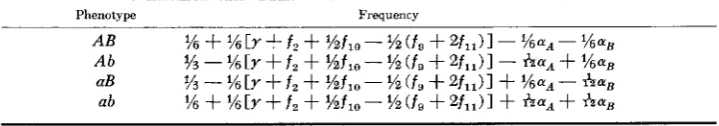THE ESTIMATION OF LINKAGE IN AUTOTETRAPLOIDS
FR.0M SINGLE BACKCROSS DATA
IAN R. FRANKLIN1
Division of Plant Industry, C.S.I.R.O. Canberra, A.C.T., Australia and Genetics Department, Uniuersity of California, Berkeley, California 94720’
Received November 7, 1966
HE estimation of the recombination fraction for two loci in autotetraploids Twas first considered by DE WINTON and HALDANE (1931), who derived expec- tations for the four phenotypes observed with intercrosses and backcrosses of some of the doubly heterozygous genotypes. These authors assumed that two sister chromatids canna4 enter the same gamete, and their calculations are equiva- lent to assuming that the four chromosomes pair as two bivalents at the first division of meiosis. This will be referred to as chromosome segregation.
The expectations for chromatid segregation, where two chromatids are chosen
at random from the eight strands segregating at meiosis, were published by SANSOME (1 933). SANOOME, however, failed to take into account the number of ways in which crossovers between two of the four chromosomes will lead to detectable recombinant strands. For example, in the bisimplex coupling genotype (AB/ab/ab/ab)
,
three (of the six crossovers involving different pairs will give rise to new combinations, but for bisimplex repulsion (Ab/aB/ab/ab) only one of the six will result in detectable recombinants. Using SANSOME’S expectations, there-fore, an estimate of the recombination from bisimplex coupling data would be approximately three times that obtained using bisimplex repulsion data.
MATHER (1936) and FISHER and MATHER (1943) showed more clearly the nature of the assumptions made by the previous authors. They concluded that the gametic frequencies from a heterozygous tetrasomic genotype would be inter- mediate between those expected from chromosome and chromatid segregations, and they introduced a parameter (a) known as the frequency of double reduction
-a measure of the frelquency with which a gamete receives two alleles derived from the same chromosome. (Y is a function of the number of crossovers between
the locus and the centromere, and the frequency with which paired chromosomes go to the same pole at ,anaphase I, and hence has a unique value f o r each locus. The gametic output of any tetrasomic genotype can be expressed in terms of a
(Table 1 ) .
MATHER also calculated expectations for backcrosses of the two bisimplex genotypes, which he felt would give estimates of the recombination fraction closely approximating the true value for small map distances. Because of their complicated nature, these expectations are more conveniently discussed in a later section.
’ Piesent address
5
92 I. R. FRANKLINTABLE 1
The expected frequencies of gametes at a single tetrasomic locus
Gametes
Parental eenotvm A A Aa M
Nulliplex (a")
Simplex (Aaaa)
Duplex ( A A a a )
Triplex (AAAa)
Quadriplex ( A A A A )
. . a
4
I + &
6
2 + a
. .
2 - 2 a
4 4-4tY
6
2 - 2 a
4 1 4 . . 4 . .
In 1947 FISHER demonstrated that unbiassed estimates of the recombination
fraction could not be obtained from single backcross data, and showed that a double backcross could yield data from which the recombination fraction can be
estimated. This procedure, however, involves the raising and identification of a very large number of individuals, and the work involved in the calculation of linkage maps in autotetraploids would be prohibitive in all but a few experimental organisms.
It was felt, therefore, that the procedure for the estimation of linkage in auto- tetraploids should be re-examined in the light of the treatment by FISHER in order to determine how closely estimates from single backcross data can approximate the recombination fraction.
Fisher's treatment: A doubly tetragenic genotype, i.e., one in which each of the two loci has four different alleles, can give rise to 136 different gametic geno- types, and these can be classified into I1 modes of formation. For example, from
TABLE 2
The modes of gamete formation for two tetrasomic loci
Double reduction Number of
Mode Type of gamete Number Frequency recombinant strands A locus B locus
. . . . . .
1 a 1 b J a A 6 f l
2 a,b*/a,b, 4 f 2
3 a 1 b 1 / a 2 b 3 24 f ,
4 albJalb2 12 f ,
5 a1bJa2b1 12 f ,
6 a1b2/a3b4 12 f ,
7 a2b1/a3b1 12 f 7
8 b 2 / a l b3 12 f ,
9 a1bJa2b3 24 f ,
10 a,b2/a,b2 12 f l , 2 1 1
1 1 alb2/a2bl 6 f l l
. . 1 1
1 . . . .
1 1 . .
1 . . 1
2 . . . .
2 . . 1
2 1 . .
2 . . . .
2 . . . .
LINKAGE IN AUTOTETRAPLOIDS 593
the genotype a,bJa,b:,/a3b3/a,b4 the gametic genotypes albl/a2b2, albl/a,b3, albl/a4b4, a2b2/a3b,, a2b2/a,b4, a3b3/a4b4 represent a single mode of formation, as each of these genotypes can be obtained from the other by permutation of the four chromosomes from which these gametes arise.
The 11 modes of gamete formation for two loci in autotetraploids are listed in Table 2. It can be seen that the recombination fraction is estimated by
y = f ‘/f4
+
%?f6 f f 6 f f i+
f 8+
f 9 f f l 0+
f l land the frequencies of double reduction at the A locus ( a A ) and at the B locus ( w ) are
a A = f 2
+
f 4+
f 8+
f l 0a B = f 2
+
f 5 f f i+
f l 0The reason that an unbiassed estimate of the recombination fraction cannot be
estimated from single backcross data becomes immediately obvious. Modes 1 and 11, which do not make the same contribution to the recombination fraction, have the same genic content, and cannot be distinguished except by progeny testing to determine whether the strands are parental or recombinant. Similarly modes 3 and 9 will always be confounded in single backcross data.
FISHER (1947) showled how to obtain an unbiassed estimate of the recombina- tion fraction using double backcross data from any but one of the doubly hetero- zygous genotypes. The ,procedure includes identification of the gametic genotypes (ten in the case of the doubly digenic genotypes) by progeny testing the first backcross individuals, and from the frequencies of the classes, a number of combi- nations of the mode frequencies can be estimated. The recombination fraction and the frequencies of double reduction are then simply calculated.
In the following section the estimable combinations of the mode frequencies from only four classes (the phenotypes observed from a single backcross), are discussed.
Expectations for single backcross data: Consider the bisimplex coupling geno- type AB/ab/ab/ab, where at the two loci A is dominant to a, and B is dominant to b. Among the progeny of a backcross to the homozygous recessive genotype (ab/ab/ab/ab) there will be four phenotypes, the expectations of which, in terms of the frequencies of the 1 1 modes of gamete formation, are shown in Table 3 .
These expectations can be quite simply calculated. For example, of the six ways in which gametes of mode 1 (no crossovers) can be formed, three will be AB/ab
TABLE 3
594 I. R. F R A N K L I N
TABLE 4
Expected frequencies for a bisimplex coupling backcross rearranged to show estimable combinations
Phenotype Frequency
and three ab/ab. The terms of Table 3 are conveniently obtained from the gametic matrices of FISHER (1947).
From the four classes, three combinations of the mode frequencies can be esti- mated. As we know that the frequency of double reduction can be calculated from any heterozygous genotype, it is logical to choose as two of the estimable combinations f z
+
f 4+
f s+
flo, and f z+
f 5+
f 7+
flo. The expectations for thefour classes can therefore be rearranged as in Table 4.
W e can estimate the quantities f 2
+
f 4+
f s+
flo, f z+
f 5+
f 7+
flo, and %fZ+
%f3
+
%f,
+
%f5+
f 6+
f 7+
f s+
%fg+
flo
or, more simply Q, W, and y+
Sfz
-%
( f g+
2fll) where ( Y ~ , ( Y ~ , and y are as defined earlier. Similarly for thebisimplex repulsion backcross (Table 5) we can estimate aA, as, y
+
fz+
% f l o -1/2 ( f g
+
2fll). The backcross progeny of the other 17 doubly digenic genotypescan be expressed in a similar way (see Appendix). In each case there are three combinations of the mode frequencies which can be estimated. These have been arranged so that two represent the frequencies of double reduction at the two loci, and the third is shown for the 19 genotypes in Table 6. I t is the first seven of these, expressed in terms of the recombination fraction, which are of the most interest with respect to estimating linkage in autotetraploids.
Combining different types of single backcross data, such as bisimplex coupling and bitriplex coupling, an estimate of the recombination fraction can be obtained which is deficient in only two mode frequencies-mode 9 and mode 11. It can be seen further that by combining data from a minimum of six single backcrosses, e.g., bitriplex coupling and repulsion, triplosimplex repulsion (two genotypes),
TABLE 5
LINKAGE I N AUTOTETRAPLOIDS
TABLE 6
The estimable combinations of mode frequencies
595
Number of genotypes
Bisimplex coupling (AB/ab/ab/ab) Y
+
%f, - % (f,+
2fli) Bisimplex repulsion ( Ab/aB/ab/ab) Y+
f,+
%fi, - % (f,+
2fli) Simploduplex coupling ( A B / A b/ab/ab) Y+
%f,* -34
(f,+
Vi,)Simploduplex repulsion ( Ab/Ab/dB/ab) Y
+
f z+
f i n+
%fs' - % (f,+
2f11) Biduplex coupling (AB/AB/ah/ab) Y - %f, -34
(f,+
2fll)Biduplex repulsion ( A b / A b / d 3 / a B ) f,
+
f,+
fs+
2f1,Biduplex neutral (AB/Ab/nB/'ab) f,
+
%fs+
% f g+
f i n Triplosimplex coupling ( A B / A b / A b / a b ) 2 f,-ff,*Triplosimplex repulsion ( A b / A b / A b / a B ) 2 f,
+
f4*? f,'+
fl, Triploduplex coupling (AB/A.B/Ab/ab) 2 2f, - f,'Triploduplex repulsion ( A B / A b/Ab/&) 2 f,' +2fin
Bitriplex coupling ( AB/ AB/ AB/ ab) 1 f,
Bitriplex repulsion (AB/AB/Atb/aB) 1 f l 0 1
1 2
1
1 2
1
19
* If the loci are interchanged, f, becomes f 5 , f, becomes f,.
biduplex neutral, and bjduplex repulsion, estimates of nine quantities can be obtained. These are (fl 1-
fld,
f 2 , ( f 3+
f g ) , f4, f 5 ,f,,
f 7 ,fs,
flo.We can at best estimaie the quantity y -
i/z
( f g+
2f1,), andif
a limited num-ber of heterozygous genotypes are available for backcross, an estimate of y may be obtained which is more indeterminate. To date there have been very few experimental studies in which mode frequencies have been estimated in auto- tetraploids; hence, there is little experimental evidence concerning the relative importance of the mode frequencies which bias the estimation of linkage from single backcross data. FISHER ( 1949) estimated the recombination fraction, y , between two loci in Lythrum salicaria to be 0.3763 and estimated ( f g
+
2f11) as0.0396. These data suggest that single backcross data would underestimate the recombination fraction by only 2 centimorgans (cM) and hence would provide a very reasonable approxiimation.
However, the standard errors on these estimates are very large, and, in any case, we obviously cannot make general statements from this single experiment. In the next section, the way in which the modes of gamete formation are generated by crossing over and segregation at meiosis is discussed in order to obtain some information on the expected frequencies of the modes of gamete formation.
The expected frequencies of the modes of gamete formation: The frequencies of the 11 modes of gamete formation are dependent on a large number of factors. The most important of these are the probability of recombination between the two loci, the proba3ility of recombination between each locus and the centromere, the frequency of multivalent formation, and the disjunction of these multivalents
596 I. R. FRANKLIN
bivalent formation is assumed, hence paired chromosomes go to opposite poles at anaphase I. In the second model there is complete quadrivalent formation, and paired chromosomes can go to the same pole. Also, in this model the effect of partner exchange between the two loci is considered.
In all of the following discussions it is assumed that each gamete always con- tains two chromatids, i.e., numerical nondisjunction is ignored.
A. Bivalent formation. Consider two linked loci A and B, each with four alleles. A doubly tetragenic genotype can be represented as albl/aZbZ/a3b3/a4b4. If there are no crossovers between the loci, all of the gametes produced will be of the type albl/azb2, i.e., mode 1. If there is a single crossover between the loci, two gamete types are possible. A chromatid from each bivalent enters the same gamete, and there are 16 ways in which two chromatids can be chosen from eight in this manner. Of these, eight will be mode 1 and eight mode 3 . If it is assumed that each of these 16 has equal probability, and there is no a priori
'4rp "her Possible modes of gamete fomstion.
l o c u s A locusB Bivalent Alternate Adjacent
s- 1 1
1 1 3
0 a 1, 3 1, 3 1, 3, 5, U
v
1
3
1 1
3 1, 3, JJ.
1, 3, 6 1, 39 5, 6 ,
-
1, 3, 5 9 96
-
8 not po6Sible 1, 3, 5 , 7, 9,LINKAGE I N AUTOTETRAPLOIDS 597
reason why this should not be so, a single crossover will give
1/2
mode 1+
1/2
mode 3.
The consequences of two crossovers are shown in Figure 1. Given a single crossover between the two loci, there are eight ways in which a second can be arranged, one of which is a 2-strand double (type l ) , two are 3-strand doubles (type 2 ) , and five are 4-strand doubles (types 3 and 4 ) . For simplicity in the model it is assumed that each of these second crossovers are equally likely, although this assumption is certainly unrealistic. I t assumes no chromatid inter- ference, and more important, that the probability of two crossovers in one bivalent has the same probability as a single crossover in each bivalent.
Summing over the four types of double crossovers, weighted according to their frequency. we obtain
3/8
mode 1,%
mode 3 and1/8
mode 6. We can consider of gamete formation as the states of a first order Markov chain. The solution to this system isthe n crossovers (n=O, 1 , 2
. .
.) as a sequence of consecutive trials, and the modesf 1 = p o + 1/2p1+ % P A + . .
. . .
. +
% [ l + 2 ( i / ) a ] p n + . ..
f 3 = 1/2p2+.. . .
+
1 / 2 p n + .. .
f G =
l / g p : ! + .
. . . .
. +
% [ l - 2 ( 1 / ) " ] p . + .. .
Y = $'if3 + f e = = % p l + % p z . ..
+
1/2[1 - (i/)"Ipn+
. . .
and
where p. is the probability of n crossovers between the loci, and y is the recombi- nation fraction.
On the further assump tion that the crossovers are Poisson distributed, i.e.
pn = cmmn/n !
,
where m is the mean nuimber of crossovers between the loci, the recombination fraction becomes
and the three mode frequencies can be expressed simply in terms of y . y =
'/z
( 1 -e-%m)i.e. f l = (l-y)z
f 3 = 2y(l--Y)
f s = y 2
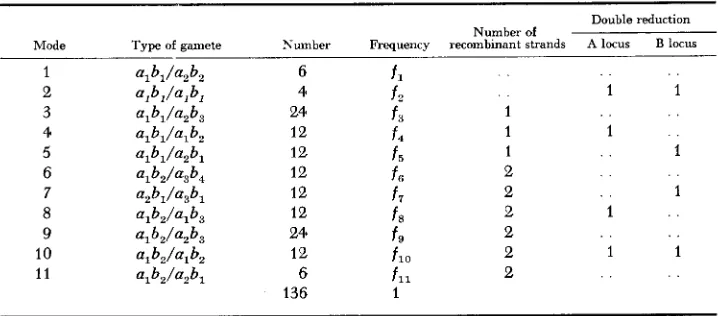
A plot of the mode frequencies against y is shown in Figure 2.
Since all modes except 1,3 and 6 are zero, the expectations of DE WINTON and HALDANE (1931) will estimate y exactly in the case of bisimplex and simplo- duplex genotypes. Their expectations for the biduplex genotypes will estimate y if f G = y 2 .
598 I. R. FRANKLIS-
0.7
0.6
V /
0 dl d2 d3 ok d5 Recombination Fraction lyl
FIGURE 2.-The expected mode frequencies assuming bivalent formation.
chromosome is paired for some of its length with one of the homologues, and another for the rest of the distance. The frequencies of modes 7, 8, and 9 are dependent upon partner exchange between the loci and will be zero if this does not occur. Since such a partner exchange must be accompanied by crossing over on both sides of the exchange to have any consequence in terms of the types of recombinant gametes which can be produced, this will be a rare event especially
if the loci are close together.
The position of the loci with respect to the centromere will also affect the fre- quencies of the modes of gamete formation. There are two distinct situations, one in which the loci are on the same arm of the chromosome, and the other in which the loci are on different arms (on either side of the centromere).
Consider two loci, A and B, on the same arm, with the A locus closest to the centromere. A single crossover between the two loci will result in two modes of gamete formation if the configuration is alternate, and four modes are possible with adjacent configuration. This is illustrated in Figure 1. The alternate con- figuration results in
and the adjacent configuration, which can orient in two different ways (assumed to be equally likely)
,
gives9/16 mode 1, 4/16 mode 3, 2/16 mode 5, 1/16 mode 11.
If we let the probabilities of these two configurations be I-c and c, respectively, we have
- B+cmode 1, 8 - 4 c m o d e 3 , 2 m o d e 5 , c m o d e l l .
-
% mode 1+
'/e mode 3LINKAGE I N AUTOTETRAPLOIDS 599
Although alternate arid adjacent configurations are not the only two possi- bilities at the first division of meiosis, all other configurations, provided that they give rise to euploid gametes, will disjoin in such a way that the disjunction can be expressed as a combination of them. The parameter c, therefore, although it will provide an approximate measure of the frequency of adjacent configuration, is more accurately equal to twice the frequency with which chromosomes. which are paired at some point, pass to the same pole at anaphase
I.
A single crossover can also occur between the A locus and the centromere. This will result in
1 6 % mode 1 -k mode 2.
16 16
This type of crossover is., of course, not possible if the loci are on different arms, and with this arrangement mode 2 requires at least two crossovers, one on each side of the centromere.
The results of double crossovers between the two loci are shown in Figure 1. There are now 24 ways in which a second crossover can be arranged, 16 of which are a result of partner exchange between the two loci. Eight of these are 3-strand doubles, and eight are &strand doubles (types
5
and 6). These 16 are not likely to be as frequent as nonpartner exchange doubles. With the exception of modes7, 8 and 9, however, the frequency of partner exchange has very little effect on the expected mode frequlencies.
Two graphs (Figures Sia, b) have been drawn to show the behavior of the mode frequencies as the map distances vary between the loci and between the loci and the centromere. These were obtained by enumerating all possible single, double and triple crossovers in the relevant regions and assuming a Poisson distribution of crossovers in each segment. A mean value of 0.03 partner exchanges per map unit, and a value of c =
q;
was chosen in the calculation of these figures. Figure 3a illustrates the situation when the loci are on different arms and equidistant from0.4 L
~
-I
0.2
O . 1
b
mo ell mode 4
0.1 az 0.3 0.4 0.5
Recombinohon Fraction lyl
FIGURE 3.-The expected mode frequencies assuming quadrivalent formation. a. Loci on
600 I . R. FRANKLIN
the centromere. Mode frequencies are plotted against y , the recombination fraction. In Figure 3b the loci are on the same arm, and the
A
locus, which is proximal, is approximately 20 map units from the centromere.It is obvious that in view of the large number of assumptions involved, the mode frequencies cannot be predicted from a knowledge of the recombination fraction alone, but a number of interesting observations can be drawn from these graphs. First, mode 10 takes a very low value, and it is not likely, even for large map distances, to make a significant difference to the estimation of the recombi- nation fraction. Similarly, if the loci are on different arms, mode 2 can also be ignored, but if they are on the same arm, mode 2 could cause a considerable disturbance to estimates of the recombination frequency where this mode fre- quency is confounded. In fact, the magnitude of mode 2 provides the most useful indication of whether the loci are on the same or different arms. The frequency of modes 7,8 and 9, which require at least one crossover on either side of a partner exchange in the region bounded by the two loci, will also have a small value, especially when the map distance is small. The significant mode frequencies, therefore, are 1,2,3,4,5,6 and 11.
The above conclusions will be even further strengthened when some of the restrictions in the model are relaxed. Bivalent formation will add to the impor- tance of modes 1,3, and 6 and decrease the value of the others. Also, the frequency of partner exchange, which has a high value in the above (a mean of three partner exchanges for a chromosome 100cM in length), will almost certainly be less than proposed; hence, modes 7, 8 and 9 will not be as frequent. Third, chiasma interference, which has been ignored to date, will reduce even further the frequency of those modes which require double and triple crossovers, namely 7, 8, 9 and 10.
It was shown previously that any estimate of the recombination fraction from single backcross data will be underestimated by the quantity
%
( f Q+
2f11). Therefore, mode 11 becomes the major factor in the indeterminacy of the recom- bination fraction. It can be seen from the graphs (Figures 3a, b) that when the loci are on different arms flli/z
( f4+
f 5 ) and on the same arm fll=
i/z
f5. This relationship is not surprising, as mode 11 arises in a very similar way to modes4
and 5 . (It is the separation at the second division which determines whether a gamete will be mode 4 or 5, or mode 11.) As the map distance, and hence the mean number of crossovers, increases, the approximation will not be as good, but only at recombination percentages of 40% o r more is the relationship likely to cease to become of value.DISCUSSION
LINKAGE I N AUTOTETRAPLOIDS 601
from simploduplex coupling as ysDc, etc. then ysDc, y B S C , ~ B s R , YSDR, YBDC, are
ranked in order of closest approximation to the recombination fraction ( y )
.
The position of the last is arlbitrary with respect to the previous three since the fre- quency of mode 2, which is confounded with y in yBSC, yBSR, and YSDR, bears norelation to the recombination fraction, and may be large or small depending on the positions of the loci and the frequency of quadrivalent formation. It would appear, however, that the estimate obtained from simploduplex data would pro- vide the least unbiassed estimate of y if other information were not available. For a given number of progeny the estimate with the lowest variance would be obtained from bisimplex coupling data. If we could be sure that the frequency of mode 2 was negligible, or could be corrected for, data from this type of cross would be the most valuable.
If
an experimenter were interested in the recombination fraction, he would presumably be interesteld in the frequencies of double reduction also, as these could provide some information about the position of the centromere (see below). The frequencies of double reduction are best estimated from bitriplex data. If we look at the expectations for bitriplex coupling, it is seen that three quantities can be estimated, namely an, 0 1 ~ and f 2 . Mode 2, which as mentioned previously pro-vides us with information about the position of the centromere with respect to the two loci, is one of the modes confounded with y in bisimplex coupling back- crosses. Hence, by combining data from these two backcrosses, we can obtain estimates of 01 l, an, f 2 arid y
-%
(f9+
2fll). If we wish to use the informationabout the expected frequencies of the mode frequencies, these four quantities are very useful in correcting for bias in estimates of y from single backcross data. The four parameters may be estimated from other combinations of backcross data, for example, bisimplex coupling and bisimplex repulsion, but many more progeny would be needed to estimate them with the same precision.
It was shown in a previous section that for small map distances we can assume that f 7 = f R = f 9 = flo = 0, and further that for loci on the same arm fll %f5,
and for different arms f l l r
'/2
(f4+
f 5 ) .Then 0 1 ~
=
f 2+
f 4 a13=
f z+
f5.These approximations suggest the following procedure in obtaining a n estimate of the map distance between two loci:
Y' = Y - %fs - f l l
(1) Estimate (from bisimplex coupling and bitriplex coupling data) aA, as, f,,
( 2 ) (a) If f,, aA, as are approximately zero, indicating a low frequency of quadrivalent for- mation, or that the loci are both close to the centromere, then
Y
=
Y'.Y zz Y'
+
%(.A+
" B ) .Y
=
Y'+
% (a' - f*)(b) If f, is zero, but aA tor aB have an appreciable value (say
>
3 % ) , indicating that theloci are on different arms, o r that one of the loci is close to the centromere
( c ) If f, is large, (i.e., of the order of the frequencies of double reduction), it indicates that
both loci are on the same arm, and in a distal position on the chromosome.
602 I. R. F R A N K L I N
It is difficult to decide how closely estimates from single backcross data will
approximate the recombination fraction. Certainly if the loci are loosely linked most of the mode frequencies will have appreciable values; hence, the above estimates will be seriously biassed. MATHER (1936) suggested that little faith can be placed in estimates of linkage values of more than 15% from single backcross data, but this author feels that this judgment is overcautious. The considerations above suggest that even unadjusted estimates, say from bisimplex coupling data, are not likely to be biassed by more than 3 or 4 map units if the recombination fraction is as high as 30%. If it is found that frequencies of double reduction have small values, this may be taken as further confidence in the estimates of the recombination percentage.
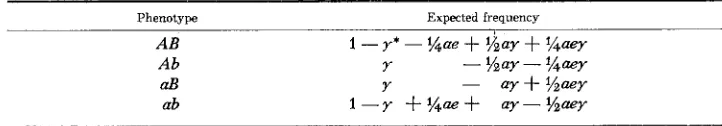
MATHER (1936) calculated expected frequencies for the progeny of a backcross of the two bisimplex genotypes, assuming that there will be only one crossover between the loci.
He
therefore suggested that these formulae be used only for closely linked loci. MATHER introduced two parameters, e-the frequency of equational separation at the locus nearest the centromere, and a-the frequency with which the centromeres of paired chromosomes pass to the same pole at anaphase I.The expectations for the bisimplex coupling genotype are shown in Table 7. The greatest difficulty in using these expected frequencies arises because they are calculated on the assumption that the two loci are located on the same chromo- some arm. If this is not the case, the expectations will be different, and it would be difficult to determine which model applies. Even if the loci are on the same arm it is necessary to know which locus is closest to the centromere, and this information could not be easily obtained from bisimplex coupling or bisimplex repulsion data. Further, this approach is not easily generalized to other back- crosses.
On the other hand, if the above information is available, and if the loci are closely linked, the procedure will allow the estimation of a and e as well as the recombination percent. These parameters are of considerable interest, since e is approximately twice the map distance between the locus and the centromere, and a is approximately half the frequency of adjacent-type configurations (i.e.,
a = $$c in the discussion of the section on expected frequencies of the modes of gamete formation.) Alternatively this information may be directly obtained from estimates of the recombination fraction and frequencies of double reduction, since
TABLE 7
The expected frequencies for the backcross of a bisimplex coupling genotype (MATHER 1936)
Phenotype Expected frequency
AB A b
aB
ab
1 -Y* - %Qe
+
%ay+
%aeYY - %ay - % a y
Y - a?+%aQY
LINKAGE I N AUTOTETRAPLOIDS 603
when the map distances are small, a increases linearly with the distance between the locus and its centrornere ( a = i/zc X map distance). Suppose therefore that we had estimates of aA, as, and yAB (the recombination fraction between the two loci)
,
and it was known that the loci were on different arms. Then we could esti- mate approximately the map distance of each locus from the centromere (yaand y n ) and c, as follows:
Y A E y A B a A / ( a A
+
a B ) Y A B a B / ( a A+
a s )c Q(a.4
+
a B ) / y A BThere are two problems which have not been considered at all in this paper. The first is the possibility of viability disturbances which will upset the estima- tion procedures outlined above. Since these disturbances are common to studies of linkage in diploids, estimation procedures will be similar, and will not be con- sidered here. The second is the possibility of numerical nondisjunction. This is a problem which would require a n extensive study, but it appears that the esti- mation of the recombination fraction would not be seriously affected, except perhaps from the simploduplex repulsion backcrosses. The estimation of the fre- quencies of double reduction, and particularly the frequency of mode 2 from the bitriplex backcross, will however be seriously biassed if numerical nondisjunction occurs, and the method of CATCHESIDE (1956) could be used to correct estimates of these quantities.
Finally, it should be pointed out that if the loci under investigation are com- pletely unlinked, i.e., on different chromosomes, the expected value of each of the mode frequencies can be calculated exactly in terms of the frequencies of double reduction at the two loci. The combination always confounded with the recombi- nation fraction (1 - aA - a B
+
aA X a B ) , and the quantity y - $$ (f9+
2f,,) will be$4
i-?4
( a A + a B - a A x a s ) .Since the recombination fraction for unlinked loci is equivalent to that given by two linked with a recombination fraction of 0.75 (SVED 1964)
,
the recombination frac:ion will be underestimated by as much as%.
( f 9
+
2f,,) is then equal toI am indebted to DRS. J. A. SVED, E. R. DEMPSTER and H. E. MCKEAN for helpful comments
throughout the preparation of this manuscript, and to MRS. BARBARA STRONG for the preparation
of the graphs.
SUMMARY
6M I. R. F R A N K L I N
APPENDIX: Expectations for Single Backcrosses
Bisimplex coupling (AB/ab/ab/ab)
AB 1 - y
+
'/z ( f s+
2f1,) - % f zA b Y - ? h a A - %
(f,
+
W,,)+
?hf,d Y - %ai3 - ?h ( f s
+
2f1,)+
% f 2ab 1 - Y
+
%aA+
%aB+
% ( f Q+
W,,)
- %f,Bisimplex repulsion (Ab/aB/ab/ab)
AB 1
+
Y-
(YE - % ( f s+
2fld+
f z+
% f l O A b 2 - Y - %.A+
as+
% ( f 9+
2fld - f z - %flO ab 1+
Y+
%a*+
%.o-
% ( f s+
'f,,)+
f z+
%f,oO A -
aB '-Y f ( y A - ? h a B f % (f, f 'fll) - f Z - sf10
Simploduplex coupling (AB/Ab/ab/ab)
A B 3 - Y - %.A- a B - % f 8 + % ( f Q + 2 f 1 1 )
Ab 2
+
Y - 3 / h A+
as+
% f s - % ( f s+
'fii) aB %a* - ! h a g+
% f g - % (f,+
2f11) ab 1 - Y+
3/hA+
X a B - % f s+
% ( f g+
'f11)Y
+
Simploduplex repulsion (Ab/Ab/aB/ab)
AB 2
+
Y-
3 / 2 a ~ - 3/&5+
f z+
f,o+
% f s - ?h ( f s+
2fii' A b 3 - Y - % a ~ + ~ / ~ B - f z - f 1 0 - % f g + ? h ( f ~ + ~ f l l ) aB 1 - Y+
3 / h A - f z-
f,o - %fa+
% (fQ+
'fldab Y + ?haA
+
f z+
f , O f % f 8-
?h ( f 9+
'fll)Biduplex coupling ( AB/AB/ab/ab)
AB 5 - 'Y - 'YA - a~
+
f 6+
( f g+
'f11) Ab 5 - a ~+
a B - f 6 - ( f g+
'f11)fd 2Y
+
" A - ( Y g - f 6-
(fs+
2f11) ab 1-
2Y+
( y A f QIB f f 6+
( f g+
'fll) Biduplex repulsion (Ab/Ab/aB/aB)AB 4 - "Y-A - h B
+
( f 6+
f 7+
f s+
'f10) Ab 1-
+
2a5-
( f 6+
f 7 f f s+
'fro)d 1 f b A
ab
- ( f 6
+
f T f f S+
'f10)+
(fs+
f 7+
f 8+
'f10) Biduplex neutral (AB/Ab/dB/ab)AB 8
-
3 a ~ - 31u,+
?h (f.3+
fs)+
( f z+
f i o )Ab 2
-
LyA+
3aB-
l/e (f.3+
f Q )-
( f z+
f10) aB 2+
3nA-
&E - '/e (f.3+
f Q )-
( f Z+
f 1 0 )ab 'YA
+
aB+
'/e (f,+
f,) f (fz+
f l 0 )Triplosimplex coupling (AB/Ab/Ab/ab) AB 6
-
a A - 3 a ~+
(fz - f 8 ) A b 6 - & A + 3 a B - ( ( f Z - f f s )ab 2aA
+
(fz - fs)aB aA - (fz
-
f 8 )Triplosimplex repulsion (Ab/Ab/Ab/aB) AB 2 - a ~ w a B - (fa f fro)
Ab 2
+
ad+
'ye-
(f,+
f,)dB (fz
+
f,)
L I N K A G E I N AUTOTETRAPLOIDS
Triploduplsx coupling (AB/,$B/aB/ab)
AB 10-4&A-2aB+ ( 2 f , - f 7 )
Ab 2aB
-
(2fZ - f 7 )d 2 + 4 u u A - a B - ( 2 f z - f 7 ) ab ( y B
+
( 2 f 2 - f 7 )Triploduplex repulsion ( A B / Ab/aB/aB)
AB 10
-
- 3a,j+
( f ? 2f10) A b 3%-
( f 7+
2flJaB 2
+
4a,,-
U,
+
2/10) ab+
(f,+
2fl")Bitriplex coupling (AB/ AB/ AB/ ab)
AB 4-un - ( ~ g
+
f z A b Qli - fzab an - f,
ab fZ
Bitriplex repulsion ( AB/AB/'aB/Ab)
AB4-a(yA-a(YH+i/f]0 A b & B - Y3f:,o
aA - %f.,o
ab Y3f LO
605
LITERATURE CITED
CATCHESIDE, D. G., 1956
DE WINTON, D., and J. B. S. HALLIANE, 1931
FISHER, R. A., 1947
Double reduction and numerical non-disjunction in tetraploid maize.
Heredity 10: 205-218.
Linkage i n the tetraploid Primula s i " k
J. Genet. 24: 121-144.
The theory of linkage in polysomic inheritance. Phil. Trans. Roy. Soc.
London B 233: 55-87. - 1949 The linkage problem in a tetrasomic wild plant,
Lythrum salican'a. Proc. 8th Intern. Congr. Genet. (Hereditas Suppl. Vol.) 225-233.
The inheritance of style length in Lythrum salicaria.
Ann. Eugenics 12: 1-23.
FISHER, R. A., and K. MATHER, 1943
MATHER, K., 1936
SANSOME, F. W., 1933 Chromatid segregation in Solanum lycopersicum. J. Genet. 2 7 : 105-132. SVED, J. A., 1964 The relationship between diploid and tetraploid recombination frequencies.
Segregation and linkage in autotetraploids. J. Genet. 32: 287-314.