0022-538X/81/090823-12$02.00/0 Vol. 39,No. 3
Mouse
Hepatitis Virus A59:
mRNA
Structure
and Genetic
Localization
of the
Sequence
Divergence from Hepatotropic
Strain
MHV-3
MICHAEL M. C. LAI,* PETER R. BRAYTON, ROBERT C. ARMEN, CHRIS D. PATTON,
CHARLES PUGH,tANDSTEPHEN A. STOHLMAN
Departmentsof Microbiology and Neurology, University of Southern California School ofMedicine, LosAngeles,California90033
Received 9April 1981/Accepted 26 May 1981
The composition and structure of the mouse hepatitis virus (MHV)-specific
RNA in actinomycin D-treated, infected L-2 cells were studied. Seven
virus-specific RNA species with molecular weights of0.6 x 106,0.9x 106,1.2 x 106,1.5
x 106,3.0 x 106,4.0x 106,and 5.4x 106 (equivalent to the viral genome) were
detected. T1oligonucleotide fingerprintingstudies suggested that the sequences
of each RNA speciesweretotally includedwithin the next larger RNA species.
Theoligonucleotidesof each RNAspecies were mapped on the 60S RNA genome
ofthe virus. Each RNAspecies contained the oligonucleotides startingfrom the
3'end of thegenomeandextending continuously for various lengths in the 3'
--5' direction.Allof the viral RNA species containedapolyadenylate stretch of100
to 130 nucleotides and probably identical sequences immediately next to the
polyadenylate. These data suggested that the virus-specific RNAs are mRNA's
and have astairlike structure similar tothat of infectious bronchitis virus, an
avian coronavirus. Aproposal is presented, basedonthemRNAstructure, for the
designation of thegenes onthe MHVgenome.Using thisproposal, thesequence
differences between A59, a weakly pathogenic strain, and MHV-3, a strongly
hepatotropic strain, werelocalizedprimarilyinmRNA's 1 and3,corresponding
togenesAand C.
Murine hepatitis viruses (MHVs) are
mem-bers of theCoronaviridae,whichinclude viruses
isolated frommanyspecies ofanimals (10, 18).
They contain a positive single-stranded 60S
RNAgenomewithamolecularweight of5.4 x
106 (6, 7, 19).Several other coronaviruses have
been reportedto have a similar genome
struc-ture(4, 8, 9).Aviralgenomeof this sizecancode
for all of the MHV structural proteins, which
includegpl80/90, pp5O,andgp23 (17), andmay
also haveenough informationtocode for several
nonstructural proteins (6). However, no
at-tempts have yet been made tolocalize the
ge-netic regions which code for thepossible gene
products.
Our laboratorieshavebeen interested in
iden-tifyingthegenetic regionsof the MHVgenome
associated with various viralproperties,
partic-ularly those responsible for the ability of the
virusto cause different diseases. Recently, we
have shown thatthegenomicRNAsofaweakly
pathogenicstrain, A59, andastrongly
hepatitis-inducing strain, MHV-3, have verysimilar
oli-gonucleotidefingerprintsexcept forafew
oligo-nucleotides (7). We have also found that a
de-myelination-inducing MHV strain, JHM-DS,
and aJHM strain whichcauses very little
de-myelination, JHM-DL, differ in onlyafew
oli-gonucleotides (S. A. Stohlman, M. M. C. Lai,
and L.Weiner,unpublisheddata). These viruses
thusprovideapotentialsystemfor the
identifi-cation of genetic regions responsible for viral
pathogenicity. As an initial step toward this purpose, we studied the structure of the
intra-cellular viral RNAs in order to identify the
mRNA specieswhicharetranscribedfrom such
geneticregions.
Several earlier reports have
attempted
tocharacterize thevirus-specific mRNA in
MHV-infected cells (11, 12). In these
studies,
only
afew RNA species of 18 to 28S were detected.
These RNAsare notlikelytorepresent the total
RNA species transcribed from the MHV
ge-nome. Recently, Stern and Kennedy (14, 15)
foundsix InRNAspeciesinthe avian
coronavi-rus-infected cells. Furthermore,
they
showedthatthesemRNA's hadanestedstrucutre.
Sev-eral laboratorieshavealso foundseven mRNA
speciesinMHV-infectedcells(13;J.
Leibowitz,
personal communication). In this report, we 823
on November 10, 2019 by guest
http://jvi.asm.org/
824 LAI ET AL.
show that themRNA's ofsimilarnature existin MHV-A59-infected cells.We have further
char-acterized their structure.
MATERIALS AND METHODS
Viruses andcells. The A59 strain of MHVwas
originallyobtained fromC.Bond,of theUniversityof California at SanDiego. This strain hasbeenshown tobeweakly pathogenic (11).Thehepatotropic MHV-3strainwasobtainedfrom M.Collins,of Microbiolog-ical Associates, Bethesda, Md. The L-2 cells were
obtained from L. Sturman,New YorkState Depart-mentofHealth,Albany,N.Y. The DBT cell linewas
obtained from K.Fujiwara, UniversityofTokyo,
Ja-pan.Viruseswereplaque purifiedonDBT cellsbefore
use.
Radiolabeling and purification of virus. For preparation of 32P-labeledviralRNA,theviruseswere grown onmonolayers of DBTcells in 15-cm dishes. Viruswasadsorbed atamultiplicityof infection ofone
tofive at370Cfor1h. Afteradsorption,the inoculum
wasremoved,and 25 ml of Dulbeccomodified
mini-mum essential media (DMEM) containing 1% heat-inactivated (56°Cfor 30min)fetal calfserum (FCS) wasadded.After 4 h at37°C,themediumwasremoved andreplacedwithphosphate-free DMEM which
con-tained 1% dialyzedFCS and 200,uCi of
'Pi
perml (ICNPharmaceuticals). Supernatantfluidswere col-lected 8 h later and clearedofcelldebrisby centrifu-gationat15,000x gfor30minat 40C. Viruswasthen pelletedandpurifiedbysedimentationon linear20 to 42%sucrosegradientsaspreviouslydescribed(6).The 32P-labeledRNAwasextractedfrom thepurifiedvirus accordingtopublished procedures (7).Preparation of intracellular virus-specific RNA. DBT and L-2cells wereused toprepare intra-cellular virus-specific RNA. Both cell lines were in-fected with MHV-A59atamultiplicityofonetofive for 1 h at 370C. After adsorption, DMEM which contained 2% dialyzedFCS and 1 to 3jigof actino-mycinDperml(a giftof MerckSharp&Dohme)was
added.At3to 4 hpostinfection,100,uCiof[3H]uridine
per mlwas added, and thevirus-specific RNAwas
extractedat6 to 7 hpostinfection.L-2cellswereused toprepare32P-labeled RNAbecause theRNA synthe-sis in DBTcells wasmarkedlyreducedwhen thecells were incubated in phosphate-depleted medium. L-2 cellswereincubated for 6to8 h inDMEMcontaining 2%dialyzedFCSand1/10of the normalconcentration ofphosphatebefore infection. Afterinfection,the L-2 cellswerecultured in phosphate-free DMEM which
contained 2%dialyzedFCS,3ugofactinomycinDper
ml,and 250 MCiof'Piperml.Virus-specificRNAwas
extracted from L-2cellsat7to9 hpostinfection. For extraction ofintracellular viral RNA,
mono-layersof infectedcellswerechilledonice and washed
three times withSSbuffer(10mMTris,60mMNaCl, and1mMEDTA, pH8.5).Thecellswerelysed in the
SSbuffercontaining0.5% TritonN-101, and the nuclei
wereremovedbycentrifugationat1,800xgfor5min.
The supernatant fluidwasthen adjustedto contain 1%sodiumdodecyl sulfate (SDS)andwasextracted
at 40Cwith chloroform-phenol (1:1). The RNA was
precipitated bythe addition of 2volumes ofethanolat -20°C overnight.
Initially, the RNA wasanalyzed byvelocity sedi-mentation in linear 15 to 30% sucrose gradients pre-pared in SS buffer (pH 8.0) which contained 0.5% SDS. Centrifugationwasperformed inanSW40 rotor at20,000 rpmfor 16 to 20 h at 200C. Fractions (0.2 ml) were collected withan ISCO fraction collector and were monitored by UV absorbance at 254 nm. 3p_ labeled DBT cellular RNAwasaddedtoprovide 28S, 18S, and4Ssedimentation markers.
Agarose gelelectrophoresis. Two different con-ditions were used for agarose gelelectrophoresis. In mostof theexperiments, the RNAsamples were dis-solved in RE buffer (50 mM boric acid,5mMsodium borate,1mMEDTA,and10mM sodiumsulfate, pH
8.1), heat denaturedby boilingat1000C for1min, and electrophoresed in 1% agarose gel slabs (15 by 20 by 0.3 cm) made in RE buffer at 90 V for 5 h. For determination of molecular weights of RNAs, the RNA samples were dissolved in RE buffer and electro-phoresed in agarose gel slabs made in RE buffer con-taining 5 mM methylmercuric hydroxide (2). After
electrophoresis, the gels were wrapped with cello-phane andexposed to Kodak XR film with an
inten-sifyingscreenfor variousperiods of time.
For preparativegelelectrophoresis, the RNA sam-pleswereelectrophoresedin agarosegels with or with-out methylmercuric hydroxide as described above. The RNA bandswereexcised from thegel, suspended in 2 to 3 ml of distilled water, and crushed with a Dounce homogenizer. For RNAs excised from the
methylmercury gels,the gel sliceswerefirst washed withtwochanges of 0.5 Manmmoniumacetatebefore homogenization.After centrifugation at 20,000xgfor 15min to remove thegel, the supernatant was adjusted to 0.15M ammonium acetate and precipitated with 2.5 volumes of ethanol. This extraction procedure removed at least 80% of the radioactivity. Further elution of thegel didnotremoveadditional RNA. The RNAwasprecipitated by sedimentation at 20,000xg for15min andredissolved in0.4ml of 0.2 M ammo-niumacetate. The RNA samples were again
centri-fugedat20,000xgfor15mintoremovecontaminating
gel pieces. The RNA in thesupernatantwasfinally precipitated with2.5volumes ofethanol.
Oligonucleotide fingerprinting.The32P-labeled
RNAextracted from purified virions or eluted from
gelswasdigestedwithRNaseT,and analyzed by two-dimensionalpolyacrylamide gel electrophoresis
essen-tially as previously described (7). Briefly, the first dimension was performed on 10% polyacrylamide-0.125% bisacrylamide gel slabsin citrate buffer, pH 3.3,containing6M urea.Electrophoresis was carried out at 700 V for 4 h. The second dimension was performedon22%polyacrylamide gel-0.15% bisacryl-amidegel slabs in Tris-borate buffer, pH 8.0, at 600 V for16h. Afterelectrophoresis, the gels were wrapped withcellophane and exposed to Kodak BB-1 film with anintensifyingscreenat40C.
Oligonucleotide mapping. The 32P-labeled 60S RNAwasdissolved in 0.3 ml of 0.05 M sodium carbon-ateandincubated at 50°C for 1 to 5 min. The reaction mixture was then neutralized with 15
pl
of 1M acetic acid andprecipitated with 2.5 volumes of ethanol. The RNAwaspelletedby sedimentation at 20,000xg for 15minandwasthen fractionated by oligodeoxythym-idylate[oligo(dT)]-cellulose column chromatography J. VIROL.on November 10, 2019 by guest
http://jvi.asm.org/
MHV-A59 mRNA STRUCTURE 825
accordingtotheprocedure ofAvivand Leder (1). In thelater partof theexperiments, the alkali digestion oftheRNAwasomitted. Instead,the total RNA from purified virus, without prior separation by sucrose
gradient sedimentation, was used directly for oligo(dT)-cellulose chromatography. The RNA
pre-pared by this procedure was degraded enough for oligonucleotide mapping. The polyadenylate [poly-(A)]-containing RNAselected by oligo(dT)-cellulose chromatography wassedimented ina 10 to25% su-crosegradient made in0.01MTris-hydrochloride,pH 7.4, 0.01 MNaCl, and 0.05% SDSinanSW41rotor at
40,000rpmfor 4.5 h. TheRNAs of differentsizeswere
pooled separatelyandwereused forT, oligonucleotide fingerprinting.
Basesequenceanalysis.Thepoly(A)-containing oligonucleotides separated by two-dimensional poly-acrylamide gel electrophoresiswereexcised from the
gels,eluted with 0.5 ml of 0.5 MNaCl, and precipitated with2.5volumes ofethanol. The oligonucleotideswere
pelleted by sedimentationat 20,000 xgfor 15mi, redissolvedin10plof waterwhichcontsinedRNase A at0.1 mg/ml, and incubated at370Cfor 30 min. Afterincubation, thedigestswerespottedon DEAE-cellulosepaperandelectrophoresedin acetic acid-pyr-idine buffer(pH 3.5)at1,500Vuntil the xylene-cyanol dyehad traveled 15cm. TheDEAE-cellulose paper wasdried andexposedtoKodakBB-1 film with inten-sifyingscreens. The basecompositionofthe oligonu-cleotideswasdeterminedby comparisonwith nucleo-tide standards obtainedbydigestionof the total
un-fractionated RNA(see Fig. 7) (3).Eachspotwascut outof the paperandwas counted in toluene-based scintillation fluid. The relative ratio of eachnucleotide
wasdeterminedfrom theradioactivitypresentineach spot.
RESULTS
Kinetics ofintracellular viral RNA
syn-thesis.MHV infection of L-2orDBT cells did notresult in inhibitionof hostcell macromolec-ular synthesis (datanot shown). Therefore, to
study the intracellular viral RNA, the RNA
synthesisof the host cellswasinhibitedby
ad-dition ofactinomycin D. We found that, with
actinomycinD ataconcentrationof 1
,ug/ml
forDBT cells and 3 pg/mlfor L-2 cells, the host
RNA synthesis was completely inhibited. At
these concentrations, no appreciable effect of
actinomycinDonviralreplicationwasnoticed, asjudged bythe kinetics of virusgrowth,virus
yield, and cytopathic effects. Similar
observa-tionshave also beenmadebyRobb and Bond
(11). However, athigherconcentrations of
acti-nomycin D (5 pg/ml), the yield of infectious
virus from DBT cellswasreducedby 100-fold.
L-2cellswere moreresistanttoactinomycin D, andtherewas noappreciablereduction in virus
yield even at 5
,ug/ml.
Wetherefore usedacti-nomycin D at 1 ug/ml for DBTcells and at 3
Ag/ml
for L-2cellsthroughoutthisstudy. Wefirst determined the kineticsof theintra-cellular virus-specific RNA synthesis in DBT
cells. Thecells wereinfected with MHV-A59at
a multiplicity of one to five and were labeled
with [3H]uridine for 1 h atvarious times after infection. Table 1 shows therateof viral RNA synthesis and virus yieldsatdifferent time points postinfection. Afteralag of2h, thevirus-specific
RNA synthesis reached a peak at 6 to 7 h
postinfection. Thereafter, therateof viral RNA synthesis declined. Bycontrast, the titer of the infectious virus released into the medium did
notreach its peak until 8to 9 hpostinfection. For most ofthe studies, we used intracellular
viral RNA whichwasprepared during maxinum
intracellular RNA synthesis. Studies performed with L-2 cells cultured in thepresenceof 3 pigof
actinomycin Dpermlyielded similar data
(Ta-ble1), with the only exception being that
maxi-mumRNAsynthesiswasdelayed by1 h inL-2 cellscompared with DBTcells.
Sucrosegradient sedimentation and elec-trophoretic analysis of MHV-specific RNA. To determine the species and structure ofthe virus-specific RNA in the MHV-infected cells, the A59-infectedDBTorL-2cellswerelabeled
with[3H]uridineor
'Pi,
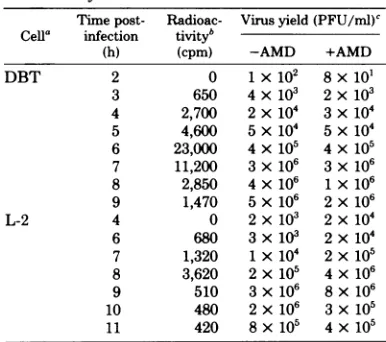
and the totalcytoplas-TABLE 1. Rate of MHV-A59virus-specificRNA
synthesisin DBT and L-2 cells
Time post- Radioac- Virus yield(PFU/ml)' Cella infection tiVityb
(h) (cpm) -AMD +AMD
DBT 2 0 1x 102 8x 101
3 650 4x10( 2 x103
4 2,700 2x104 3x104
5 4,600 5 x104 5 x 104 6 23,000 4 x105 4 x105 7 11,200 3x106 3 x105
8 2,850 4 x106 1x106
9 1,470 5x106 2x106
L-2 4 0 2 x103 2x104
6 680 3x103 2 x104 7 1,320 1x104 2x105
8 3,620 2 x105 4 x106
9 510 3 x106 8x106
10 480 2 x106 3 x105
11 420 8x105 4x105
aDBT
cells
were treated with actinomycin D (AMD)at 1ug/mland[3H]uridineat100pCi/ml.L-2 cellsweretreated withactinomycinDat3pg/mland[3H]uridineat 10pCi/ml.Thecellswerelabeled for1
h atthe specified timepostinfection.After labeling,
thecellsweredisruptedwith 1%SDS,andtheamount
of[3H]uridineincorporation wasdeterminedby
pre-cipitationwithtrichloroacetic acid.
b The amount of trichloroacetic acid-insoluble
[3H]uridine
incorporationafterlabelingfor1h inthe A59-infected cells in the presence ofactinomycinD. Counts of the uninfected cellshavebeensubtracted.'DeterminedbyplaqueassayonDBT
cells.
on November 10, 2019 by guest
http://jvi.asm.org/
[image:3.499.255.448.366.537.2]826 LAI ETAL.
mic RNAwassubjectedtosucrose
gradient
sed-imentation. At least fourRNAspeciescould be
detected: 60S, 32S, 22S, and 18S (Fig. 1). The
60S RNA cosedimented with the
genomic
RNAextracted from thepurifiedvirus(Fig. lb).
Fur-thermore, the T1 oligonucleotide fingerprint of
this RNA wasidentical to that of the 60S viral
genomicRNA(seebelow).Therefore,thisRNA
species represented the genomic RNA of the
virus. The rest of the RNAs
represented
thevirus-specific subgenomic
RNAs,
since themock-infected,
actinomycin
D-treated cellsdidnotcontain anyRNA of this size
(Fig.
la). AnadditionalspeciesofRNA, 4S,
likely
represented28S 18S 4S
a)
10 20 30 40 50 60 FRACTIONNUMBER
70
thetRNA's whichwerefoundin theuninfected
cellsaswell. ThisRNA
species
wasnotfurther
characterized.
The
virus-specific
RNAs were further sepa-ratedby
electrophoresis
on1%agarosegels.Sixvirus-specific
RNA bandscould beresolved(Fig.
2a).
Longer
exposureof thegels
didnotrevealanyadditional RNA bands. When these RNAs
were
analyzed
by
electrophoresisondenaturing
gels
whichcontainedmethylmercurichydroxide
(2),
essentially
similarresultswereobtained. Six RNAspecies
wereconsistently
resolved(Fig.
2b).
However,
in a few RNA preparations, anadditional RNA species was found (Fig.
2c).
These
findings
confirmed the datarecentlyob-tained in other laboratories with the
A59-in-fected Sac (-) or 17 CI-1 cells,whichalso
con-tained sevenvirus-specificRNAspecies (13; H.
Wege,
S.Siddell,
M.Sturm,andV.terMeulen,
in V. ter
Meulen,
S.Siddell,
and H.Wege,ed.,
Biochemistry
andBiology of Coronaviruses,
inpress; J.
Leibowitz,
personalcommunication).
We
designated
theseRNAs,inorder ofdecreas-ing size,
RNAspecies1through7.Themolecularweights
ofthese RNAspeciesweredeterminedfrom the
migration
rates of the RNAs in theagarose
gels containing
methylmercurichydrox-ide
(Fig. 2b,
c). Under these conditions, themigration
rate of the RNA was inverselypro-portional
to the logarithm of the molecular18S
-C)x
0.
CV FRACTION NUMBER
FIG. 1. Sucrose gradient sedimentationof the vi-rus-specific RNA intheMHV-infectedcells. [3H]ur-idine-labeled RNA was extracted from the
MHV-A59-infectedandactinomycin D-treatedDBTcellsat
7hpostinfection andthen sedimentedthrougha 15
to30%linearsucrosegradient containing0.5%SDS inanSW40rotorat20,000rpmfor20h. (a) The
32p-labeled RNA from the mock-infected and
acti-nomycinD-treated DBTcellswascosedimented. (b)
The32P-labeled 60S RNAextractedfromthepurified MHV-A59 virionwasincluded forcomparison. The
positionsofthe28S, 18S, and4SmarkerRNAs are
indicated.
FIG. 2. Agarosegel electrophoresis ofthe
virus-specific RNA in the A59-infectedcells. The 32P-la-beled RNAfromtheA59-infected L-2 cellswasheat
denaturedat100°Cfor1min and electrophoresedon
1%agarosegelwithout(a)orwith (band c) methyl-mercurichydroxideat90Vfor5h.TheRNAs in(a), (b), and (c) were from differentpreparations. The
positionsofthe 35SRoussarcoma virusRNAand 28S and 18SrRNA's areindicated. The gels in(b)
and(c) wereoverexposedto reveal the lesserRNA species.
12
10_
I
1 8 I
X 6
-0~
4
2
_ v
I
o
0
x M 4
C) I
J. VIROL.
on November 10, 2019 by guest
http://jvi.asm.org/
[image:4.499.57.252.225.528.2] [image:4.499.279.426.389.546.2]MHV-A59 mRNA STRUCTURE 827
weight (2). From these data, the molecular
weightsoftheintracellular virus-specific RNAs were determined to be5.4 x 106, 4.0 x 106, 3.0
X 106¶ 1.5X 106, 1.2 x 106,0.9 x 106,and 0.6 x
106 for RNAspecies1through7,respectively.
Itisnoteworthy that the relative ratio of these
RNAspeciesremainedquiteconstant,withthe
exception of RNAs1and4.RNA1 (equivalent
totheviral genome)wasthemostvariable
spe-cies. Insomepreparations,itwasthe
predomi-nantRNAspecies (Fig. 2b, c).Itisprobable that
the ratio of all the subgenomic RNA species
remainedconstant,whereas the increase ofRNA
1 was due to accumulation of genomic RNA
destinedtobeincorporated into the virion. RNA
4 waspresentinthe lowestamountin allof the RNA preparations. In L-2 cells,this RNA spe-cieswasusuallynotdetectable.
Oligonucleotide fingerprintingof the
vi-rus-specific RNAs. Thesumofthe molecular
weights of the six subgenomic RNAs exceeded
the genome size, suggesting that these RNAs
overlap in their sequences. To determine the
structural relationship between the various
RNAspecies,oligonucleotide fingerprintsof
ev-ery virus-specific RNA, except RNA 4, which
was usuallynotdetectable inoursystem, were
examined. The 32P-labeled RNAs were eluted fromagarosegels, digested with RNase T1, and
separated by two-dimensional polyacrylamide gel electrophoresis.The T1 oligonucleotide
fin-gerprints became increasingly complex as the
size of the RNAs increased (Fig. 3).
Further-more,mostof theoligonucleotidespresentin the
small RNAspecieswereincluded in the
finger-prints of thenextlarger RNAspecies.Also,most
of the oligonucleotides in each RNA species
were also presentin the 60Sgenomic RNAof
MHV-A59, suggestingthattheyrepresented
se-quences of the same polarity as the genomic
RNA. The RNA 1 had an oligonucleotide
fin-gerprint identical tothatof the genomic RNA, confirmingthat RNA 1representsthe
intracel-lular form of the genomic RNA. Finally, every
RNA speciescontaineda
poly(A)
spot(located
at the lower left comers of the
fingerprints),
indicating that they are
polyadenylated
andprobablyfunctionasmRNA's.
The oligonucleotide fingerprintsofseveral of
the RNAspeciesalso containedsomeanomalous
oligonucleotides: (i) RNA 7containedan
oligo-nucleotide, 19, which was present only very
weaklyor not at allinothersubgenomic RNA
species;(ii)RNA 6containedan
oligonucleotide,
19a, and RNA 5 contained anoligonucleotide,
3a, both ofwhichwere not present in theviral
genome,nor weretheydetectedinother
subge-nomic RNA species; (iii) RNAs 2 and 3 were
slightly contaminated with the degradation
products ofRNA 1; however, thecontaminating
oligonucleotides could bediscerned due to the muchlowermolar ratioatwhich they occurred (Fig. 3). It appeared that the amounts of some
of the oligonucleotides, e.g., oligonucleotides 3
and5in RNA 3, were reduced in RNA 2. The
origin of these anomalous oligonucleotides is
currently beingstudied.Itispossible that these
oligonucleotidesrepresent the extreme 5' ends
of the subgenomicRNA species.
Mapping of thesubgenomic RNA species
onthe RNAgenome. Theoligonucleotide
fin-gerprints of the virus-specificRNA species
sug-gested that the MHV subgenomic RNAs had
overlapping sequences, with the sequences of
the smaller RNA being contained within the
next-larger RNA species.However, these studies
didnotreveal the genetic localization of these
mRNA'sonthe RNAgenome.Itwasimportant
to determine the genetic regions from which
each mRNAspecieswastranscribed. To
accom-plish this,we studied themapposition of each
oligonucleotideonthe RNA genome. The
par-tially degraded 3P-labeled 60SRNAofA59 was
fractionated by chromatography on
oligo(dT)-cellulose. Thepoly(A)-containing RNAwas
se-lected and fractionated bysucrose gradient
sed-imentation. The genomic RNA from A59 was
degradedtosuchanextentthat thepeak size of
the RNAwas188 (Fig. 4). The
poly(A)-contain-ing RNAs of different sizeswerepooledasshown
in Fig. 4 and analyzed by T1 oligonucleotide
fingerprinting. Several of the RNAfingerprints
are presented in Fig. 5. All these RNAs
con-tained the sequences starting from the 3' end
andextending for various distance toward the 5'
endof thegenome.Therefore,therelative
posi-tion ofeacholigonucleotideonthe RNAgenome
could be arranged in several groups from 3'
toward the5' end.Furthermore,thepositionof
theoligonucleotidesonthe RNAgenomecould
be estimated from the smallest size of the
poly(A)-containing RNA in which the
oligonu-cleotidewasdetected. Bythisapproach,all the
oligonucleotides of thegenomicRNA of MHV-A59 were mappedin severalgroupsonthe
ge-nome (Fig. 6). By comparison with this
oligo-nucleotide map, it.was found that all of the
oligonucleotidesinRNA7 werelocatedatthe3'
end of thegenome. Therestof thevirus-specific
RNAs alsocontained theoligonucleotides start-ing from the 3' end and extending for various
lengths toward the 5' end (Fig. 6). This result
suggested thatthesubgenomic RNAsof
MHV-A59hada structuresimilartothatproposedfor
avian infectiousbronchitisvirus (14, 15).
How-ever, itwas also clear from Fig. 6 that several
RNAspecies,including2, 3, 5,and6, contained
the oligonucleotides which either were absent
on November 10, 2019 by guest
http://jvi.asm.org/
828 LAI ET AL.
A.
:E0-
A0
~
0
B o0
C
00
00500 0OO 10 X52
37. C-O44~~ 1
350 4342 1 ~ 3
340 16 822
00 10
[image:6.499.60.450.74.572.2]14 13 <1f05
FIG. 3. Oligonucleotide fingerprintingof the intracellular A59-specific RNAs. The virus-specific RNAs wereelutedfromagarose gels, digested with RNaseT1, and separated bytwo-dimensionalpolyacrylamide gel electrophoresis. Electrophoresiswasfromleft to right in the firstdimension and from bottom to top inthe second dimension. The numbering of the spots is as indicated in the schematic sketch (B). Only the oligonucleotides whichappearedforthefirst time in the mRNA's of increasing size were numbered. The numbersattheupperleftcornersofeach panel represent the RNA species referred to inFig.2.Abbreviations: V,viriongenomic RNA;A, poly(A).
from theviral genome or were not contiguous nome.Whether theseoligonucleotidesrepresent
with therestof theoligonucleotides onthege- splicing of the sequences is unknown at the
J. VIROL.
on November 10, 2019 by guest
http://jvi.asm.org/
MHV-A59 mRNA STRUCTURE 829
I A1 - 1 -
HF--25 -28S
CM 35S
20
'-l 60S
x
E 15
0
X 10
5
0 5 10 15 20
Fraction Number
FIG. 4. Sucrosegradientsedinentationof the
par-tially degraded poly(A)-containing A59 genomic RNA. Thepoly(A)-containingfraction fromthe
par-tially degradedA59RNAwasselectedby
oligo(dT)-cellulose column chromatographyand sedimentedon
a 10 to25%sucrosegradient made in 0.01MTris hydrochloride,pH7.4, 0.01MNaCI,and 0.05% SDS
inanSW41 rotorat40,000rpmfor4.Eh. TheRNA
ofdifferent sizeswaspooled into the fivefractionsas
indicated.
presenttime.
Sequences immediately adjacenttothe 3' ends of thevirus-specific RNAs.The oligo-nucleotidemappingstudiessuggestedthatallof
the virus-specific RNAs contained the oligo-nucleotides starting from the 3' end of the
ge-nome. Thus, they should have identical 3' end
sequences.However,this conclusionwas
uncer-tain since severalRNAspeciescontained oligo-nucleotidesnotpresentin thegenome,andsome
of theoligonucleotidesinthe smaller RNA spe-cieswerenotpresentin thelargerRNAs. Itwas
therefore possible that some of these RNAs
might have different 3'-endsequences. To test
this possibility, the poly(A) spotsfrom the
oli-gonucleotide fingerprintsof each RNA (Fig. 3)
wereisolated, digestedwith RNaseA,and sep-aratedbyelectrophoresisonDEAE-cellulose
pa-peratpH 3.5. Thesepoly(A) spotsshould
con-tain not only the poly(A) tracks but also the
sequences which are immediately adjacent to
the poly(A) and extend inward until the first guanineresidue.Figure7shows suchananalysis
for RNAs 5, 6, and 7. From the molar
yields of each base, the base composition of
the sequences immediately adjacent to the poly(A) spots was determined to contain C(AC)(AG)(A2U)(A3C)(A4N)Alol-13o (Table 2).
It canbe seen that all of the RNAs contained
identical base compositionsandlikely had
iden-tical sequences at the 3' end. These data also
show that all of the RNA species contained a
poly(A) stretch of 100 to 130 nucleotides long.
The presence of (AG) residues within this
stretchcannotbe explainedat thepresenttime.
It was reproducible and may represent
se-quencesprotected from RNase T1 digestionby
the poly(A) stretch. Similar studies have been
performed with thepoly(A) spots of the RNAs
1, 2, and 3. All ofthem appeared to have the samebase sequences,although the molar yield
of each base was not confirmed by counting
radioactivity because ofthe low counts
associ-ated with each spot (data not shown). These
data thereforesuggestthatallthevirus-specific
RNA species contained identical 3' end
se-quences.
Genetic localization of the sequence di-vergencebetween A59and MHV-3.Wehave
previouslyshown that A59,aweakly pathogenic
strain, and MHV-3, a strongly hepatotropic
strain, haveverysimilaroligonucleotide
finger-printsexceptfortwounique oligonucleotidesin
MHV-3 and four in A59 (7). Some of these
oligonucleotides might be derived from the
ge-netic regionsresponsible for the biological
dif-ferences between these two virus strains. We
therefore attemptedtolocalizethese
oligonucle-otides on the RNA genome. As shown
previ-ously,A59contains fouruniqueoligonucleotides,
9, 13,19,and 28, whicharenotpresentin
MHV-3 (7). Comparison of Fig. 3 with Fig. 5 shows thattwoofthe A59-specific oligonucleotides, 9 and13, were presentinthegeneticregionat6 to
10kilobases from the 3' end which corresponds
toRNA3(Fig.6). Incontrast,thegenetic
local-ization of theoligonucleotides 19 and 28 is less
certain. Although oligonucleotide 19 appeared
tobepresentin RNA7, itwasabsent in RNAs 5and6. Oligonucleotide28appearedalsotobe presentin RNA3 (Fig. 3),but itwasfoundby fingerprinting degradedvirion RNAtobe closer
tothe 5' end ofthegenome (Fig. 5).This
oligo-nucleotide was present at lower molar yields
than other
oligonucleotides,
thusincreasingthedifficultyofprecise
genetic
mapping.
Theoligon-ucleotide 28 was therefore not
represented
inFig.6.
We alsomappedthetwo
MHV-3-specific
oli-gonucleotides,aandb,onthe RNAgenomeby
fingerprinting the
poly(A)-containing
RNA ofdifferent size fractions.
Figure
8showssomeofthe representative
fingerprints.
By
thisap-proach, itwasdeterminedthat a waslocalized
at 6 to 7 kilobasesfrom the 3'
end,
whereas bwaslocalizedvery closetothe 5' end.Thesetwo
regions correspond to the
regions
containedwithin RNA 3 and RNA 1,
respectively.
Fromon November 10, 2019 by guest
http://jvi.asm.org/
[image:7.499.53.244.62.264.2]J. VIROL. 830 LAI ET AL.
FIG. 5. T1-oligonucleotidefingerprints of the poly(A)-containingA59 RNAsofdifferent size fractions. The RNAscollectedasindicated inFig.4wereanalyzedbyT,oligonucleotide fingerprinting.Oligonucleotides whichappeared for the first time intheRNAsof increasing sizewerenumbered. The numbering of the total largeT1oligonucleotidesisshownforthe60S RNAand isidenticaltothat inFig.3(B).
the genetic mapsof A59 and MHV-3, we
con-cluded thatatleast RNAs3and1contained the
geneticdifferencesbetween thesetwostrains.
DISCUSSION
Our resultsshowthat L-2 cellsinfectedwith
the A59 strain of MHV contain six to seven
on November 10, 2019 by guest
http://jvi.asm.org/
MHV-A59 mRNA STRUCTURE 831 608
4
45S
4
35S 28S 18S 15S
4
4
4
18 16 14 12 10 8 6 4 2 0
5D
I i I i - tl%35'~
~~i
1[1[@
§ § | 1[12 ][4,30,
][,40 31
33,31,21,7 4 22337 19mRNA
(19a) # 7
(16) (5.15,11) .#, *2
w*#2
VA
#1FIG. 6. Oligonucleotidemap of the A59 genome and its corresponding subgenomic RNAs. TheT1 oligo-nucleotides of the A59genomic RNA (numbered as shown in Fig. 3 and 5) were arranged into several groups ondifferent regions ofthe genome. Since the genomic RNA has a molecular weight of 5.4 x1iO,thegenome length is assumedto be 18kitobases. The subgenomic RNAs are also presented. The oligonucleotides in parenthesesondifferent subgenomicRNAs either wereabsentfromthegenomic RNA or were not localized in the corresponding regionsof the genome. Oligonucleotides in common with those on the genomic RNA were not labeledon thesubgenomic RNAs. The circled spots, 9, 13, and 19, arespecificfor A59 andnot presentin MHV-3(7).
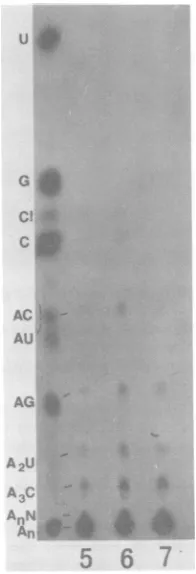
FIG. 7. Base sequenceanalysisofthe
poly(A)-con-taining oligonucleotides ofthe A59 mRNA's. The
poly(A)-containing oligonucleotides were isolated from the oligonucleotidefingerprintsofeach virus-specificRNAspecies asshown inFig.3. These oli-gonucleotidesweredigestedwith RNase A and elec-trophoresedonDEAE-cellulosepaperinacetic acid-pyridinebuffer,pH 3.5, at 1,500Vfor4h. The60S RNAdigestedwith RNase A andT,wasincludedas
TABLE 2. Partial base sequenceanalysisofthe 3'-end sequencesa
RNAspecies Basecomposition 5 C(AC)(AG)(A2U)(A3C)(A4N)Aioi
6 C(AC)(AG)(A2U)(A3C)(A4N)Aj13 7 C(AC)(AG)(A2U)(A3C)(A4N)A13o
a The poly(A)-containing
oligonucleotides
were eluted from thetwo-dimensionalfingerprints, digested with RNase A, andseparated by electrophoresis onDEAE-cellulosepaper atpH 3.5 (Fig. 1).Each spot wascutout, and therelative ratio of each spotwas determinedbycountingintoluene-based scintillation fluid.
virus-specific RNA species. Similar findings
haverecently been obtained with other cell lines,
including Sac (-) and 17CI-1 cells(13;Wegeet
al., in press;J. L. Liebowitz, K. C.Wilhehnsen,
and C. W. Bond, personal communication).
However, in L-2 cells, one of the al., in press;
Liebowitzetal.,personal communication).
How-ever, in L-2cells, oneof the RNAs, 4, wasnot
always detectable or present only in very low
quantity. It is not clear whether this variance represents a host cell-determined differential expression of viral RNA. We have also shown
that all of the virus-specific RNAs contain
poly(A)segmentsof100 to130nucleotideslong.
This is similar to the length of the poly(A)
present in the viral genome, which has been
estimated tobe about 90nucleotides
long
(21).
mono-andoligonucleotidemarkers(3). The relative ratio ofeach spotwasdetermined. Onlythosefrom
the mRNA's 7, 6, and5 areshown here. Abbrevia-tions:A,AMP; C,CMP; G,GMP; U, UMP;C."cyclic
CMP;N,anynucleotidesotherthanAMP.
on November 10, 2019 by guest
http://jvi.asm.org/
[image:9.499.52.445.59.200.2] [image:9.499.99.197.289.576.2]832 LAI ET AL.
FIG. 8. Oligonucleotidefingerprints ofthepartiallydegraded, poly(A)-containingMHV-3genomicRNA species. The32P-labeled,60SMHV-3 RNAwasbriefly digested,selectedby oligo(dT)-cellulose
chromatogra-phy, and sedimentedbysucrosegradientsedimentationasdescribedforMHV-A59(Fig. 4).Therepresentative
RNAfractionswerepooled andanalyzedbyT,oligonucleotidefingerprinting. Only representativefingerprints
areshown here.(A) 18S;(B) 35S; (C) 50S; (C) 60S. Oligonucleotidesaandbarespecificfor MHV-3 andnot presentin A59(7).
The presence of poly(A) sequences in these RNAs suggests that they represent mRNA's.
This conclusion is consistent with the recent
findings that these virus-specific RNAs were
present in the polysome fractions of MHV-in-fected cells(M. M.C.Lai, C.D.Patton, andS.
A. Stohlman, unpublished data; 13) and that
these RNAscouldserve asmnRNA'sfor in vitro translation (12; S.Siddell,H.Wege,A.Barthel, and V.terMeulen, inV. terMeulen, S.Siddell,
andH. Wege, ed., BiochemistryandBiologyof
Coronaviruses, in press).
Thestructureof these viral mRNA's contains
someveryunusual features.Theoligonucleotide
fingerprinting studies suggest thattheyhave a
"nestedstructure,"astermedbyStern and
Ken-nedy (14, 15); i.e., the sequences of the small
mRNA are contained within the next-larger
mRNA species. Furthermore, the mapping of
theoligonucleotidesonthe genome suggests that
the sequencesof all of thesemRNA'sstartfrom the 3' endof the genome. These data strongly suggest that the structures of the mRNA's of MHV-A59 arevery similartothatof avian in-fectious bronchitisvirus (14, 15). In further sup-port of this proposal, we found that all ofthe
mRNA species contained identical sequences
immediately adjacent to the 3'-poly(A). The
presence ofseveral anomalousoligonucleotides
insomemRNAspecies, however,suggeststhat
MHVmRNA'smightbemorecomplexthan the
infectious bronchitis virus model. Some
oligo-J. VIROL.
on November 10, 2019 by guest
http://jvi.asm.org/
[image:10.499.60.456.69.428.2]MHV-A59 mRNA STRUCTURE 833
18
5'
I
AI l
16 14
i
70
l
B I
12 10
i F
Il
I
c
8
D
'E
Fi
G6 4 2 0 Kb
§ I F 3'
| | mRNA
#5
4'
#4
I
I
I
I
#
3AvAI #2
[image:11.499.45.444.51.200.2]II
I~~
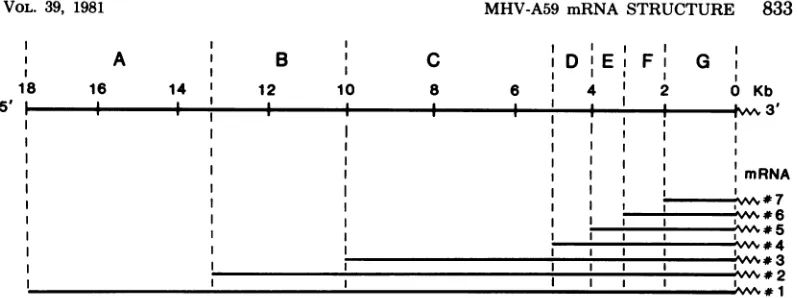
# 1FIG. 9. Proposedmedel of the structure of the MHVmRNA's and the corresponding genetic regions on the genome. The proposed seven genetic regions on the MHV genome are based on the data reported here and from other laboratories (13; Wege et al., in press; Leibowitz, personal communication). The boundaries of each gene were estimated by converting the molecular weights of the RNAs into kilobases (kb). Each gene corresponds to the portion ofeach mRNA species which does not overlap with the next-smaller mRNA species.
nucleotideswerepresentin thesmaller mRNA's
butnotinthelargerRNAs,whereasothers were
notpresentin the genome. These findings
sug-gestthatthe 5' ends of these mRNA'scould be
quite unusual. Preliminary data suggestthatall
of them contain thecapsequencesatthe 5' ends.
Weare currently investigating the
oligonucleo-tides whichmightcontain sucha capstructure.
ThemRNA's with suchastairlike relationship
areverysimilar tothose found in retroviruses
and togaviruses (5, 16, 21). In these mRNA's,
usuallyonlytheportionof the sequences which
is unique to a particular mRNA species and
which is not shared with the next-smaller
mRNA istranslated. Ifasimilar relationship of
mRNA'stogenomeoccursduringreplication of
MHV, the MHV RNAgenomecould be divided
intosevengenetic regionscorrespondingtothese
mRNA species (Fig. 9). This model offers a
rational basis for studying the geneticstructure
of the RNA genome of MHV. Thecoding
poten-tials of the mRNA's in another strain of MHV,
JHM, have been studiedbyin vitrotranslation
(12;Siddelletal.,inpress).If these datacanbe
appliedtothe A59strain, itcould suggest that thegeneproductsof thefollowinggenesare:for B,anonstructural
protein
p30;forC,
gpl80/90;for DorE, nonstructuralp14; forF, gp23; and
forG, pp6O. In analogytoother virus systems, gene A might code foran RNApolymerase. It remains to beinvestigated whether MHV-A59 codes forsimilarproducts.
The studyof theoligonucleotide maps ofall
of the mRNA species has also allowed us to
localize the genetic differences between two
MHV strains,MHV-3 and A59. Previously, we
have shown that these two strains differ only
slightlyintheiroligonucleotide fingerprints (7).
Since these two strainsare genetically related
butdiffer in their pathogenic
potentials-MHV-3 causes severe hepatitis, whereas A59is only
weakly pathogenic-these oligonucleotide
dif-ferences could suggest the genetic regions
re-sponsiblefor suchpathogenic potentials (7). Our
mapping studies indicate that one of these
re-gions might correspond to mRNA 3 and thus
would reside ingene C, which codes for gp9O/
180,whereas the othercorresponds tomRNA 1
and therefore would reside in gene A, which
might code for an RNA polymerase. Whether
thesetworegionsaredirectlyresponsiblefor the
viralpathogenicityremainstobeinvestigated.If
gene C is indeedresponsiblefor the viral
path-ogenicity, itwill suggestan interesting
mecha-nism for viralpathogenesis:thegeneproductof
thisgene, gp90/180, is themajorenvelope
gly-coprotein in MHV (17) and could determine the host range of the virus. The nonpathogenic
strain, A59, might have mutations inthis gene
and thus becameunabletoinfect thetargetcells.
Further studiesarerequiredtoresolve thisissue.
ACKNOWLEDGMENTS
Wethank T. Hanson for excellent technical assistance and J.Lopez for editorialhelp.
This work was supported in part by grant PCM-4567, awardedbytheNationalScienceFoundation,andbyPublic HealthService grants NS 15079and CA16113,awardedby theNational Institutes of Health.
ADDENDUM INPROOF
We haverecentlyshownthat all of theA59-specific RNAs containidentical5' endsequences, suggesting
synthesisof these RNAs by splicingor another
un-usualmechanism (M.M.C.Lai,C. D.Patton,andS. A.Stohlnan,submitted forpublication).
LITERATURE CITED
1. Aviv,H.,andP.Leder.1972.Purification ofbiologically
active globin messenger RNA by chromatographyin
oligothymidylic acid-cellulose. Proc. Natl. Acad. Sci.
on November 10, 2019 by guest
http://jvi.asm.org/
834 LAI ET AL. U.S.A. 59:1408-1412.
2. Bailey, J. M.,and N. Davidson. 1976.Methylmercury as areversible agent for agarose gel electrophoresis. Anal. Biochem. 70:75-85.
3.Barrell,B.G.1971.Fractionation and sequenceanalysis ofradioactive nucleotides, p. 751-779. In G. L. Cantoni and D. R. Davies (ed.), Procedures in nucleic acid research, vol. 2. Harper & Row,Publishers, New York.
4. Guy,J.S.,and D. A. Brian. 1979. Bovine coronavirus genome. J. Virol. 29:293-300.
5.Hayward,W. S. 1977. Size andgeneticcontentof viral RNAs in avian oncovirus-infectedcells.J.Virol. 24:47-63.
6. Lai, M. M.C., and S. A.Stohlman.1978. RNAof mouse hepatitis virus. J. Virol. 26:236-242.
7. Lai, M. M.C.,and S. A.Stohlman.1981.Comparative analysis ofRNA genomes of mouse hepatitisviruses.J. Virol. 38:661-670.
8. Lomniezi,B., and I. Kennedy.1977. Genome of infec-tious bronchitis virus. J. Virol. 24:99-107.
9. MacNaughton, M. R., and M. H. Madge. 1978. The genome of human coronavirus strain 229E. J. Gen. Virol.39:497-504.
10.McIntosh, K. 1973. Coronaviruses: a comparative review. Curr.Top. Microbiol.Immunol.63:85-129.
11. Robb,J.A., and C. W. Bond. 1979. Pathogenicmurine coronaviruses. I.Characterization of biological behavior invitro and virus-specificintracellularRNA ofstrongly neurotropic JHM and weakly neurotropic A59V viruses. Virology94:352-370.
12. Siddell,S. G., H. Wege, A. Barthel, and V. ter Meulen. 1980.Coronavirus JHM: cell-free synthesis of structural proteinp60.J.Virol. 33:10-17.
13.Spaan, W. J. M., P. J. M. Rottier, M. C. Horzinek, and B. A. M.vanderZeijst.1981.Isolation and identifi-cation ofvirus-specificmRNAs in cells infected with mousehepatitis virus (MHV-A59). Virology 108:424-434.
14. Stern,D.F., and S. I. T.Kennedy. 1980. Coronavirus multiplication strategy. I. Identification and character-ization ofvirus-specific RNA. J. Virol. 34:665-674. 15. Stern, D.F., and S. L. T.Kennedy. 1980. Coronavirus
multiplication strategy.II.Mapping the avian infectious bronchitis virusintracellular RNAspeciestothe ge-nome. J. Virol.36:440-449.
16. Strauss, J.H., and E. G. Strauss. 1977.Togaviruses,p.
111-166.In D. P.Nayak (ed.), The molecular biology of animal viruses.MarcelDekker,Inc., New York. 17. Sturman, LS. 1977. Characterization of a coronavirus.
I. Structuralproteins: effects of preparative conditions onthe migration of protein in polyacrylamide gels. Virology 77:637-649.
18.Tyrrell,D.,J.Almeida,C.Cunningham,W.Dowdle, M.Hofstad, K.McIntosh, M. Tajima, R. Zakstel-skaya, B.Esterday, A. Kapikian, and R. Bingham. 1975.Coronaviridae. Intervirology 5:76-82.
19.Wege,H., A.Muller,andV. terMeulen.1978.Genomic RNA of the murine coronavirus JHM. J. Gen. Virol. 41:217-227.
20. Weiss, S. R., H. E. Varmus, and J. M. Bishop. 1977. The size and genetic composition of virus-specific RNAs in thecytoplasm ofcellsproducingavian sarcoma-leu-kosis viruses. Cell 12:983-992.
21.Yogo, Y., N.Hirano, S. Hino, H. Shibuta, and M. Matumoto. 1977.Polyadenylate in the virion RNA of mousehepatitisvirus. J. Biochem. 82:1103-1108.
J. VIROL.
on November 10, 2019 by guest
http://jvi.asm.org/