsilosis and Candida lusitaniae. Inhibition with peptidomimetic inhibitors. Eur. J. Biochem, 268: 2669–2677.
27.Borg, M., and R. Ru¨chel. 1988. Expression of extracellular acid proteinase by proteolytic Candida spp. during experimental infection of oral mucosa. Infect. Immun, 56: 626–631 28.el-Maghrabi, E. A., D. M. Dixon, and J. W. Burnett. 1990. Characterization of Candida
albicans epidermolytic proteases and their role in yeast-cell adherence to keratinocytes. Clin. Exp. Dermatol, 15: 183–191.
29.Ollert, M. W., R. Sohnchen, H. C. Korting, U. Ollert, S. Brautigam, and W. Brautigam. 1993. Mechanisms of adherence of Candida albicans to cultured human epidermal
keratinocytes. Infect. Immun, 61: 4560–4568.
31.Sardi JC, Duque C, Camargo GA, Hofl ing JF, Gon ç alves RB. Periodontal conditions and prevalence of putative periodontopathogens and Candida spp. in insulin-dependent type 2 diabetic and non-diabetic patients with chronic periodontitis. A pilot study. Arch Oral Biol, 2011; 56: 1098–1105.
32.Rodrigues AG, M å rdh PA, Pina-Vaz C, Martinez-de-Oliveira J, Fonseca AF. Germ tube formation changes surface hydrophobicity of Candida cells. Infect Dis Obstet Gynecol, 1999; 7: 222–226.
33.McCullough MJ, Clemons KV, Stevens DA. Molecular and phenotypic characterization
of genotypic Candida albicans subgroups and comparison with Candida dubliniensis and Candida stellatoidea. J Clin Microbiol, 1999; 37: 417–421.
34.Ellepola ANB, Samaranayake LP. Oral candidal infections and antimycotics CritRev Oral Biol Med, 2000; 11: 172–198.
35.Peterson DE, Minah GE, Overholser CD, et al. Microbiology of acute periodontal infection in myelosuppressed cancer patients. J Clin Oncol, 1987; 5: 1461–1468.
36.Soysa NS, Samaranayake LP, Ellepola AN. Diabetes mellitus as a contributory factor in oral candidosis. Diabet Med, 2006; 23: 455–459.
37.Kuhn DM, Chandra J, Mukherjee PK, Ghannoum MA. Comparison of biofilms formed by Candida albicans and Candida parapsilosis on bioprosthetic surfaces. Infec Immun, 2002; 70: 878-888.
38.Cho T, Hamatake H, Kaminishi H, Hagihara Y, Watanabe K. The relationship between cyclic adenosine 3Æ, 5Æ-monophosphate and morphology in exponential phase Candida albicans. J Med Vet Mycol, 1992; 30: 35–42.
39.Abad-Zapatero, C., R. Goldman, S. W. Muchmore, C. Hutchins, K. Stewart, J. Navaza, C. D. Payne, and T. L. Ray. 1996. Structure of a secreted aspartic protease from C. albicans complexed with a potent inhibitor.
40.Louise O’ Connora,, Sine´ad Lahiffa, Fiona Caseya, Maura Glennona, Martin Cormicanb, Majella Mahera. Quantification of ALS1 gene expression in Candida albicans biofilms by RT-PCR using hybridisation probes on the LightCyclere. Molecular and Cellular Probes,
2005; 19: 153–162.
42.Samaranayake YH, Dassanayake RS, Cheung BP, et al. Differential phospholipase gene expression by Candida albicans in artifi cial media and cultured human oral epithelium. APMIS, 2006; 114: 857–866.
43.Ogasawara A, Odahara K, Toume M, et al. Change in the respiration system of Candida albicans in the lag and log growth phase. Biol Pharm Bull, 2006; 29: 448–450.
44.Georgina Cheng, Karen Wozniak, Matthew A. Wallig,1 Paul L. Fidel, Jr.,2 Suzanne R. Trupin, and Lois L. Hoyer,*.Comparison between Candida albicans Agglutinin-Like Sequence Gene Expression Patterns in Human Clinical Specimens and Models of Vaginal